Pilot-Scale Production of Chito-Oligosaccharides Using an Innovative Recombinant Chitosanase Preparation Approach



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Culture Conditions
2.3. Enzyme Purifications
2.4. Chitosanase Activity Assay
2.5. Determination of Protein Concentration
2.6. Bulk Preparation of Chitosanase
2.7. Thin-Layer Chromatographic Analysis of Enzymatic Hydrolysate
2.8. Pilot-Scale Production of Chito-Oligosaccharides
3. Results and Discussions
3.1. Enzyme Expression and Induction
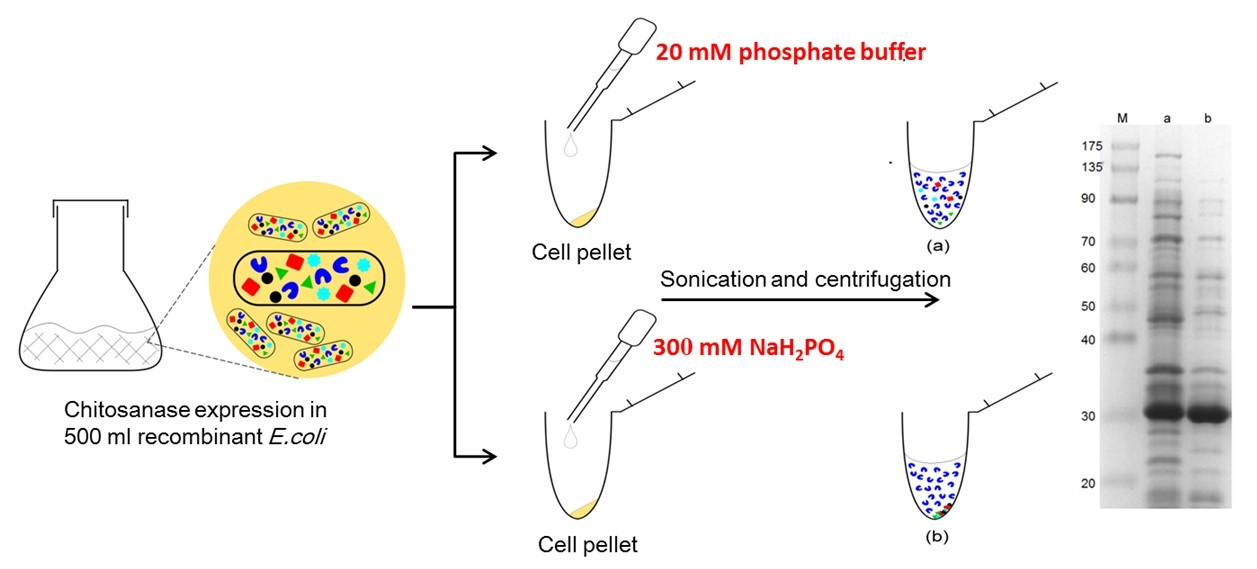
3.2. Effect of Chemical Species on Protein Precipitation
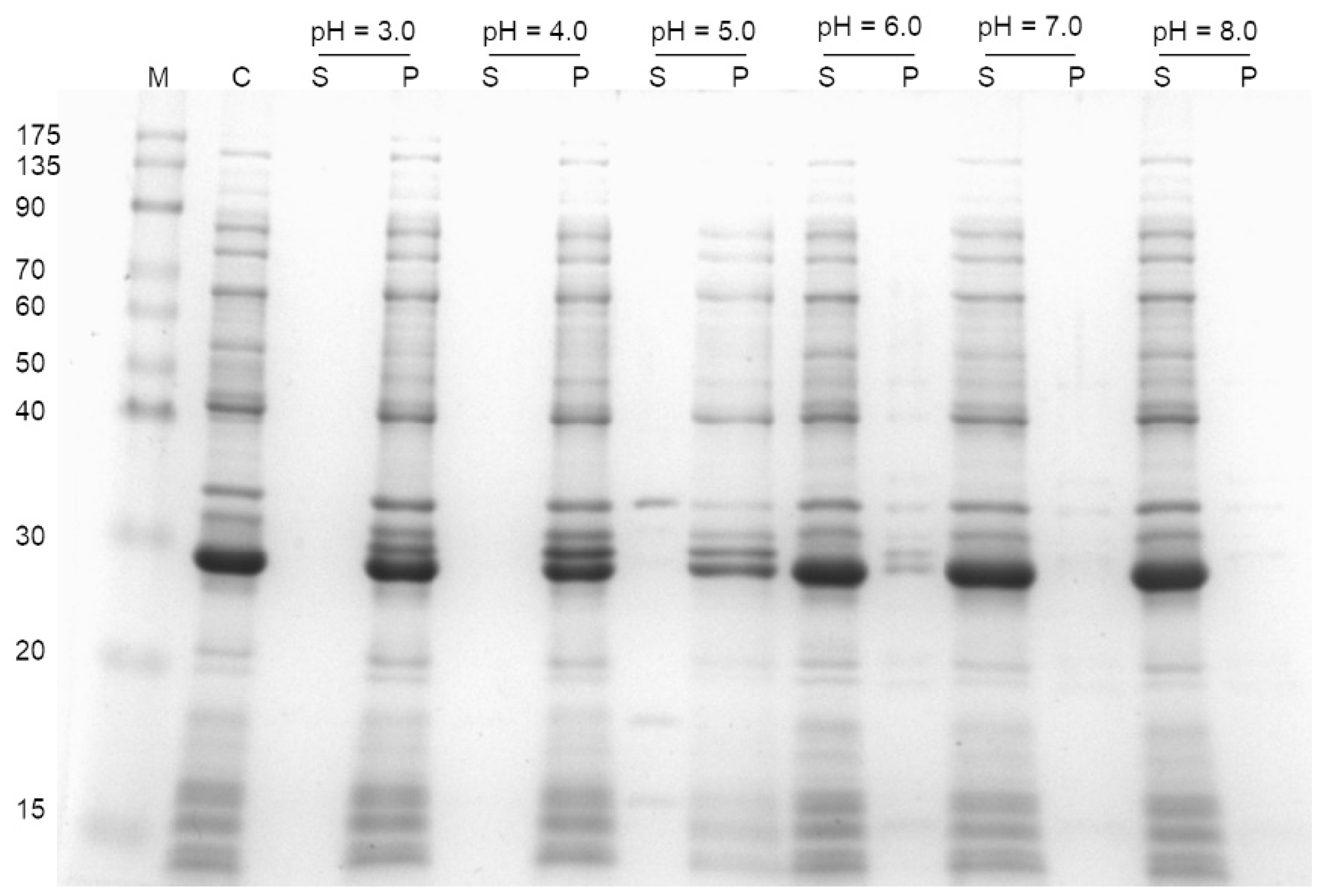
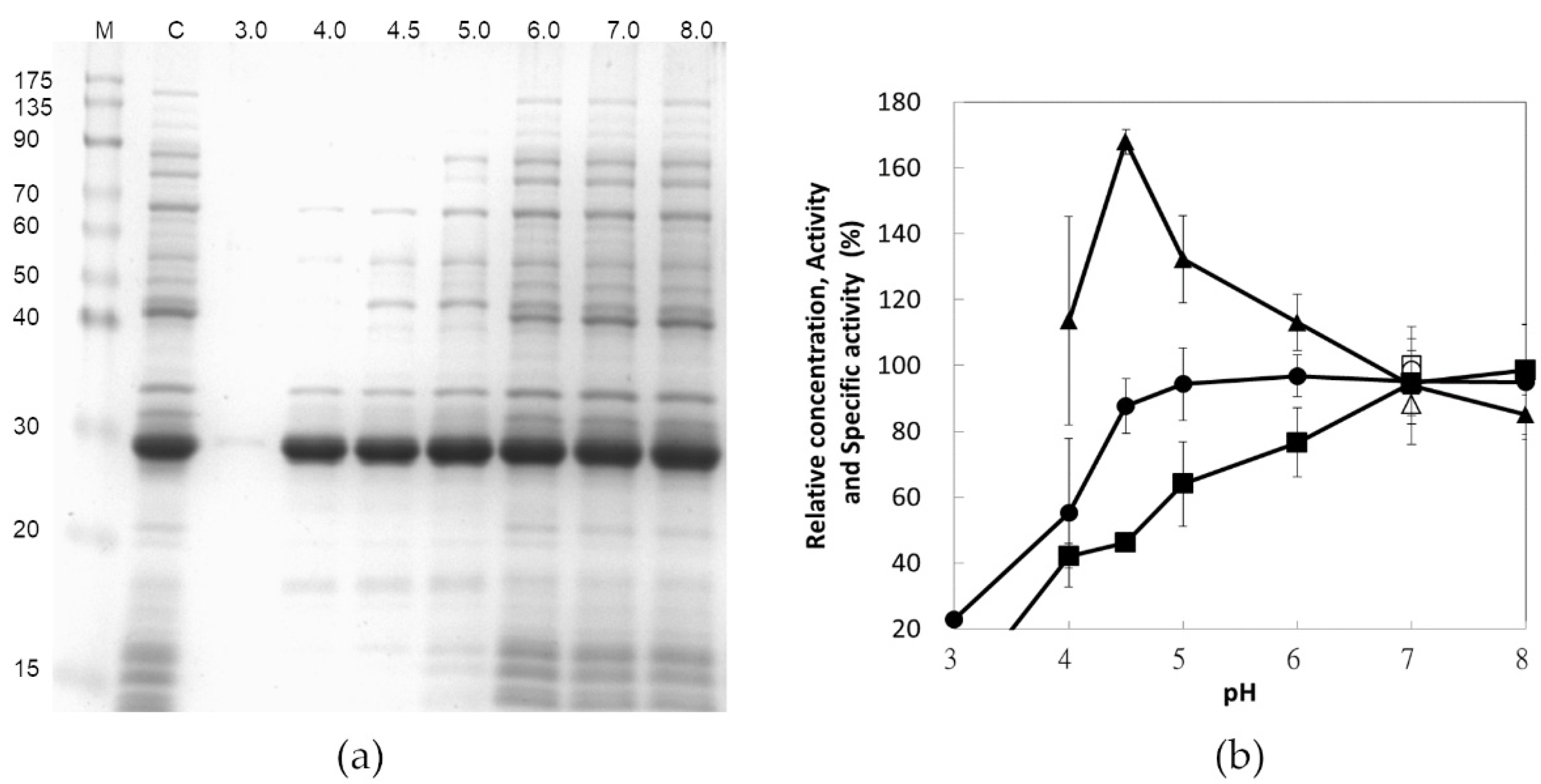
3.3. Effect of pH on Protein Precipitation
3.4. Effect of Buffer Concentration on Protein Precipitation
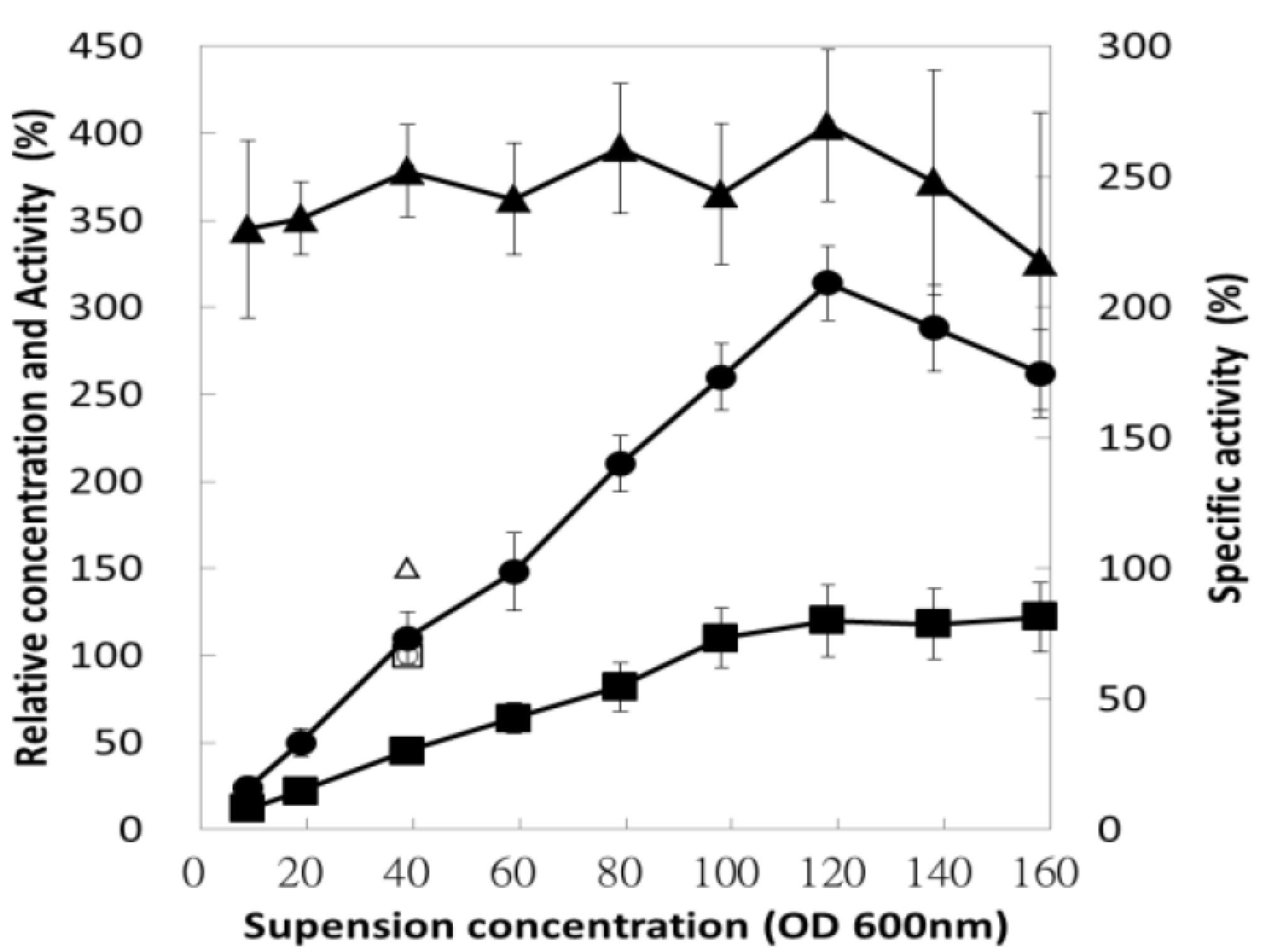
3.5. Effect of Bacterial Suspension Concentration
3.6. Bulk Preparation of Chitosanase
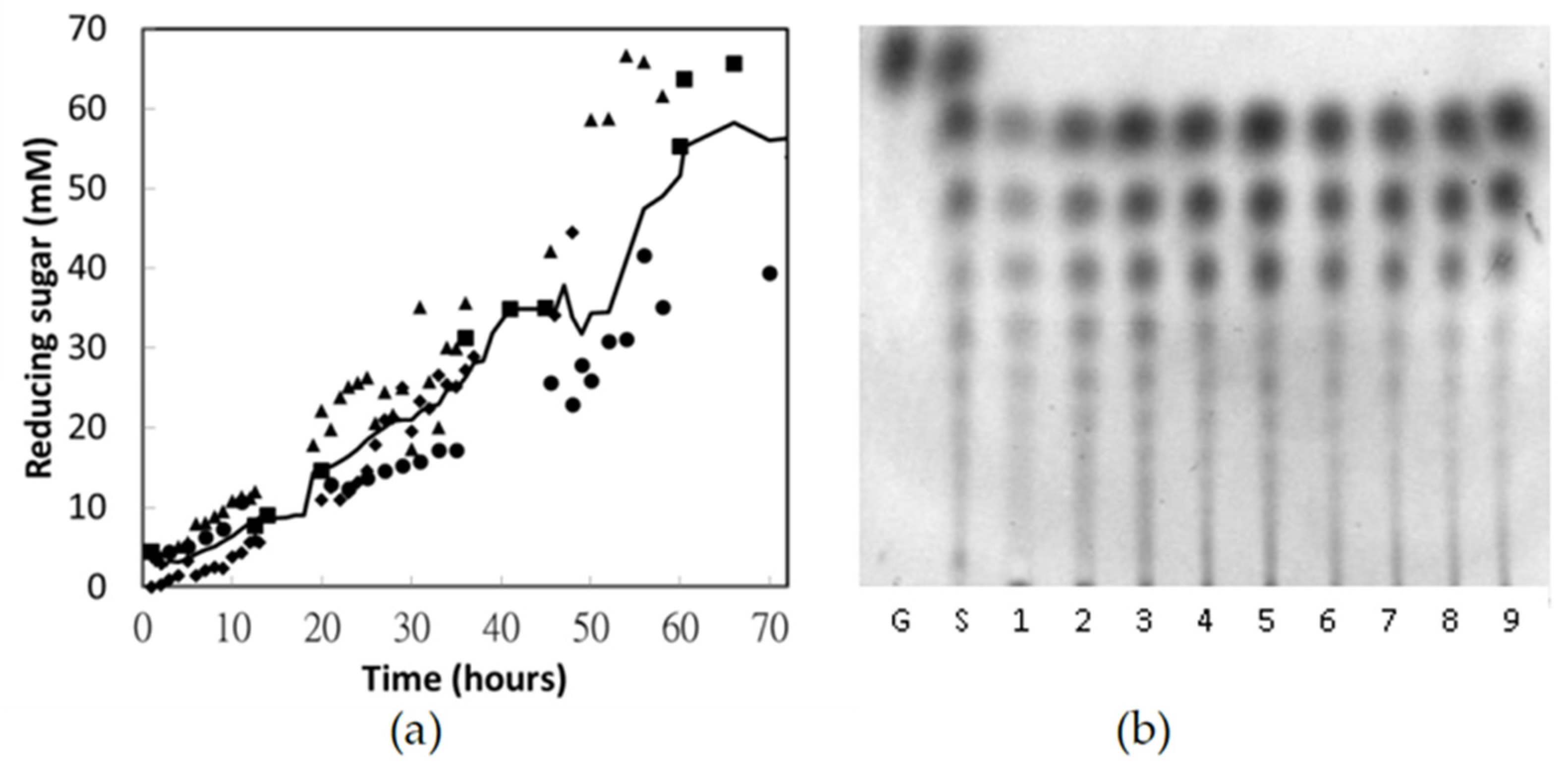
3.7. Pilot-Scale Production of Chito-Oligosaccharides
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Somashekar, D.; Joseph, R. Chitosanases—Properties and applications: A review. Bioresour. Technol. 1996, 55, 35–45. [Google Scholar] [CrossRef]
- Suzuki, K.; Mikami, T.; Okawa, Y.; Tokoro, A.; Suzuki, S.; Suzuki, M. Antitumor effect of hexa-N-acetylchitohexaose and chitohexaose. Carbohydr. Res. 1986, 151, 403–408. [Google Scholar] [CrossRef]
- Qin, C.; Du, Y.; Xiao, L.; Li, Z.; Gao, X. Enzymic preparation of water-soluble chitosan and their antitumor activity. Int. J. Biol. Macromol. 2002, 31, 111–117. [Google Scholar] [CrossRef]
- Zikakis, J.P. Chitin, Chitosan, and Related Enzymes; Academic Press: London, UK, 1984. [Google Scholar]
- Muzzarelli, R.; Jeuniaux, C.; Gooday, G.W. Chitin in Nature and Technology, 3rd ed.; Plenum Press: New York, NY, USA, 1985. [Google Scholar]
- Aktuganov, G.E.; Melentiev, A.I.; Varlamov, V.P. Biotechnological Aspects of the Enzymatic Preparation of Bioactive Chitooligosaccharides. Appl. Biochem. Microbiol. 2019, 55, 323–343. [Google Scholar] [CrossRef]
- Ming, M.; Kuroiwa, T.; Ichikawa, S.; Sato, S.; Mukataka, S. Production of Chitosan Oligosaccharides at High Concentration by Immobilized Chitosanase. Food Sci. Technol. Res. 2006, 12, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Kuroiwa, T.; Noguchi, Y.; Nakajima, M.; Sato, S.; Mukataka, S.; Ichikawa, S. Production of chitosan oligosaccharides using chitosanase immobilized on amylose-coated magnetic nanoparticles. Process Biochem. 2008, 43, 62–69. [Google Scholar] [CrossRef]
- Jeon, Y.-J.; Kim, S.-K. Continuous production of chitooligosaccharides using a dual reactor system. Process Biochem. 2000, 35, 623–632. [Google Scholar] [CrossRef]
- Santos-Moriano, P.; Woodley, J.M.; Plou, F.J. Continuous production of chitooligosaccharides by an immobilized enzyme in a dual-reactor system. J. Mol. Catal. B Enzym. 2016, 133, 211–217. [Google Scholar] [CrossRef]
- Kumar, A.B.V.; Varadaraj, M.C.; Gowda, L.R.; Tharanathan, R.N. Characterization of chito-oligosaccharides prepared by chitosanolysis with the aid of papain and Pronase, and their bactericidal action against Bacillus cereus and Escherichia coli. Biochem. J. 2005, 391, 167–175. [Google Scholar]
- Terbojevich, M.; Cosani, A.; Muzzarelli, R.A.A. Molecular parameters of chitosans depolymerized with the aid of papain. Carbohydr. Polym. 1996, 29, 63–68. [Google Scholar] [CrossRef]
- Sun, L.; Adams, B.J.; Gurnon, J.R.; Ye, Y.; Van Etten, J.L. Characterization of Two Chitinase Genes and One Chitosanase Gene Encoded by Chlorella Virus PBCV-1. Virology 1999, 263, 376–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, C.H.; De Zoysa, M.; Kang, D.H.; Lee, Y.D.; Whang, I.S.; Nikapitiya, C.; Heo, S.J.; Yoon, K.T.; Affan, A.; Lee, J.H. Isolation, Purification, and Enzymatic Characterization of Extracellular Chitosanase from Marine Bacterium Bacillus subtilis CH2. J. Microbiol. Biotechnol. 2011, 21, 1021–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.Y.; Li, Y.-K. An Aspergillus chitosanase with potential for large-scale preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem. 2000, 32, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Osswald, W.F.; Shapiro, J.P.; Doostdar, H.; McDonald, R.E.; Niedz, R.P.; Nairn, C.J.; Hearn, C.J.; Mayer, R.T. Identification and Characterization of Acidic Hydrolases with Chitinase and Chitosanase Activities from Sweet Orange Callus Tissue. Plant Cell Physiol. 1994, 35, 811–820. [Google Scholar] [CrossRef]
- Henrissat, B.; Callebaut, I.; Fabrega, S.; Lehn, P.; Mornon, J.P.; Davies, G. Conserved catalytic machinery and the prediction of a common fold for several families of glycosyl hydrolases. Proc. Natl. Acad. Sci. USA 1995, 92, 7090–7094. [Google Scholar] [CrossRef] [Green Version]
- Viens, P.; Lacombe-Harvey, M.-È.; Brzezinski, R. Chitosanases from Family 46 of Glycoside Hydrolases: From Proteins to Phenotypes. Mar. Drugs 2015, 13, 6566–6587. [Google Scholar] [CrossRef]
- Ando, A.; Noguchi, K.; Yanagi, M.; Shinoyama, H.; Kagawa, Y.; Hirata, H.; Yabuki, M.; Fujii, T. Primary structure of chitosanase produced by Bacillus circulans MH-K1. J. Gen. Appl. Microbiol. 1992, 38, 135–144. [Google Scholar] [CrossRef] [Green Version]
- De Araújo, N.K.; Pagnoncelli, M.G.B.; Pimentel, V.C.; Xavier, M.L.O.; Padilha, C.E.A.; De Macedo, G.R.; dos Santos, E.S. Single-step purification of chitosanases from Bacillus cereus using expanded bed chromatography. Int. J. Biol. Macromol. 2016, 82, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Goo, B.G.; Park, J.K. Characterization of an alkalophilic extracellular chitosanase from Bacillus cereus GU-02. J. Biosci. Bioeng. 2014, 117, 684–689. [Google Scholar] [CrossRef]
- Ho, S.L.; Lan, J.C.-W.; Tan, J.S.; Yim, H.S.; Ng, H.S. Aqueous biphasic system for the partial purification of Bacillus subtilis carboxymethyl cellulase. Process Biochem. 2017, 58, 276–281. [Google Scholar] [CrossRef]
- Liang, T.-W.; Chen, W.-T.; Lin, Z.-H.; Kuo, Y.-H.; Nguyen, A.D.; Pan, P.-S.; Wang, S.-L. An Amphiprotic Novel Chitosanase from Bacillus mycoides and Its Application in the Production of Chitooligomers with Their Antioxidant and Anti-Inflammatory Evaluation. Int. J. Mol. Sci. 2016, 17, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yu, H.; Tian, S.; Yang, H.; Wang, J.; Zhu, W.-Y. Recombinant expression insulin-like growth factor 1 in Bacillus subtilis using a low-cost heat-purification technology. Process Biochem. 2017, 63, 49–54. [Google Scholar] [CrossRef]
- Liu, Y.-L.; Jiang, S.; Ke, Z.-M.; Wu, H.-S.; Chi, C.-W.; Guo, Z.-Y. Recombinant expression of a chitosanase and its application in chitosan oligosaccharide production. Carbohydr. Res. 2009, 344, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, J.; Wang, S. Purification and characterization of the chitosanase from Aeromonas sp. HG08. Afr. J. Biotechnol. 2009, 8, 2830–2834. [Google Scholar]
- Chen, Y.-Y.; Lin, S.-Y.; Yeh, Y.-Y.; Hsiao, H.-H.; Wu, C.-Y.; Chen, S.-T.; Wang, A.H.-J. A modified protein precipitation procedure for efficient removal of albumin from serum. Electrophoresis 2005, 26, 2117–2127. [Google Scholar] [CrossRef]
- Teotia, S.; Lata, R.; Gupta, M.N. Chitosan as a macroaffinity ligand: Purification of chitinases by affinity precipitation and aqueous two-phase extractions. J. Chromatogr. A 2004, 1052, 85–91. [Google Scholar] [CrossRef]
- Mirica, K.A.; Lockett, M.R.; Snyder, P.W.; Shapiro, N.D.; Mack, E.T.; Nam, S.; Whitesides, G.M. Selective Precipitation and Purification of Monovalent Proteins Using Oligovalent Ligands and Ammonium Sulfate. Bioconj. Chem. 2012, 23, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.-Y.; Shrestha, K.L.; Wu, Y.-J.; Tasi, H.-J.; Chen, C.-C.; Yang, J.-M.; Ando, A.; Cheng, C.-Y.; Li, Y.-K. Structural simulation and protein engineering to convert an endo-chitosanase to an exo-chitosanase. Protein Eng. Des. Sel. 2008, 21, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Reddy, N.; Li, Y.; Yang, Y. Alkali-catalyzed low temperature wet crosslinking of plant proteins using carboxylic acids. Biocatal. Bioreact. Des. 2009, 25, 139–146. [Google Scholar] [CrossRef]
- Chumsae, C.; Zhou, L.L.; Shen, Y.; Wohlgemuth, J.; Fung, E.; Burton, R.; Radziejewski, C.; Zhou, Z.S. Discovery of a Chemical Modification by Citric Acid in a Recombinant Monoclonal Antibody. Anal. Chem. 2014, 86, 8932–8936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baehaki, A.; Nurhayati, T.; Palupi, N.S.; Suhartono, M.T. Production and characterization of protease from patogenic bacteria Escherichia coli. J. Chem. Pharm. Res. 2015, 7, 776–780. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|
| Crude extract a | 109 | 2507 | 23 | 100 | 1.0 |
| Selective precipitation | 53 | 2226 | 42 | 89 | 1.8 |
| FPLC | 37 | 2109 | 57 | 84 | 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, C.-Y.; Tsai, C.-H.; Liou, P.-J.; Wang, C.-H. Pilot-Scale Production of Chito-Oligosaccharides Using an Innovative Recombinant Chitosanase Preparation Approach. Polymers 2021, 13, 290. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13020290
Cheng C-Y, Tsai C-H, Liou P-J, Wang C-H. Pilot-Scale Production of Chito-Oligosaccharides Using an Innovative Recombinant Chitosanase Preparation Approach. Polymers. 2021; 13(2):290. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13020290
Chicago/Turabian StyleCheng, Chih-Yu, Chia-Huang Tsai, Pei-Jyun Liou, and Chi-Hang Wang. 2021. "Pilot-Scale Production of Chito-Oligosaccharides Using an Innovative Recombinant Chitosanase Preparation Approach" Polymers 13, no. 2: 290. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13020290