
Mastoparan, a Peptide Toxin from Wasp Venom Conjugated Fluvastatin Nanocomplex for Suppression of Lung Cancer Cell Growth


Abstract
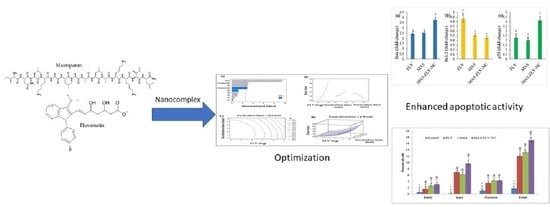
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Formulation and Optimization of MAS-FLV Nanocomplex (MAS-FLV-NC)
2.3. In Vitro Cell Line Studies of MAS-FLV-NC in Lung Cancer Cells (A549 Cells)
2.3.1. IC50 Determination
2.3.2. Apoptotic Activity
2.3.3. Cell Cycle Analysis
2.3.4. Mitochondrial Membrane Potential (MMP)
2.3.5. Determination of Marker Molecules by ELISA
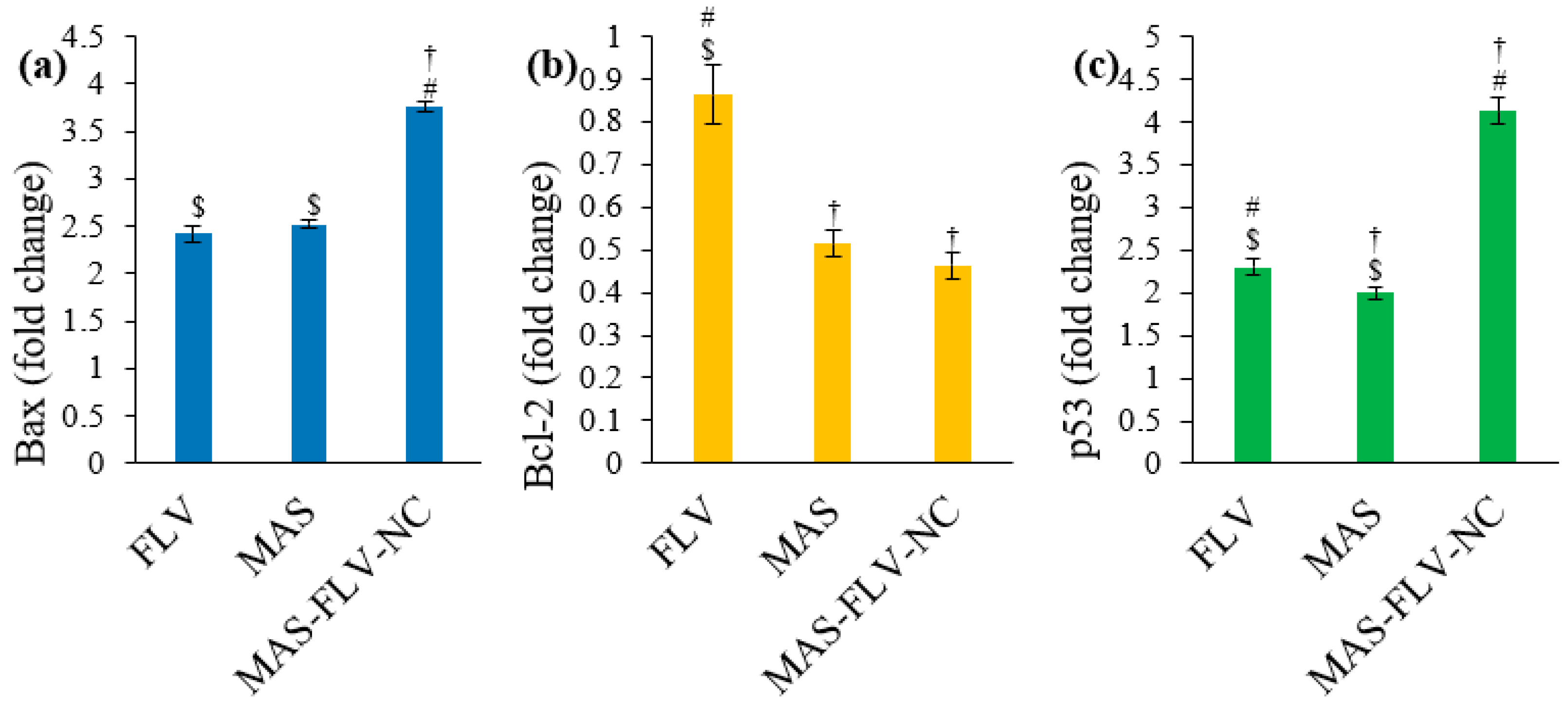
2.3.6. Estimation of Bax, Bcl-2, and p53 Expressions Using RT-PCR
3. Results
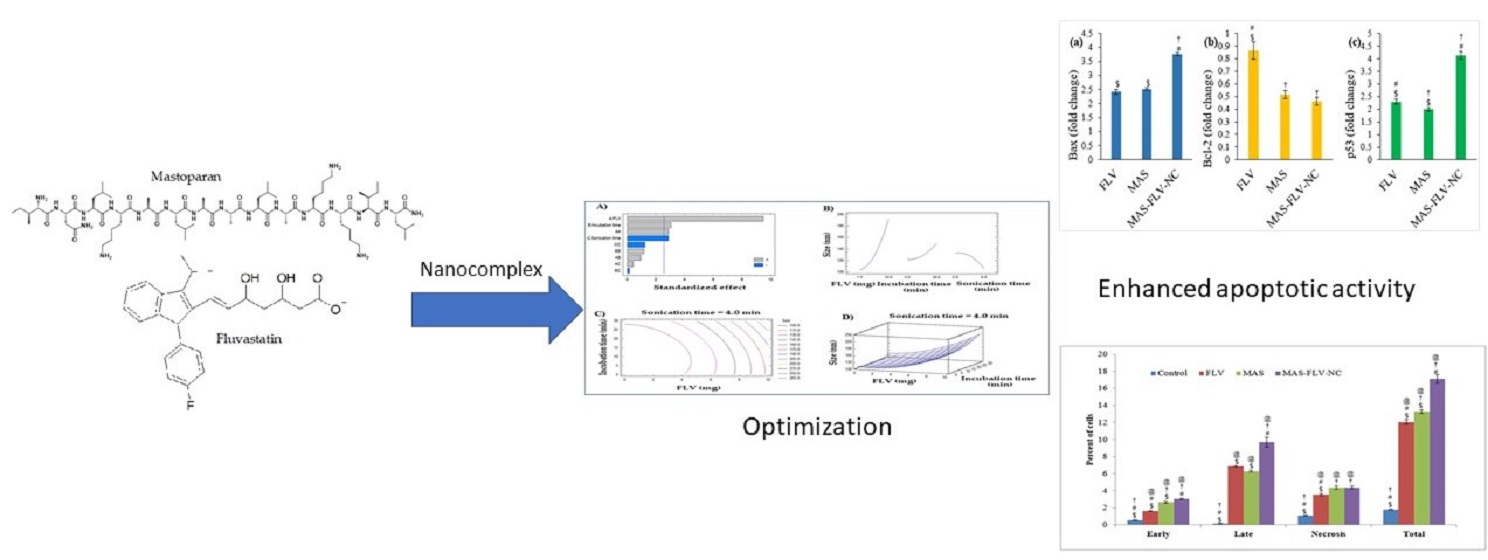
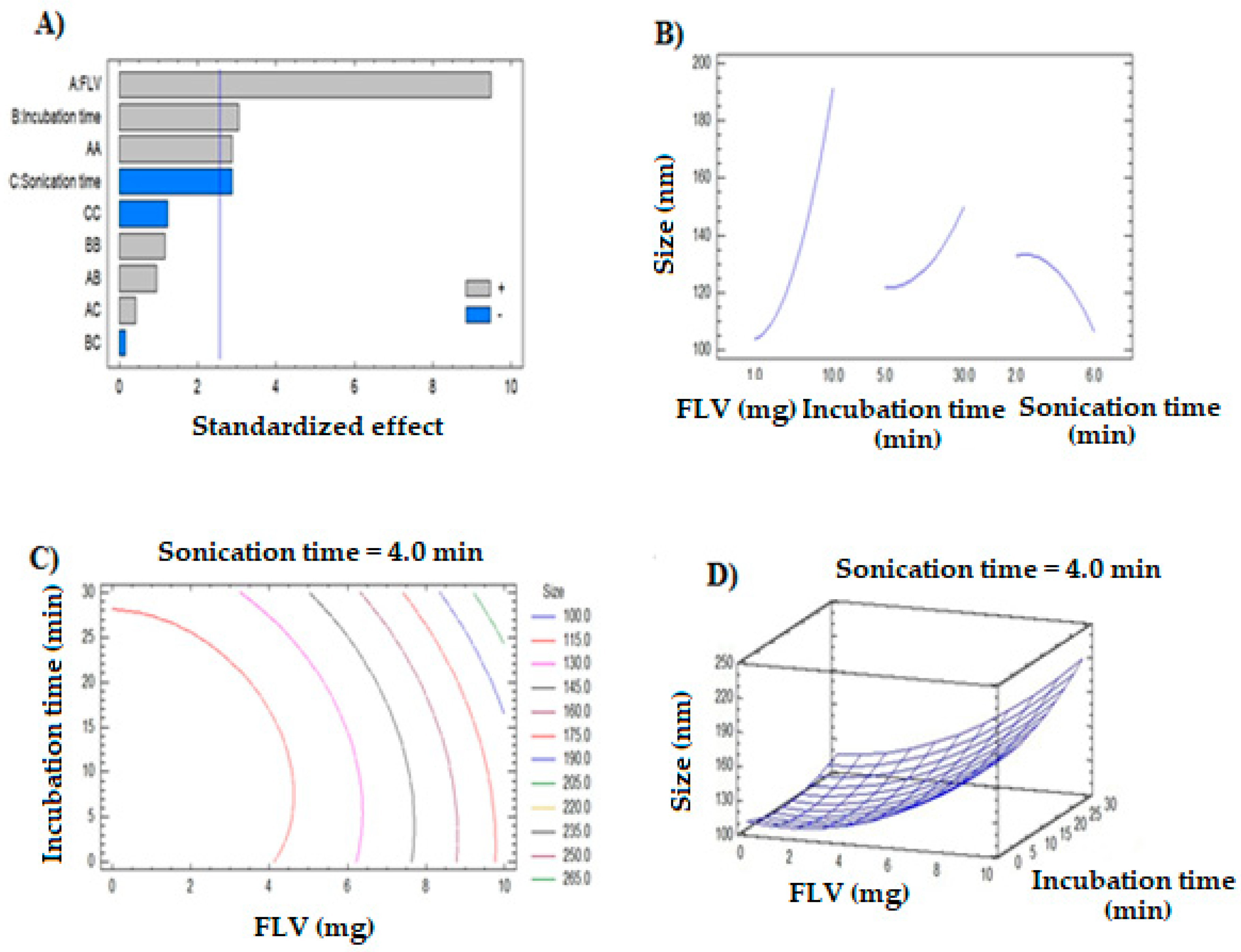
3.1. Formulation and Optimization of MAS-FLV-NC
3.2. In Vitro Cell Line Studies of MAS-FLV-NC in Lung Cancer Cells (A549 Cells)
3.2.1. IC50 Determination
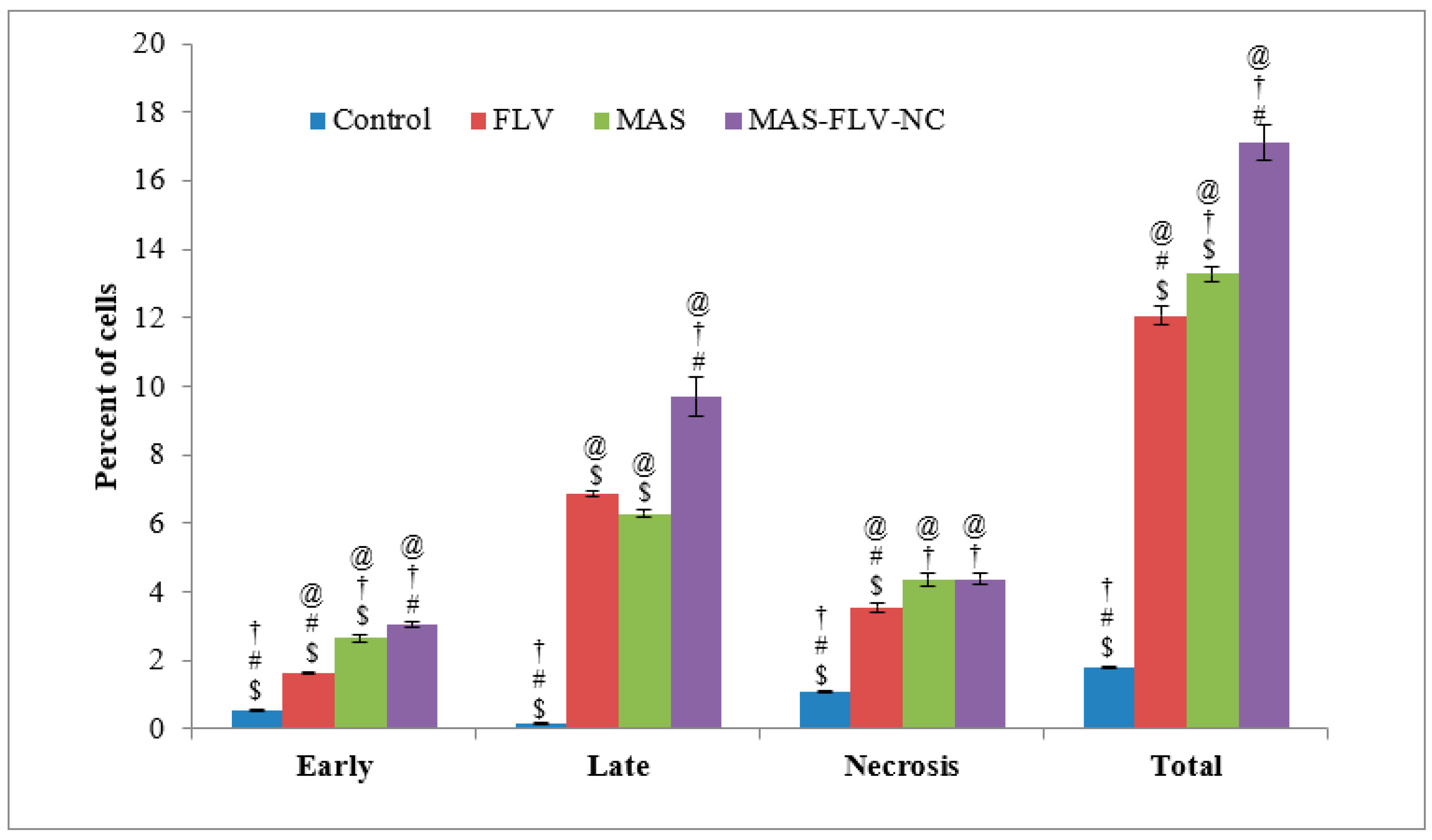
3.2.2. Apoptotic Activity
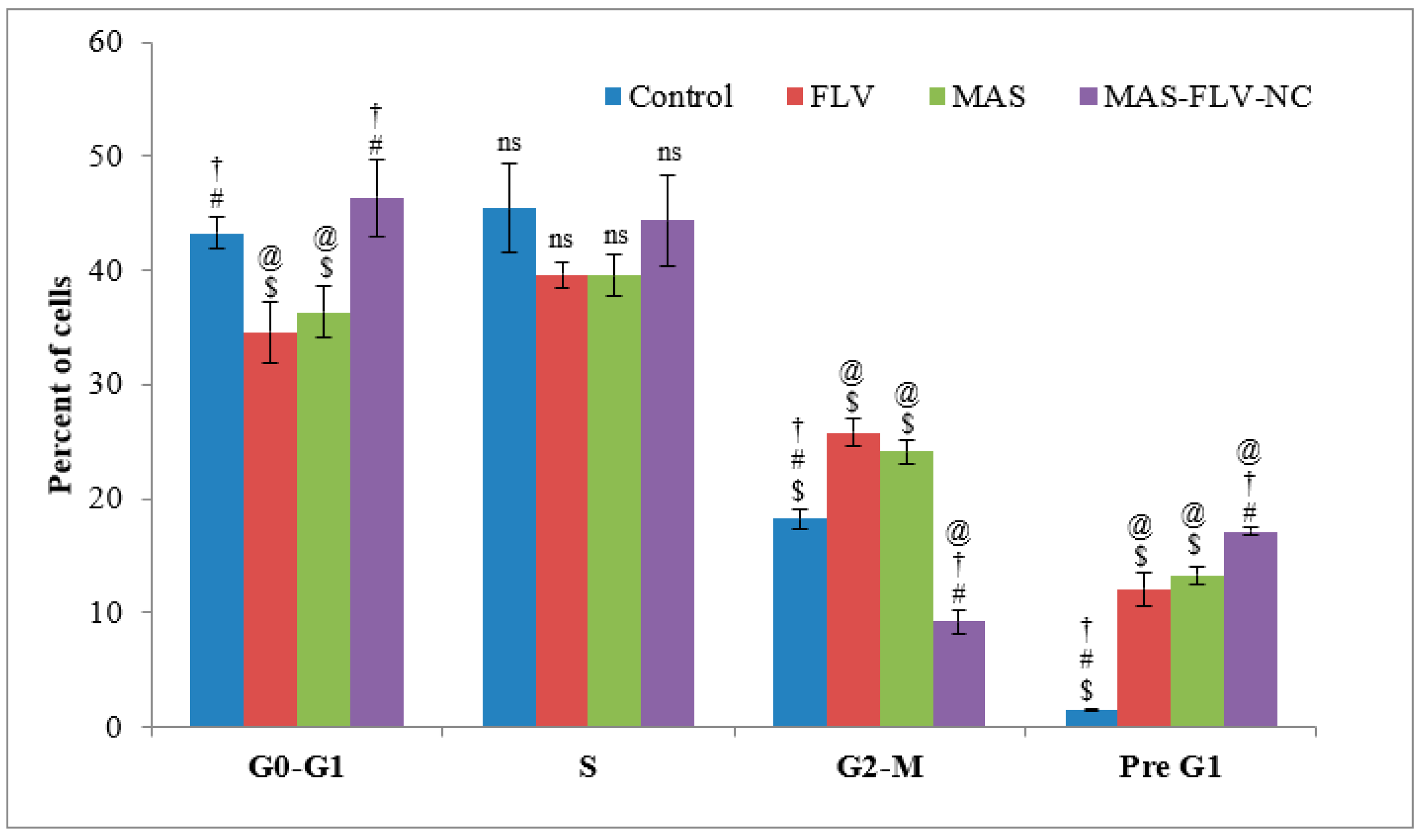
3.2.3. Cell Cycle Analysis
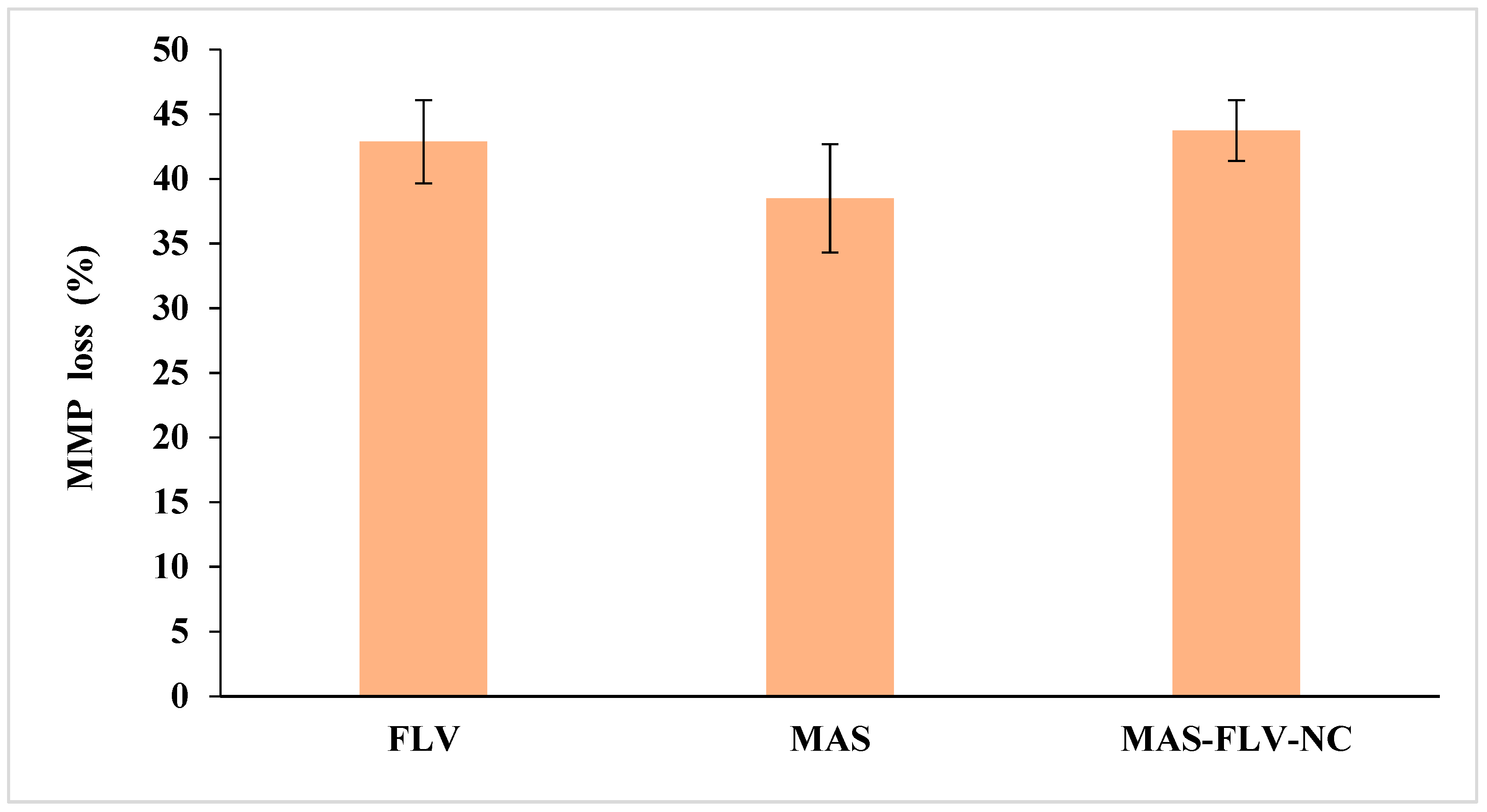
3.2.4. Mitochondrial Membrane Potential (MMP)
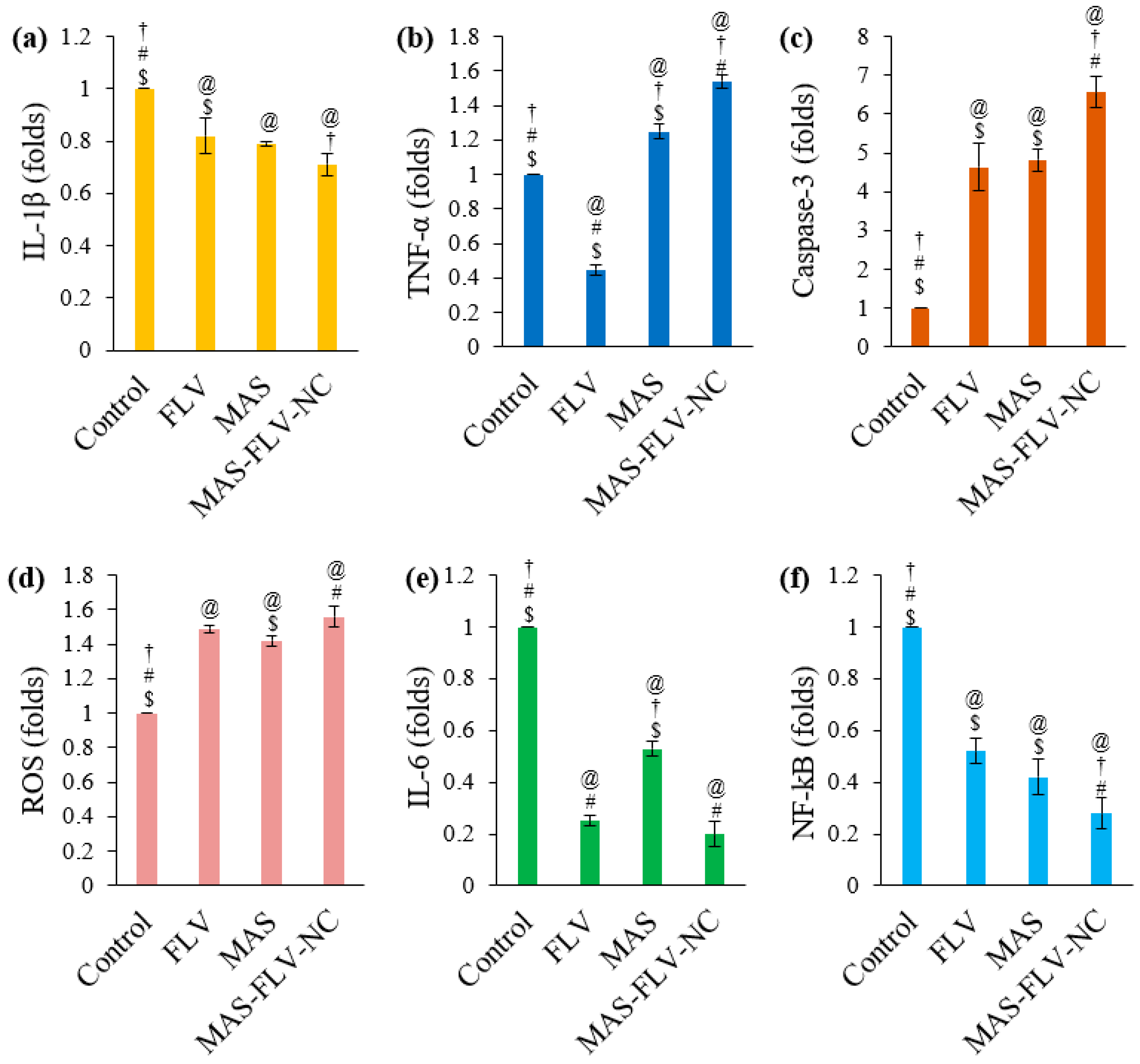
3.2.5. Determination of Marker Molecules by ELISA
3.2.6. Estimation of Bax, Bcl-2, and p53 Expressions Using RT-PCR
4. Discussion
4.1. Optimization and Selection of Best Possible MAS-FLV-NC
4.2. In Vitro Efficacy Studies of MAS-FLV-NC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pirker, R. Conquering lung cancer: Current status and prospects for the future. Pulmonology 2020, 26, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Barta, J.A.; Powell, C.A.; Wisnivesky, J.P. Global Epidemiology of Lung Cancer. Ann. Glob. Health 2019, 85, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, M.R.; Jablons, D.; He, B. Lung cancer therapeutics that target signaling pathways: An update. Expert Rev. Respir. Med. 2010, 4, 631–645. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wang, J.; Xu, D. Cell-penetrating peptides as noninvasive transmembrane vectors for the development of novel multifunctional drug-delivery systems. J. Control. Release 2016, 229, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Sawant, R.R.; Patel, N.R.; Torchilin, V.P. Therapeutic delivery using cell-penetrating peptides. Eur. J. Nanomed. 2013, 5, 141–158. [Google Scholar] [CrossRef]
- Jones, S.; Howl, J. Enantiomer-Specific Bioactivities of Peptidomimetic Analogues of Mastoparan and Mitoparan: Characterization of Inverso Mastoparan as a Highly Efficient Cell Penetrating Peptide. Bioconjug. Chem. 2012, 23, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Ikram, A.; Raza, A.; Saeed, S.; Paracha, R.Z.; Younas, Z.; Khadim, M.T. Therapeutic potential of novel mastoparan-chitosan nanoconstructs against clinical MDR Acinetobacter baumannii: In silico, in vitro and in vivo Studies. Int. J. Nanomed. 2021, 16, 3755–3773. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.P.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int. J. Biol. Sci. 2018, 14, 599–607. [Google Scholar] [CrossRef] [PubMed]
- De Azevedo, R.A.; Figueiredo, C.R.; Ferreira, A.K.; Matsuo, A.L.; Massaoka, M.H.; Girola, N.; Auada, A.V.V.; Farias, C.F.; Pasqualoto, K.F.M.; Rodrigues, C.P.; et al. Mastoparan induces apoptosis in B16F10-Nex2 melanoma cells via the intrinsic mitochondrial pathway and displays antitumor activity in vivo. Peptides 2015, 68, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Hilchie, A.L.; Sharon, A.J.; Haney, E.F.; Hoskin, D.W.; Bally, M.B.; Franco, O.L.; Corcoran, J.A.; Hancock, R.E.W. Mastoparan is a membranolytic anti-cancer peptide that works synergistically with gemcitabine in a mouse model of mammary carcinoma. Biochim. Biophys. Acta. 2016, 1858, 3195–3204. [Google Scholar] [CrossRef]
- Hamid, H.M.A.; Darwish, Z.E.; Elsheikh, S.M.; Mourad, G.M.; Donia, H.M.; Afifi, M.M. Following cytotoxic nanoconjugates from injection to halting the cell cycle machinery and its therapeutic implications in oral cancer. BMC Cancer 2021, 21, 170. [Google Scholar] [CrossRef]
- Irby, D.; Du, C.; Li, F. Lipid–Drug Conjugate for Enhancing Drug Delivery. Mol. Pharm. 2017, 14, 1325–1338. [Google Scholar] [CrossRef] [Green Version]
- Ajithkumar, K.C.; Pramod, K. Doxorubicin-DNA adduct entrenched and motif tethered artificial virus encased in pH-responsive polypeptide complex for targeted cancer therapy. Mater. Sci. Eng. C 2018, 89, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Abioye, A.O.; Chi, G.T.; Kola-Mustapha, A.T.; Ruparelia, K.; Beresford, K.; Arroo, R. Polymer-Drug Nanoconjugate—An Innovative Nanomedicine: Challenges and Recent Advancements in Rational Formulation Design for Effective Delivery of Poorly Soluble Drugs. Pharm. Nanotechnol. 2016, 4, 38–79. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cheetham, A.G.; Angacian, G.; Su, H.; Xie, L.; Cui, H. Peptide–drug conjugates as effective prodrug strategies for targeted delivery. Adv. Drug Deliv. Rev. 2017, 110–111, 112–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Hasan, A.; Babadaei, M.M.N.; Bloukh, S.H.; Edis, Z.; Rasti, B.; Sharifi, M.; Falahati, M. Application of gelatin nanoconjugates as potential internal stimuli-responsive platforms for cancer drug delivery. J. Mol. Liq. 2020, 318, 114053. [Google Scholar] [CrossRef]
- Garwood, E.R.; Kumar, A.S.; Baehner, F.L.; Moore, D.H.; Au, A.; Hylton, N.; Flowers, C.I.; Garber, J.; Lesnikoski, B.-A.; Hwang, E.S.; et al. Fluvastatin reduces proliferation and increases apoptosis in women with high grade breast cancer. Breast Cancer Res. Treat. 2010, 119, 137–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Bai, R.; Wang, Q.; Wang, K.; Li, X.; Liu, K.; Ryu, J.; Wang, T.; Chang, X.; Ma, W.; et al. Fluvastatin Inhibits HMG-CoA Reductase and Prevents Non-Small Cell Lung Carcinogenesis. Cancer Prev. Res. 2019, 12, 837–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Su, Z.; DeWitt, J.P.; Xie, L.; Chen, Y.; Li, X.; Han, L.; Li, D.; Xia, J.; Zhang, Y.; et al. Fluvastatin Prevents Lung Adenocarcinoma Bone Metastasis by Triggering Autophagy. EBioMedicine 2017, 19, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Al-Wahaibi, L.H.; Al-Saleem, M.S.M.; Ahmed, O.A.A.; Fahmy, U.A.; Alhakamy, N.A.; Eid, B.G.; Abdel-Naim, A.B.; Abdel-Mageed, W.M.; AlRasheed, M.M.; Shazly, G.A. Optimized Conjugation of Fluvastatin to HIV-1 TAT Displays Enhanced Pro-Apoptotic Activity in HepG2 Cells. Int. J. Mol. Sci. 2020, 21, 4138. [Google Scholar] [CrossRef]
- Zein, R.; Sharrouf, W.; Selting, K. Physical Properties of Nanoparticles That Result in Improved Cancer Targeting. J. Oncol. 2020, 2020, 5194780. [Google Scholar] [CrossRef] [PubMed]
- Politis, S.N.; Colombo, P.; Colombo, G.; Rekkas, D.M. Design of experiments (DoE) in pharmaceutical development. Drug Dev. Ind. Pharm. 2017, 43, 889–901. [Google Scholar] [CrossRef]
- Hsiao, K.Y.; Wu, Y.-J.; Liu, Z.N.; Chuang, C.W.; Huang, H.H.; Kuo, S.M. Anticancer Effects of Sinulariolide-Conjugated Hyaluronan Nanoparticles on Lung Adenocarcinoma Cells. Molecules 2016, 21, 297. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Chen, Q.; Ding, T.; Sun, J. SiO2 nanoparticle-induced impairment of mitochondrial energy metabolism in hepatocytes directly and through a Kupffer cell-mediated pathway in vitro. Int. J. Nanomed. 2014, 9, 2891–2903. [Google Scholar] [CrossRef] [Green Version]
- Eimani, B.G.; Sanati, M.H.; Houshmand, M.; Ataei, M.; Akbarian, F.; Shakhssalim, N. Expression and prognostic significance of bcl-2 and bax in the progression and clinical outcome of transitional bladder cell carcinoma. Cell J. 2014, 15, 356–363. [Google Scholar]
- Qi, X.-F.; Zheng, L.; Lee, K.-J.; Kim, D.-H.; Kim, C.-S.; Cai, D.-Q.; Wu, Z.; Qin, J.-W.; Yu, Y.-H.; Kim, S.-K. HMG-CoA reductase inhibitors induce apoptosis of lymphoma cells by promoting ROS generation and regulating Akt, Erk and p38 signals via suppression of mevalonate pathway. Cell Death Dis. 2013, 4, e518. [Google Scholar] [CrossRef] [Green Version]
- Jerome, K.R.; Sloan, D.D.; Aubert, M. Measurement of CTL-induced cytotoxicity: The caspase 3 assay. Apoptosis 2003, 8, 563–571. [Google Scholar] [CrossRef]
- Ngkelo, A.; Meja, K.; Yeadon, M.; Adcock, I.; Kirkham, P.A. LPS induced inflammatory responses in human peripheral blood mononuclear cells is mediated through NOX4 and Giα dependent PI-3kinase signalling. J. Inflamm. 2012, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Loppnow, H.; Zhang, L.; Buerke, M.; Lautenschläger, M.; Chen, L.; Frister, A.; Schlitt, A.; Luther, T.; Song, N.; Hofmann, B.; et al. Statins potently reduce the cytokine-mediated IL-6 release in SMC/MNC cocultures. J. Cell. Mol. Med. 2011, 15, 994–1004. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Fahmy, U.A.; Badr-Eldin, S.M.; Ahmed, O.A.A.; Asfour, H.Z.; Aldawsari, H.M.; Algandaby, M.M.; Eid, B.G.; Abdel-Naim, A.B.; Awan, Z.A.; et al. Optimized Icariin Phytosomes Exhibit Enhanced Cytotoxicity and Apoptosis-Inducing Activities in Ovarian Cancer Cells. Pharmaceutics 2020, 12, 346. [Google Scholar] [CrossRef]
- Shinde, G.; Shiyani, S.; Shelke, S.; Chouthe, R.; Kulkarni, D.; Marvaniya, K. Enhanced brain targeting efficiency using 5-FU (fluorouracil) lipid-drug conjugated nanoparticles in brain cancer therapy. Prog. Biomater. 2020, 9, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, Y.; Parikh, A.; Joyce, P.; Chung, R.; Liu, L.; Afinjuomo, F.; Hayball, J.D.; Petrovsky, N.; Barclay, T.G.; et al. Doxorubicin-Loaded Delta Inulin Conjugates for Controlled and Targeted Drug Delivery: Development, Characterization, and In Vitro Evaluation. Pharmaceutics 2019, 11, 581. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, A.; Alayoubi, A.; El-Malah, Y.; Nazzal, S. Modeling the effect of sonication parameters on size and dispersion temperature of solid lipid nanoparticles (SLNs) by response surface methodology (RSM). Pharm. Dev. Technol. 2014, 19, 342–346. [Google Scholar] [CrossRef]
- Md, S.; Alhakamy, N.A.; Aldawsari, H.M.; Husain, M.; Kotta, S.; Abdullah, S.T.; Fahmy, U.A.; Alfaleh, M.A.; Asfour, H.Z. Formulation design, statistical optimization, and in vitro evaluation of a naringenin nanoemulsion to enhance apoptotic activity in a549 lung cancer cells. Pharmaceuticals 2020, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Black, K.A.; McFarland, R.D.; Grisham, J.W.; Smith, G.J. S-phase block and cell death in human lymphoblasts exposed to benzo[a]pyrene diol epoxide or N-acetoxy-2-acetylaminofluorene. Toxicol. Appl. Pharmacol. 1989, 97, 463–472. [Google Scholar] [CrossRef]
- Sakamuru, S.; Attene-Ramos, M.S.; Xia, M. Mitochondrial Membrane Potential Assay. Methods Mol. Biol. 2016, 1473, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Alhakamy, N.A.; Ahmed, O.A.A.; Fahmy, U.A.; Md, S. Development and in vitro evaluation of 2-methoxyestradiol loaded polymeric micelles for enhancing anticancer activities in prostate cancer. Polymers 2021, 13, 884. [Google Scholar] [CrossRef]
- Bent, R.; Moll, L.; Grabbe, S.; Bros, M. Interleukin-1 Beta-A Friend or Foe in Malignancies? Int. J. Mol. Sci. 2018, 19, 2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-R.; Li, J.-J.; Huang, C.-X.; Jiang, H. Fluvastatin inhibits the expression of tumor necrosis factor-+A7E and activation of nuclear factor-+A7o-B in human endothelial cells stimulated by C-reactive protein. Clin. Chim. Acta 2005, 353, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Rocha, T.; de Barros, L.L.S.; Fontana, K.; de Souza, B.M.; Palma, M.S.; da Cruz-Höfling, M.A. Inflammation and apoptosis induced by mastoparan Polybia-MPII on skeletal muscle. Toxicon 2010, 55, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.-M.; Chou, T.-C.; Ding, Y.-A.; Li, M.-L. Stimulation of TNF-α, IL-1β and nitrite release from mouse cultured spleen cells and lavaged peritoneal cells by mastoparan M. Immunol. Cell Biol. 1999, 77, 476–482. [Google Scholar] [CrossRef]
- Nishikawa, H.; Kitani, S. Inhibitory effect of ganglioside on mastoparan-induced cytotoxicity and degranulation in lipid raft of connective tissue type mast cell. J. Biochem. Mol. Toxicol. 2011, 25, 158–168. [Google Scholar] [CrossRef]
- Cairns, J.A.; Guy, G.R.; Tan, Y.H. Interleukin-6 regulates the cytotoxic effect of tumour necrosis factor on U937 cells. Immunology 1992, 75, 669–673. [Google Scholar] [PubMed]
- Alhakamy, N.A.; Ahmed, O.A.A.; Fahmy, U.A.; Shadab, M. Apamin-conjugated alendronate sodium nanocomplex for management of pancreatic cancer. Pharmaceuticals 2021, 14, 729. [Google Scholar] [CrossRef] [PubMed]
- Rathinaraj, P.; Muthusamy, G.; Prasad, N.R.; Gunaseelan, S.; Kim, B.; Zhu, S. Folate–Gold–Bilirubin Nanoconjugate Induces Apoptotic Death in Multidrug-Resistant Oral Carcinoma Cells. Eur. J. Drug Metab. Pharm. 2020, 45, 285–296. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Factor Values | ||
|---|---|---|---|
| Factor A: FLV Amount (mg) | Factor B: Incubation Time (min) | Factor C: Sonication Time (min) | |
| 1 | 5.5 | 5 | 2 |
| 2 | 10 | 17.5 | 2 |
| 3 | 5.5 | 17.5 | 4 |
| 4 | 5.5 | 17.5 | 4 |
| 5 | 5.5 | 30 | 2 |
| 6 | 1 | 5 | 4 |
| 7 | 1 | 17.5 | 6 |
| 8 | 5.5 | 30 | 6 |
| 9 | 10 | 5 | 4 |
| 10 | 10 | 30 | 4 |
| 11 | 5.5 | 5 | 6 |
| 12 | 1 | 30 | 4 |
| 13 | 1 | 17.5 | 2 |
| 14 | 10 | 17.5 | 6 |
| 15 | 5.5 | 17.5 | 4 |
| Run | Independent Factors | Dependent Factor | |||
|---|---|---|---|---|---|
| Factor A: FLV Amount (mg) | Factor B: Incubation Time (min) | Factor C: Sonication Time (min) | Response 1: Mean Particle Size (nm) | ||
| Observed | Predicted | ||||
| 1 | 5.5 | 5 | 2 | 132 | 125.8 |
| 2 | 10 | 17.5 | 2 | 191 | 193.9 |
| 3 | 5.5 | 17.5 | 4 | 129 | 128.0 |
| 4 | 5.5 | 17.5 | 4 | 127 | 128.0 |
| 5 | 5.5 | 30 | 2 | 145 | 156.0 |
| 6 | 1 | 5 | 4 | 90 | 103.9 |
| 7 | 1 | 17.5 | 6 | 82 | 79.1 |
| 8 | 5.5 | 30 | 6 | 121 | 127.3 |
| 9 | 10 | 5 | 4 | 176 | 179.4 |
| 10 | 10 | 30 | 4 | 234 | 220.1 |
| 11 | 5.5 | 5 | 6 | 112 | 101.0 |
| 12 | 1 | 30 | 4 | 123 | 119.6 |
| 13 | 1 | 17.5 | 2 | 119 | 111.4 |
| 14 | 10 | 17.5 | 6 | 165 | 172.6 |
| 15 | 5.5 | 17.5 | 4 | 128 | 128.0 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F-Ratio | p-Value |
|---|---|---|---|---|---|
| A: FLV | 15488.0 | 1 | 15488.0 | 89.76 | 0.0002 |
| B: Incubation time | 1596.13 | 1 | 1596.13 | 9.25 | 0.0287 |
| C: Sonication time | 1431.13 | 1 | 1431.13 | 8.29 | 0.0346 |
| A2 | 1440.23 | 1 | 1440.23 | 8.35 | 0.0342 |
| AB | 156.25 | 1 | 156.25 | 0.91 | 0.3850 |
| AC | 30.25 | 1 | 30.25 | 0.18 | 0.6928 |
| B2 | 236.308 | 1 | 236.308 | 1.37 | 0.2946 |
| BC | 4.0 | 1 | 4.0 | 0.02 | 0.8849 |
| C2 | 266.769 | 1 | 266.769 | 1.55 | 0.2688 |
| Total error | 862.75 | 5 | 172.55 | -- | -- |
| Total (corr.) | 21574.9 | 14 | -- | -- | -- |
| Factor | Low | High | Optimum |
|---|---|---|---|
| FLV (mg) | 1.0 | 10.0 | 1.00001 |
| Incubation time (min) | 5.0 | 30.0 | 12.1339 |
| Sonication time (min) | 2.0 | 6.0 | 6.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhakamy, N.A.; Ahmed, O.A.A.; Md, S.; Fahmy, U.A. Mastoparan, a Peptide Toxin from Wasp Venom Conjugated Fluvastatin Nanocomplex for Suppression of Lung Cancer Cell Growth. Polymers 2021, 13, 4225. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13234225
Alhakamy NA, Ahmed OAA, Md S, Fahmy UA. Mastoparan, a Peptide Toxin from Wasp Venom Conjugated Fluvastatin Nanocomplex for Suppression of Lung Cancer Cell Growth. Polymers. 2021; 13(23):4225. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13234225
Chicago/Turabian StyleAlhakamy, Nabil A., Osama A. A. Ahmed, Shadab Md, and Usama A. Fahmy. 2021. "Mastoparan, a Peptide Toxin from Wasp Venom Conjugated Fluvastatin Nanocomplex for Suppression of Lung Cancer Cell Growth" Polymers 13, no. 23: 4225. https://0-doi-org.brum.beds.ac.uk/10.3390/polym13234225