
Screening for Methane Utilizing Mixed Communities with High Polyhydroxybutyrate (PHB) Production Capacity Using Different Design Approaches



Abstract
:1. Introduction
2. Materials and Methods
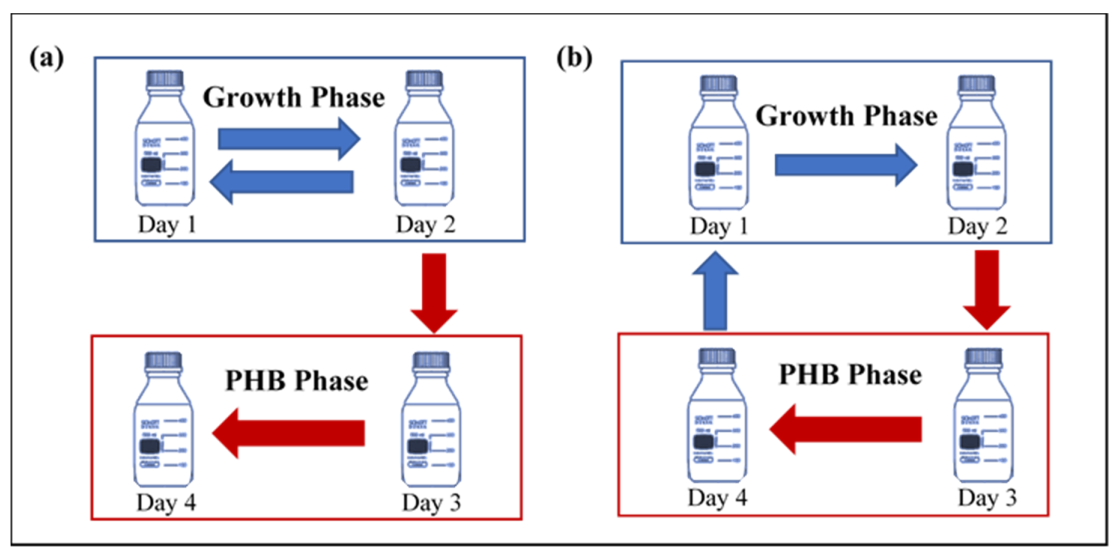
2.1. Setup and Operational Condition
2.2. Media Composition (Inoculum and Synthetic Media Composition)
2.3. Analytical Techniques
2.3.1. OD Measurements
2.3.2. Gas Consumption
2.3.3. PHB Extraction
2.3.4. PHB Quantification
2.4. Microbial Analysis
2.5. Characterization Techniques
2.5.1. Sudan Black B Staining
2.5.2. Nucleic Magnetic Resonance (NMR)
3. Results and Discussion
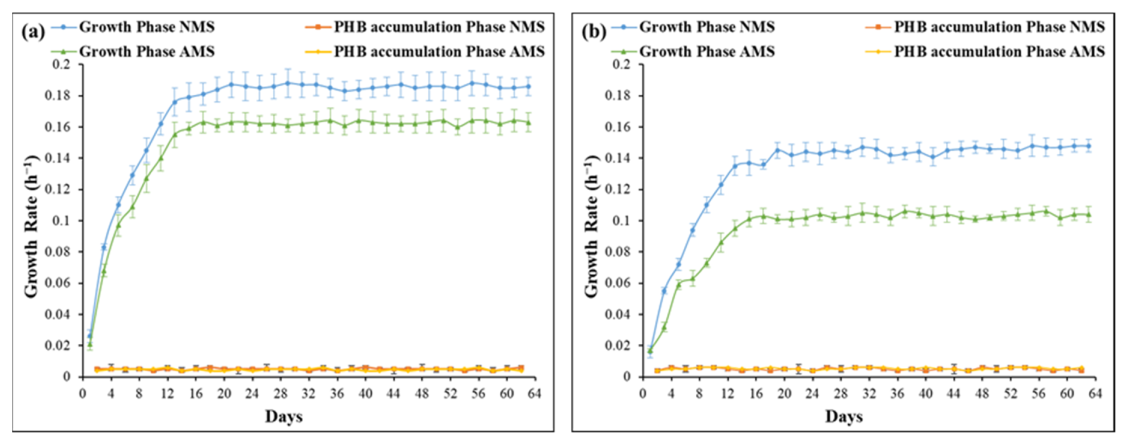
3.1. Growth Rate
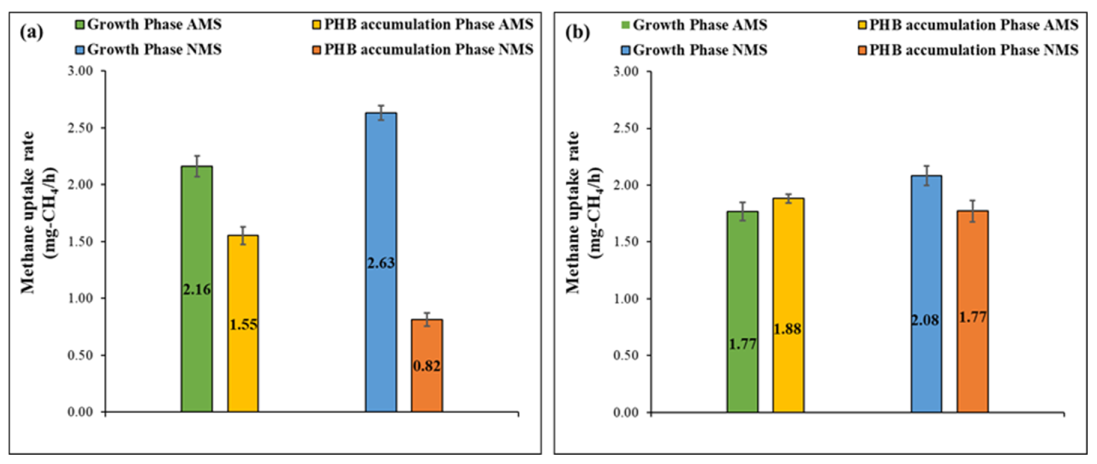
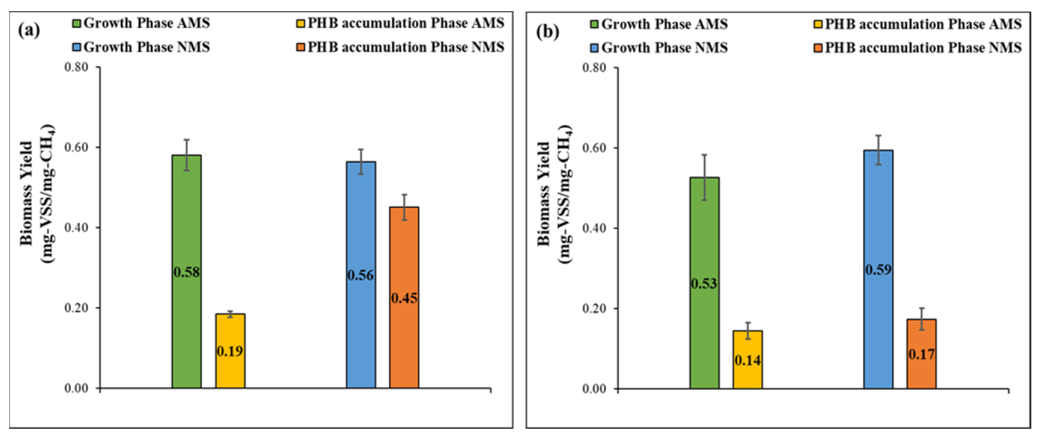
3.2. Methane Uptake and Biomass Yield
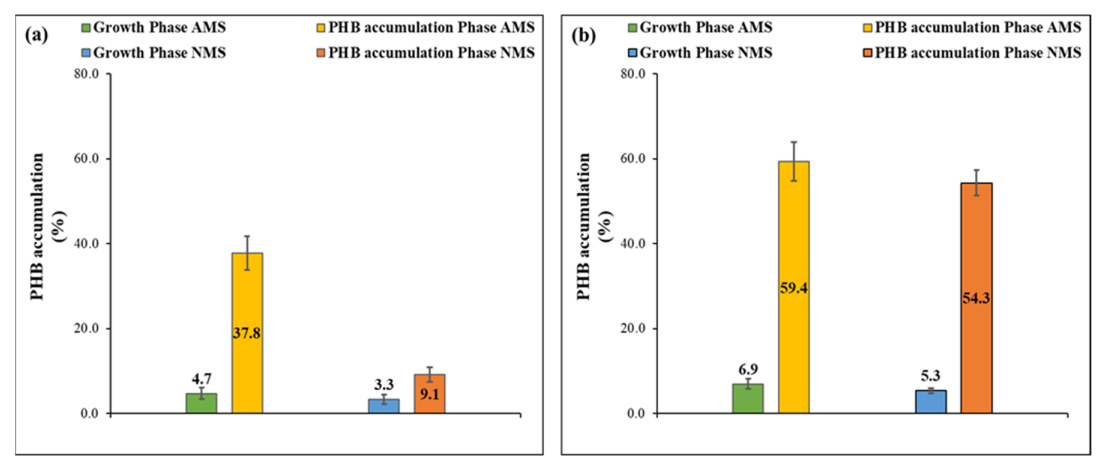
3.3. PHB Accumulation
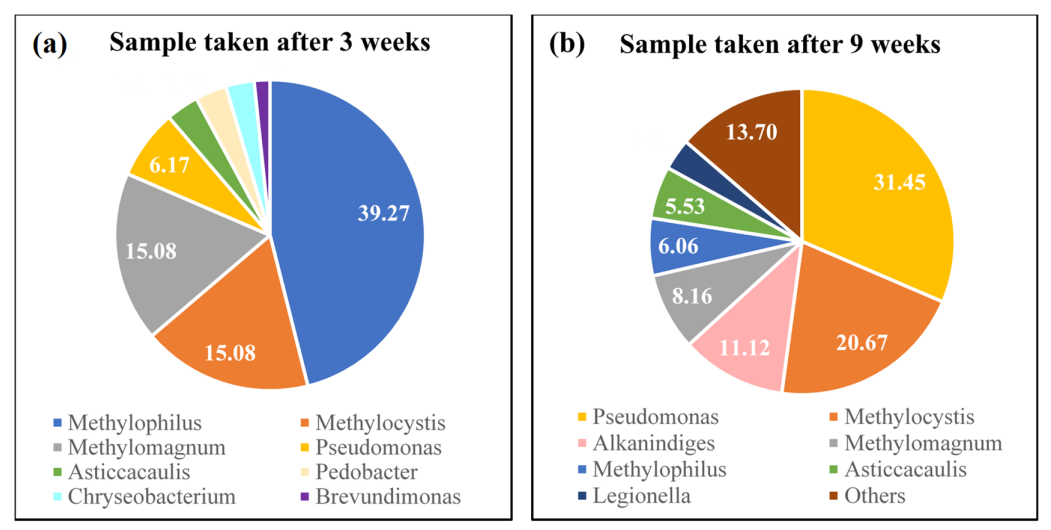
3.4. Microbial Analysis
3.5. PHB Characterization
3.5.1. Sudan Black and Extracted Powder
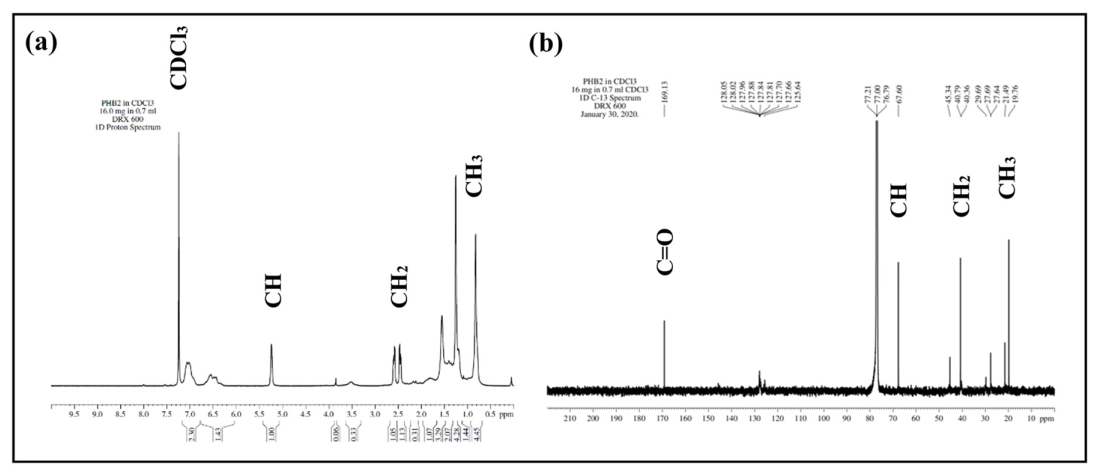
3.5.2. Nuclear Magnetic Resonance (NMR)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, R.; Zimmermann, W. Microbial Enzymes for the Recycling of Recalcitrant Petroleum-based Plastics: How Far are We? Microb. Biotechnol. 2017, 10, 1308–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maser, C. Interactions of Land, Ocean and Humans: A Global Perspective; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Tokiwa, Y.; Calabia, B.P.; Ugwu, C.U.; Aiba, S. Biodegradability of Plastics. Int. J. Mol. Sci. 2009, 10, 3722–3742. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.J.; Dawes, E.A. Occurrence, Metabolism, Metabolic Role, and Industrial Uses of Bacterial Polyhydroxyalkanoates. Microbiol. Rev. 1990, 54, 450–472. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, Z.; Wen, Q.; Zhao, L.; Lee, D.-J.; Yang, L.; Wang, Y. Insights into Feast-Famine Polyhydroxyalkanoate (PHA)-Producer Selection: Microbial Community Succession, Relationships with System Function and Underlying Driving Forces. Water Res. 2018, 131, 167–176. [Google Scholar] [CrossRef]
- Zaheer, M.R.; Kuddus, M. PHB (poly-β-hydroxybutyrate) and Its Enzymatic Degradation. Polym. Adv. Technol. 2018, 29, 30–40. [Google Scholar] [CrossRef]
- Satoh, H.; Mino, T.; Matsuo, T. PHA Production by Activated Sludge. Int. J. Biol. Macromol. 1999, 25, 105–109. [Google Scholar] [CrossRef]
- Shashoua, Y. Conservation of Plastics; Routledge: London, UK, 2012; Volume 53, Number 9. [Google Scholar]
- Khanna, S.; Srivastava, A.K. Statistical Media Optimization Studies for Growth and PHB Production by Ralstonia Eutropha. Process. Biochem. 2005, 40, 2173–2182. [Google Scholar] [CrossRef]
- Son, H.; Park, G.; Lee, S. Growth-associated Production of Poly-β-hydroxybutyrate from Glucose or Alcoholic Distillery Wastewater by Actinobacillus sp. EL-9. Biotechnol. Lett. 1996, 18, 1229–1234. [Google Scholar] [CrossRef]
- Ahn, W.S.; Park, S.J.; Lee, S.Y. Production of poly(3-hydroxybutyrate) from whey by cell recycle fed-batch culture of recombinant Escherichia coli. Biotechnol. Lett. 2001, 23, 235–240. [Google Scholar] [CrossRef]
- Tamer, I.M.; Moo-Young, M.; Chisti, Y. Optimization of Poly(β-hydroxybutyric acid) Recovery from Alcaligenes Latus: Combined Mechanical and Chemical Treatments. Bioprocess. Eng. 1998, 19, 459–468. [Google Scholar] [CrossRef]
- Pieja, A.J.; Sundstrom, E.R.; Criddle, C.S. Poly-3-hydroxybutyrate Metabolism in the Type II Methanotroph Methylocystis Parvus OBBP. Appl. Environ. Microbiol. 2011, 77, 6012–6019. [Google Scholar] [CrossRef] [Green Version]
- AlSayed, A.; Fergala, A.; Khattab, S.; Eldyasti, A. Kinetics of Type I Methanotrophs Mixed Culture Enriched from Waste Activated Sludge. Biochem. Eng. J. 2018, 132, 60–67. [Google Scholar] [CrossRef]
- Asenjo, J.A.; Suk, J.S. Microbial Conversion of Methane into Poly-β-hydroxybutyrate (PHB): Growth and Intracellular Product Accumulation in a Type II Methanotroph. J. Ferment. Technol. 1986, 64, 271–278. [Google Scholar] [CrossRef]
- Hanson, R.S.; Hanson, T.E. Methanotrophic Bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [CrossRef]
- Karthikeyan, O.P.; Chidambarampadmavathy, K.; Cirés, S.; Heimann, K. Review of Sustainable Methane Mitigation and Biopolymer Production. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1579–1610. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, J.; Wang, X.; Zhang, Y. Coupled Effects of Methane Monooxygenase and Nitrogen Source on Growth and Poly-β-hydroxybutyrate (PHB) Production of Methylosinus Trichosporium OB3b. J. Environ. Sci. 2017, 52, 49–57. [Google Scholar] [CrossRef]
- Criddle, C.; Rostkowski, K.H.; Sundstrom, E.R. Process for the Selection of PHB-Producing Methanotrophic Cultures. U.S. Patent 9,062,340, 23 June 2015. [Google Scholar]
- Tays, C.; Guarnieri, M.T.; Sauvageau, D.; Stein, L.Y. Combined Effects of Carbon and Nitrogen Source to Optimize Growth of Proteobacterial Methanotrophs. Front. Microbiol. 2018, 9, 2239. [Google Scholar] [CrossRef]
- Myung, J.; Galega, W.M.; van Nostrand, J.D.; Yuan, T.; Zhou, J.; Criddle, C.S. Long-Term Cultivation of a Stable Methylocystis-Dominated Methanotrophic Enrichment Enabling Tailored Production of Poly(3-hydroxybutyrate-co-3-hydroxyvalerate). Bioresour. Technol. 2015, 198, 811–818. [Google Scholar] [CrossRef]
- Sundstrom, E.R.; Criddle, C.S. Optimization of Methanotrophic Growth and Production of Poly(3-hydroxybutyrate) in a High-Throughput Microbioreactor System. Appl. Environ. Microbiol. 2015, 81, 4767–4773. [Google Scholar] [CrossRef] [Green Version]
- AlSayed, A.; Fergala, A.; Eldyasti, A. Influence of Biomass Density and Food to Microorganisms Ratio on the Mixed Culture Type I Methanotrophs Enriched from Activated Sludge. J. Environ. Sci. 2018, 70, 87–96. [Google Scholar] [CrossRef]
- Teeka, J.; Imai, T.; Reungsang, A.; Cheng, X.; Yuliani, E.; Thiantanankul, J.; Poomipuk, N.; Yamaguchi, J.; Jeenanong, A.; Higuchi, T.; et al. Characterization of Polyhydroxyalkanoates (PHAs) Biosynthesis by Isolated Novosphingobium sp. THA-AIK7 Using Crude Glycerol. J. Ind. Microbiol. Biotechnol. 2012, 39, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Fergala, A.; AlSayed, A.; Eldyasti, A. Factors Affecting the Selection of PHB Accumulating Methanotrophs from Waste Activated Sludge While Utilizing Ammonium as Their Nitrogen Source. J. Chem. Technol. Biotechnol. 2018, 93, 1359–1369. [Google Scholar] [CrossRef]
- García, A.; Segura, D.; Espín, G.; Galindo, E.; Castillo, T.; Peña, C. High Production of Poly-β-hydroxybutyrate (PHB) by an Azotobacter Vinelandii Mutant Altered in PHB Regulation Using a Fed-batch Fermentation Process. Biochem. Eng. J. 2014, 82, 117–123. [Google Scholar] [CrossRef]
- Jiang, Y.; Song, X.; Gong, L.; Li, P.; Dai, C.; Shao, W. High Poly(β-hydroxybutyrate) Production by Pseudomonas Fluorescens A2a5 from Inexpensive Substrates. Enzym. Microb. Technol. 2008, 42, 167–172. [Google Scholar] [CrossRef]
- Bugnicourt, E.; Cinelli, P.; Lazzeri, A.; Alvarez, V. Polyhydroxyalkanoate (PHA): Review of Synthesis, Characteristics, Processing and Potential Applications in Packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Huang, R.; Wang, B.Z.; Bodelier, P.L.E.; Jia, Z.J. Competitive Interactions between Methane- and Ammonia-Oxidizing Bacteria Modulate Carbon and Nitrogen Cycling in Paddy Soil. Biogeosciences 2014, 11, 3353–3368. [Google Scholar] [CrossRef]
- Nyerges, G.; Han, S.K.; Stein, L.Y. Effects of Ammonium and Nitrite on Growth and Competitive Fitness of Cultivated Methanotrophic Bacteria. Appl. Environ. Microbiol. 2010, 76, 5648–5651. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.A.; Nyerges, G.; Kozlowski, J.A.; Poret-Peterson, A.T.; Stein, L.Y.; Klotz, M.G. Model of the Molecular Basis for Hydroxylamine Oxidation and Nitrous Oxide Production in Methanotrophic Bacteria. FEMS Microbiol. Lett. 2011, 322, 82–89. [Google Scholar] [CrossRef]
- de Visscher, A.; van Cleemput, O. Induction of Enhanced CH4 Oxidation in Soils: NH4+ Inhibition Patterns. Soil Biol. Biochem. 2003, 35, 907–913. [Google Scholar] [CrossRef]
- Madigan, M.T.; Bender, K.S.; Buckley, D.H.; Sattley, W.M.; Stahl, D.A. Brock Biology of Microorganisms, 14th ed.; Pearson Education Limited: Boston, MA, USA, 2015. [Google Scholar]
- Arcangeli, J.P.; Arvin, E. Modelling the Growth of a Methanotrophic Biofilm: Estimation of Parameters and Variability. Biodegradation 1999, 10, 177–191. [Google Scholar] [CrossRef]
- Kevbrina, M.V.; Okhapkina, A.A.; Akhlynin, D.S.; Kravchenko, I.K.; Nozhevnikova, A.N.; Gal’chenko, V.F. Growth of Mesophilic Methanotrophs at Low Temperatures. Microbiology 2001, 70, 384–391. [Google Scholar] [CrossRef]
- Fergala, A.; Alsayed, A.; Khattab, S.; Ramirez, M.; Eldyasti, A. Development of Methane-Utilizing Mixed Cultures for the Production of Polyhydroxyalkanoates (PHAs) from Anaerobic Digester Sludge. Environ. Sci. Technol. 2018, 52, 12376–12387. [Google Scholar] [CrossRef]
- Gilman, A.; Laurens, L.M.; Puri, A.W.; Chu, F.; Pienkos, P.T.; Lidstrom, M.E. Bioreactor Performance Parameters for an Industrially-Promising Methanotroph Methylomicrobium Buryatense 5GB1. Microb. Cell Fact. 2015, 14, 182. [Google Scholar] [CrossRef] [Green Version]
- Joergensen, L.; Degn, H. Mass Spectrometric Measurements of Methane and Oxygen Utilization by Methanotrophic Bacteria. FEMS Microbiol. Lett. 1983, 20, 331–335. [Google Scholar] [CrossRef]
- Leak, D.J.; Dalton, H. Growth Yields of Methanotrophs-1. Effect of Copper on the Energetics of Methane Oxidation. Appl. Microbiol. Biotechnol. 1986, 23, 470–476. [Google Scholar] [CrossRef]
- Hölscher, T.; Breuer, U.; Adrian, L.; Harms, H.; Maskow, T. Production of the Chiral Compound (R)-3-hydroxybutyrate by a Genetically Engineered Methylotrophic Bacterium. Appl. Environ. Microbiol. 2010, 76, 5585–5591. [Google Scholar] [CrossRef] [Green Version]
- Pieja, A.J.; Rostkowski, K.H.; Criddle, C.S. Distribution and Selection of Poly-3-Hydroxybutyrate Production Capacity in Methanotrophic Proteobacteria. Microb. Ecol. 2011, 62, 564–573. [Google Scholar] [CrossRef]
- Korotkova, N.; Lidstrom, M.E.; Chistoserdova, L. Identification of Genes Involved in the Glyoxylate Regeneration Cycle in Methylobacterium Extorquens AM1, Including Two New Genes, meaC and meaD. J. Bacteriol. 2005, 187, 1523–1526. [Google Scholar] [CrossRef] [Green Version]
- Korotkova, N.; Lidstrom, M.E. Connection between Poly-β-hydroxybutyrate Biosynthesis and Growth on C1 and C2 Compounds in the Methylotroph Methylobacterium Extorquens AM1. J. Bacteriol. 2001, 183, 1038–1046. [Google Scholar] [CrossRef] [Green Version]
- Latha, D.; Sandya; Shahina; Rekha. Biopolymer Production Using Pseudomonas Sp. (MTCC) and Its Application in Agriculture. Am. J. Biochem. Mol. Biol. 2013, 3, 293–303. [Google Scholar] [CrossRef]
- Yu, J. Pseudomonas Oleovorans-an Overview|ScienceDirect Topics. 2007. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/topics/immunology-and-microbiology/pseudomonas-oleovorans (accessed on 30 March 2020).
- Mozejko-Ciesielska, J.; Szacherska, K.; Marciniak, P. Pseudomonas Species as Producers of Eco-friendly Polyhydroxyalkanoates. J. Polym. Environ. 2019, 27, 1151–1166. [Google Scholar] [CrossRef] [Green Version]
- Phanse, N.; Chincholikar, A.; Patel, B.; Rathore, P.; Vyas, P.; Patel, M. Screening of PHA (Poly Hydroxyalkanoate) Producing Bacteria from Diverse Sources International Journal of Biosciences (IJB). Int. J. Biosci. 2011, 1, 27–32. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Groups | 13C NMR (ppm) | 1H NMR (ppm) |
|---|---|---|
| CH3 | 19.76 | 1.2 |
| CH2 | 40.79 | 2.4–2.6 |
| CH | 67.60 | 5.2 |
| C=O | 169.13 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salem, R.; Soliman, M.; Fergala, A.; Audette, G.F.; ElDyasti, A. Screening for Methane Utilizing Mixed Communities with High Polyhydroxybutyrate (PHB) Production Capacity Using Different Design Approaches. Polymers 2021, 13, 1579. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13101579
Salem R, Soliman M, Fergala A, Audette GF, ElDyasti A. Screening for Methane Utilizing Mixed Communities with High Polyhydroxybutyrate (PHB) Production Capacity Using Different Design Approaches. Polymers. 2021; 13(10):1579. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13101579
Chicago/Turabian StyleSalem, Rana, Moomen Soliman, Ahmed Fergala, Gerald F. Audette, and Ahmed ElDyasti. 2021. "Screening for Methane Utilizing Mixed Communities with High Polyhydroxybutyrate (PHB) Production Capacity Using Different Design Approaches" Polymers 13, no. 10: 1579. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13101579