
Protective Effects of Chlorogenic Acid against Carbon Tetrachloride-Induced Hepatotoxicity in Mice


1
Department of Optometry, Da-Yeh University, Changhua 515006, Taiwan
2
Department of Applied Cosmetology, National Tainan Junior College of Nursing, Tainan 700, Taiwan
*
Author to whom correspondence should be addressed.
Processes 2022, 10(1), 31; https://0-doi-org.brum.beds.ac.uk/10.3390/pr10010031
Submission received: 3 November 2021
/
Revised: 16 December 2021
/
Accepted: 21 December 2021
/
Published: 24 December 2021
(This article belongs to the Special Issue Antioxidant Properties of Medicinal Plants)
Abstract
:The protective effects of chlorogenic acid (CGA) against liver injury were evaluated by its reduction in carbon tetrachloride (CCl4)-induced hepatic damage in ICR mice. The animals were orally given CGA (60, 100, and 200 mg/kg, respectively) or silymairn (200 mg/kg) daily with 0.3% CCl4 administration (3 mL/kg, dissolved in olive oil) after medicament treatment on the 7th day. Compared with the normal group, CCl4 caused severe impairment in liver according to the evidence of significant reduction in the level of total albumin and expansion (p < 0.05) of the activities in aspartate aminotransferase (AST) and alanine aminotransferase (ALT), cholesterol, triglyceride (TG), and total albumin in serum, decreased the level of glutathione (GSH), and diminished the activities of catalase, superoxide dismutase (SOD), glutathione reductase (GSH-Rd), and glutathione peroxidase (GSH-Px) in liver while increasing the level of hepatic thiobarbituric acid-reactive substances (TBARS). However, oral administration of CGA or silymarin could significantly (p < 0.05) decrease the serum levels of AST, ALT, cholesterol, TG, and total albumin and elevated the serum total albumin and the activities of GSH, catalase, SOD, GSH-Rd, and GSH-Px while leading to decline the TBARS in liver compared with CCl4-intoxicated group. Moreover, histopathology displayed that CGA decreased the formation of lesions in liver resulted from CCl4. The outcomes indicate that CGA shows the efficiency hepatoprotective consequences for CCl4-incited liver injuries in mice by the elevation of the activities of antioxidant enzymes and hindrance of lipid peroxidation.
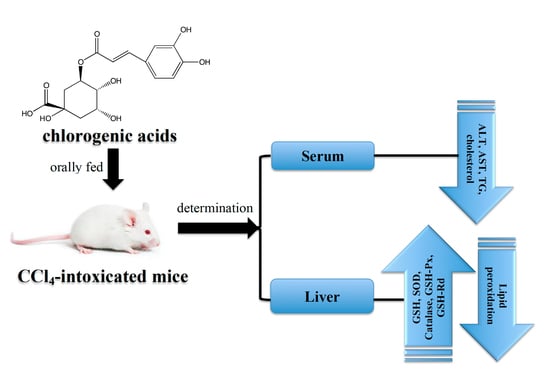
1. Introduction
There are many hepatotoxins in life, such as ethanol in alcoholic beverages, acetaminophen in antipyretic and analgesic drugs, and carbon tetrachloride (CCl4) in chemicals, which could cause liver damage and are characterized by causing various degrees of degeneration and death of hepatocytes [1]. Reactive oxygen species (ROS), a highly aggressive and unstable free radical including superoxide anions and hydroxyl radicals, plays a vital role in the pathogenesis and progression of liver diseases. It has been confirmed to be involved in the hepatotoxicity caused by CCl4 [2,3]. Many studies suggested that CCl4 has been proven to be a hepatoxin that effectively induces liver damage in hepatopathy animal models, and has been widely used in experiments [4,5]. The conjugated combination of trichloromethyl radicals, a metabolite of CCl4 in the liver, with hepatocyte proteins is the initial step of the liver injury process, which subsequently results in lipid peroxidation of membrane, and ultimately to hepatocyte necrosis [6]. It is known that natural antioxidants have specific functions to scavenge ROS and diminish lipid peroxidation, so they can effectively prevent liver pathologies related to oxidative stress [2,7]. A main defense strategy of effective prevention and treatment liver pathologies include diminishing the manufacture of reactive metabolites and inhibiting lipid peroxidation by promoting the levels of effective member of the antioxidant defense system, e.g., catalase, glutathione (GSH), and superoxide dismutase (SOD) [8,9,10].
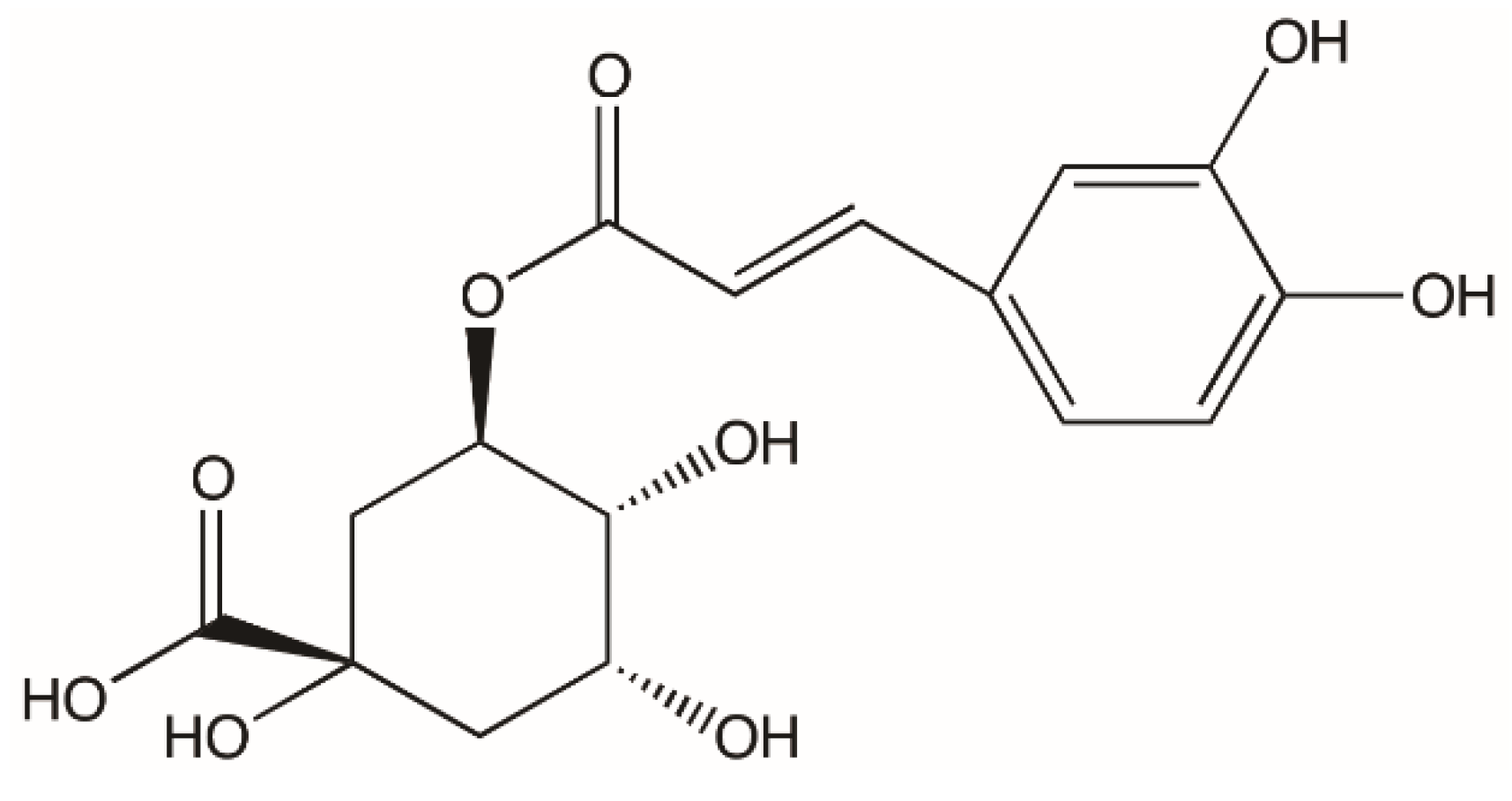
Polyphenols are considered to be helpful to human health and can exert various biological effectiveness, such as diminishing excessive metal chelation, free radicals, increasing enzyme activities, and regulating the pathways of signal transduction [11]. Chlorogenic acid (CGA) (Figure 1), a phenolic compound produced by the esterification of cinnamic acid and (-)-quinic acid, is widely found in vegetables, fruits, and plants; a main intake of CGA in the diet are from coffee beans [12,13,14]. CGA has been confirmed to exhibit good biological benefits such as anti-mutagenic, anti-cancer, and antioxidant activities in vitro, as its structure has vicinal hydroxyl groups on aromatic residues [15]. In the past few years, CGA has been proven to be associated with reducing the relative risk of many diseases, such as diabetes type 2, cardiovascular disorders, and Alzheimer’s disease [16,17,18] and contribute to health benefits including antibacterial and anti-inflammatory activity [19,20]. In addition, it has also been found in animal experiments that their lactones could enhance the effect of insulin [21].
Recent studies reflected that increase in oxidative stress is one of the principal pathogenesis of liver disorder. It is confirmed from many documents that CGA has good biological benefits and excellent antioxidant activity, which is sufficient to reduce oxidative stress. Therefore, we hypothesized that the administration of CGA may be able to protect the liver by enhancing the antioxidant defense system in vivo and thereby combating CCl4 mediated hepatotoxicity in experimental animals. In this study, mice were orally gavage daily with CGA or silymarin accompanied by CCl4. Serum biochemical values, including AST, ALT, cholesterol, TG, and total albumin, and the antioxidant status in liver, such as GSH content, antioxidant enzymes activities, and TBARS content were analyzed to display liver damage in experimental animals. The grade of liver injury caused by CCl4 was also observed by histopathological examination.
2. Materials and Methods
2.1. Chemicals
Carbon tetrachloride (reagent grade, 99.9%), silymarin (≥95%), and CGA were acquired from the Sigma Chemical Co. (St. Louis, MO, USA). All the other analytical grade chemicals and reagents used in this study were received from local chemical companies.
2.2. Animals
The male ICR mice (20 ± 2 g) were purchased from the Animal Department of BioLASCO Taiwan Company and were permitted to isolation and adapt for seven days before experimentation. Mice were taken care of under standard research facility states of a 12 h light-dark cycle in a temperature of 23 ± 2 °C and relative humidity of 50 ± 5% controlled room. All mice had free access to food and water. Our Institutional Animal Care and Use Committee approved all animal experiments in this study (IACUC protocol ORDR-AS-2015-P011). The animals were cared for in accordance to institutional ethical guidelines.
2.3. Treatment
Sixty mice were randomly divided into six groups (ten mice were in each group): (I) normal control, (II) CCl4 group, (III) silymarin + CCl4 group, (IV) CGA (60 mg/kg) + CCl4 group, (V) CGA (100 mg/kg) + CCl4 group, and (VI) CGA (200 mg/kg) + CCl4 group. Group I was given normal saline for 7 consecutive days. For stimulating acute hepatotoxicity in this animal experiment, mice of Groups II, III, IV, V, and VI were injected intraperitoneally (i.p.) with 0.3% CCl4 (3 mL/kg, dissolved in olive oil) after medicament treatment on the 7th day. Group II was only given CCl4. Group III was given silymarin (200 mg/kg) daily for 7 consecutive days. The CGA was dissolved in normal saline and orally administrated to Groups IV, V, and VI at dosages of 60, 100, and 200 mg/kg, respectively, for 7 consecutive days. All animals were sacrificed at the end of experiment and blood was collected into heparinized tubes (50 U/mL) for evaluation of serum AST, ALT, cholesterol, and TG. The liver samples were collected for biochemical assessments and histopathologic analysis.
2.4. Measurement of Serum AST, ALT, Cholesterol, and TG
In order to assess liver damage, the activities of AST, ALT, cholesterol, TG, and total albumin were measured in serum of mice by commercially available test kits (Randox Laboratories Ltd., Crumlin, UK).
2.5. Measurement of GSH, Catalase, SOD, GSH-Rd, and GSH-Px
The homogenates from liver were homogenized in cold Tris buffer (5 mmol/L containing 2 mmol/L EDTA, pH 7.4, 4 °C) utilizing a homogenizer. The homogenates were centrifuged at 10,000 rpm for 10 min at 4 °C. The supernatants were collected and used immediately for the measurement of the activities of GSH, catalase, SOD, GSH-Rd, and GSH-Px following the manufacturer’s instructions on the Randox Laboratories Ltd. (Crumlin, UK) kit.
2.6. Measurement of Lipid Peroxidation
The amount of TBARS in liver was measured to quantity the lipid peroxidation [22]. The TBARS, an index of lipid peroxidation, was quantitated by reaction with malondialdehyde (MDA) and thiobarbituric acid (TBA). Briefly, the homogenates originating from livers were mixed with TBA reagent. The response blends were set in a boiling water shower for half an hour and centrifuged at 1811× g for 5 min. The supernatants were collected and measured via the absorbance at 535 nm.
2.7. Histopathological Evaluation
The histopathological section was commissioned to National Chung Hsing University in Taiwan to make and perform blind reading. Before performing the commissioned histopathology, we coded the samples in the form of serial numbers, so the organization commissioned to conduct the tissue section test did not know the grouping of all the samples in this experiment. After the interpretation of the samples’ histopathology were completed, they were sent back to the investigator, and the investigator would analyze the data according to the group corresponding to the serial number. The commissioning organization and pathologist that was commissioned to perform the histopathological section experiment were unaware of the treatment group status of each sample during the histologic slide evaluation phase.
Briefly, the livers were first fixed in the neutral buffered formalin (10%) and for being handled for paraffin embedding according to the typical microtechnique. Four-micron thick sections of samples stained with haematoxylin and eosin dyes were used for the observation of histopathological damage under microscope (Olympus, Tokyo, Japan).
2.8. Statistical Analysis
The values were reported as the mean ± standard deviation (SD), and one-way analysis of variance (ANOVA) followed by Duncan’s multiple-range test were applied to perform the statistical differences between the groups. The p value < 0.05 was regarded as statistically significant.
3. Results
3.1. Effect of CGA in CCl4-Induced Hepatotoxicity
There was a remarkable increase (p < 0.05) of serum activity of AST, ALT, and total albumin in CCl4 treated group when contrasted with normal controls (Table 1), demonstrating that CCl4 induced hepatotoxicity. By contrast, mice treated with CGA at dosages of 60, 100, and 200 mg/kg were observed to have significant (p < 0.05) reductions in AST, ALT, and total albumin as compared to the CCl4-treated group. Silymarin at dosage of 200 mg/kg similarly diminished the activity of serum enzymes and increased the level of total albumin. These outcomes recommended the chance of CGA to give preservation against liver harm caused by CCl4 treatment.
Concerning serum lipid indicators, influences of CGA on cholesterol and TG were also examined (Table 1). CCl4 administration markedly amplified (p < 0.05) the cholesterol and TG when contrasted with normal control. However, CGA treatment significantly lessened (p < 0.05) cholesterol and TG in serum when contrasted with the CCl4-treated group. Similar outcomes were also observed in silymarin treated animals.
3.2. Hepatic Antioxidant Enzyme Activities
The antioxidant status of tissue could be used as an indicator by measuring the activity of antioxidant enzyme, such as catalase, SOD, GSH-Rd, and GSH-Px. The hepatic catalase, SOD, GSH-Rd, and GSH-Px activities in the CCl4 treated group were significantly lower than that in the normal control group (p < 0.05). Conversely, hepatic catalase, SOD, GSH-Rd, and GSH-Px activities in the CGA-treated groups at a dosage of 60, 100, and 200 mg/kg were significantly larger (p < 0.05) than that in the CCl4 treated group (Table 2). Silymarin at dosage of 200 mg/kg similarly diminished the activity of hepatic enzymes. The outcomes proposed the probability of CGA to give conservation against liver injury caused by CCl4 treatment.
3.3. Lipid Peroxidation and GSH
The content of TBARS is widely regarded as an indicator of free radical regulation of lipid peroxidation damage. In the CCl4 treated group, the TBARS were markedly greater than the control group (p < 0.05), demonstrating that CCl4 has stimulated severely injury to the liver. By contrast, CGA treated group (applied dose: 60, 100, and 200 mg/kg) showed remarkably lesser content of TBARS than that the CCl4 intoxicated group (p < 0.05). Silymarin also restrained the growing TBARS contents upon CCl4 treatment (Table 3).
In the antioxidant defense system, GSH is a very effective non-enzymatic antioxidant that diminishes the reactive metabolites toxicity of CCl4. CCl4 administration markedly diminished the hepatic GSH contents when contrasted with the normal control group (p < 0.05). By contrast, CGA treatment (applied dose: 60, 100, and 200 mg/kg) showed remarkably elevated content of GSH than that in CCl4 intoxicated group (p < 0.05). Similar outcomes were also observed in the silymarin group. These outcomes showed that the hepatic free radicals being induced were successfully eliminated when administered CGA.
3.4. Histopathologic Examination
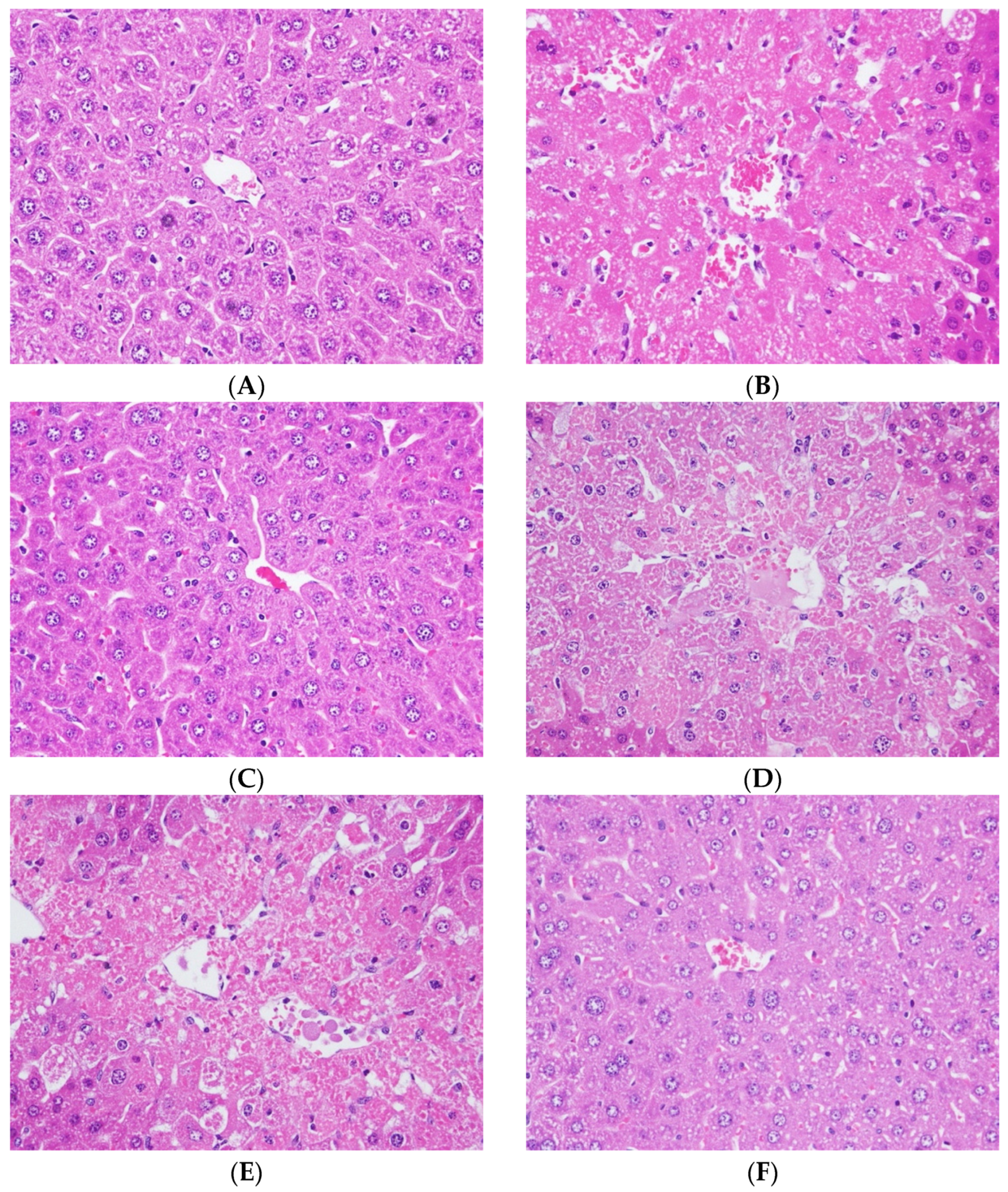
Histopathological examination is the most direct evidence for the degree of liver injury. In normal control mice, liver sections displayed typical hepatic cells with well-integrated cytoplasm, clearly nucleus (Figure 2A). In the CCl4-intoxicated group, liver sections visibly revealed moderated liver damage symptoms, such as moderate vacuolization, moderate diffused necrosis in hepatic lobules, as well as the mild inflammatory cell infiltration of hepatocytes (Figure 2B). The liver lesions of the silymarin-treated group were much lighter compared with the pathological changes found in the CCl4-intoxicated group (Figure 2C). These mice presented mild and moderate levels of hepatocellular necrosis, mild inflammatory infiltration of lymphocytes, and trace ballooning deterioration. Furthermore, a mild necrosis of hepatocytes, inflammatory cell infiltration, and ballooning deterioration were found in the liver of CGA-treated groups at 60, 100, and 200 mg/kg, respectively (Figure 2D–F).
Table 4 shows a semi-quantitative evaluation of histopathologic examinations including necrosis of hepatocytes, inflammatory cell infiltration, and ballooning deterioration. All damage grades and scores of histopathologic examinations in the CCl4-intoxicated group were markedly greater as compared to the normal control (p < 0.05), demonstrating that CCl4 could cause severe injury to the liver. On the contrary, all the treated doses of CGA markedly diminished (p < 0.05) the damage grades of hepatocyte necrosis, inflammatory cell infiltration, and ballooning deterioration when contrasted with CCl4-intoxicated group. Similar scores were also observed in the silymarin group. Additionally, there was no remarkable difference (p > 0.05) between each score of histopathologic examinations in silymarin group and CGA-treated group.
In the light of histopathologic examinations, serious hepatic injury resulted by CCl4 was amazingly diminished by the CGA treatment, which was in well correspondence with the outcomes of the serum biochemical analysis and liver antioxidant defense system.
4. Discussion
Carbon tetrachloride can be metabolized by Cytochrome P-450 into trichloromethyl radical (CCl3) and trichloromethyl peroxyl radical (CCl3O2), and further initiate peroxidation. Therefore, it is often used in experimental hepatopathy models [23]. Both radicals have the potential to combine with proteins or lipids and result in membrane lipid peroxidation, protein denaturation, and eventually cell necrosis [24]. Many reports have indicated that one of the major hypotheses of the liver protection efficacy may be related to the antioxidant ability to remove free radicals [25,26]. In fact, many studies have suggested that several natural products with antioxidant capacity, such as vitamin E [27], 5-O-Demethylnobiletin [4], and baicalein [28], diminish CCl4-induced hepatotoxic effects by removing ROS to prevent lipid peroxidation. In the current study, we discovered that CGA administration remarkably restrained CCl4-induced severe hepatic damage as proven by diminished AST, ALT, TG, and cholesterol, and increased total albumin in serum.
Silymarin is a flavonoid complex with good antioxidant capacity, which is isolated from the seeds of milk thistle (Silybum marianum, Compositae). It has been applied for clinical medicine to treat toxic liver diseases for more than 20 years. Silymarin has been proven to have excellent ROS scavenging ability and to increase GSH content in animals [29,30]. Furthermore, silymarin prevents CCl4-induced liver damage and oxidative damage through multiple mechanisms, including reducing the metabolic activation of CCl4, or blocking the reaction process of the free radical chain, acting as an antioxidant to remove ROS, or a combination of these effects [30]. In fact, many animal model studies and our previous studies had used silymarin as a positive control, which could effectively prevent lipid peroxidation and diminished the hepatotoxicity induced by CCl4 [29,30,31,32]. In this study, the positive control used silymarin, and the dose (200 mg/kg) used was based on our previous research [31,33]. Compared with the CCl4 treatment group, silymarin effectively reduced serum ALT, AST, total cholesterol, and TG levels, and significantly increased total albumin content.
Several reports have indicated that antioxidant enzymes in the body, including catalase, SOD, GSH-Rd, and GSH-Px, can protect tissues against oxidative damage [34,35]. SOD, an exceedingly efficacious oxidative enzyme, transformed the superoxide anions into hydrogen peroxide that would be metabolized to oxygen and water by catalase [36]. GSH-Px involves the hepatic detoxification of xenobiotics and transforms hydroperoxides and hydrogen peroxide to harmless metabolites. GSH-Rd is also involved in the hepatic detoxification of GSH conjugated with xenobiotic compounds [26,37]. These antioxidant enzymes would be lost their activity due to the elimination of ROS or inhibit lipid peroxidation. In addition, they are also effortlessly inactivated in response to the toxicity of CCl4 [38]. In the current study, hepatic antioxidant enzymes’ activities were remarkably diminished in CCl4 toxicity alone, suggesting increased hepatic oxidative injury in mice. Otherwise, CGA treatment to CCl4-intoxicated animals were remarkably increased hepatic catalase, SOD, GSH-Rd, and GSH-Px activities, implying that CGA has the capability to rehabilitate or maintain the activities of these antioxidant enzymes in CCl4-intoxicated liver.
Previous investigations on the mechanism of hepatotoxicity caused by CCl4 exhibited that GSH, an exceedingly efficacious antioxidant, could easily remove H2O2, ROS, and xenobiotic harmfulness [39]. Specifically, GSH consumption has a substantial correlation with the level of liver necrosis [40]. Consequently, increasing or maintaining the hepatic GSH conjugation is necessary to slow down CCl4-induced liver damage. GSH is effortlessly conjugated with xenobiotic compounds to be oxidized to GSSG, and also reacts with the selenium-containing GSH-Px isoenzymes, which eventually leads to a decrease in GSH levels. GSSG, the GSH in oxidation state, can react quickly with GSH-Rd and NADPH to reduce to GSH. Due to these recycling mechanisms in the antioxidant defense system, GSH is a very effective antioxidant against intracellular oxidative stress. [41]. In this study, significantly lower hepatic GSH level as a result of CCl4 was markedly elevated by CGA treatment, demonstrating that CGA can effectively prevent hepatic GSH consumption caused by CCl4.
Free radical derivatives formed by the metabolic transformation of CCl4 can cause lipid peroxidation in liver cells, which is one of the key hypotheses of CCl4-induced hepatotoxicity [42]. Various tissue damages and cell deaths are generally considered to be related to oxidative stress caused by lipid peroxidation, which is also an important underlying source of the progression of numerous chronic and acute diseases [43]. In the current study, CCl4-mediated toxicity resulted in a promotion of hepatic TBARS amount and CGA administration remarkably switched these results, demonstrating that CGA can successfully restrain lipid peroxidation resulted by CCl4.
Histopathological studies could provide a direct and important evidence for the biochemical analysis. Significant hepatotoxicity characterization can be inspected in the liver pathological tissue sections of the CCl4 group. Nevertheless, CGA treatment remarkably diminished these hepatotoxicity results in the liver pathological tissue sections (Figure 2); it directly proves that CGA has an excellent protective effect against liver injury caused by CCl4. The results of semi-quantitative analysis (Table 4) are also consistent with histological observations that CGA treatment remarkably diminished the liver toxicity of mice induced by CCl4. Additionally, there was no remarkable differentiation (p > 0.05) between each score of semi-quantitative assessment in CGA-treated group and silymarin group. In general, the outcomes of serum biochemical analysis and liver antioxidant enzyme examinations have shown that the hepatoprotective effect of CGA is equivalent to silymarin at all test doses and has been directly proven by liver histopathological examinations.
5. Conclusions
The outcomes of our study prove that CGA was successful in prophylaxis of CCl4-induced hepatotoxicity in mice through multiple mechanisms, including obstructing the process of lipid peroxidation, enhancing, or maintaining antioxidant enzymes activity, and increasing GSH level. Therefore, CGA in the diet may be beneficial as a hepatoprotective agent against chemical-induced liver toxicity in vivo.
Author Contributions
Y.-W.H. and C.-F.T. conceived and designed the research. Y.-W.H., Y.-Y.C. and C.-F.T. performed the experiments and analyzed the data. Y.-W.H. and C.-F.T. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wu, J.; Danielsson, A.; Zern, M.A. Toxicity of hepatotoxins: New insights into mechanisms and therapy. Expert Opin. Investig. Drugs 1999, 8, 585–607. [Google Scholar] [CrossRef]
- Vitaglione, P.; Morisco, F.; Caporaso, N.; Fogliano, V. Dietary antioxidant compounds and liver health. Crit. Rev. Food Sci. Nutr. 2004, 44, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofer, T. Oxidative stress in human toxicology. Antioxidants 2021, 10, 1159. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.N.; Kim, S.H.; Dey, D.K.; Park, S.M.; Nasif, O.; Bajpai, V.K.; Kang, S.C.; Lee, J.; Park, J.G. 5-O-Demethylnobiletin alleviates CCl4-induced acute liver injury by equilibrating ROS-mediated apoptosis and autophagy induction. Int. J. Mol. Sci. 2021, 22, 1083. [Google Scholar] [CrossRef]
- Geetha, S.; Jayamurthy, P.; Pal, K.; Pandey, S.; Kumar, R.; Sawhney, R.C. Hepatoprotective effects of sea buckthorn (Hippophae rhamnoides L.) against carbon tetrachloride induced liver injury in rats. J. Sci. Food Agric. 2008, 88, 1592–1597. [Google Scholar] [CrossRef]
- Boll, M.; Weber, L.W.; Becker, E.; Stampfl, A. Pathogenesis of carbon tetrachloride-induced hepatocyte injury bioactivation of CCl4 by cytochrome P450 and effects on lipid homeostasis. Z Naturforsch. C J. Biosci. 2001, 56, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhathal, P.S.; Rose, N.R.; Mackay, I.R.; Whittingham, S. Strain differences in mice in carbon tetrachloride-induced liver injury. Br. J. Exp. Pathol. 1983, 64, 524–533. [Google Scholar]
- Al-Amarat, W.; Abukhalil, M.H.; Althunibat, O.Y.; Alfwuaires, M.A.; Alnamshan, M.M.; Alqosaibi, A.I.; Ahmeda, A.F.; Kamel, E.M.; Arab, H.H.; Mahmoud, A.M. Galangin attenuates liver injury, oxidative stress and inflammation, and upregulates Nrf2/HO-1 signaling in streptozotocin-induced diabetic rats. Processes 2021, 9, 1562. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Hirata, Y.; Saito, T.; Kumagai, H. Combined effects of amino acids in garlic and buna-shimeji (Hypsizygus marmoreus) on suppression of CCl4-induced hepatic injury in rats. Foods 2021, 10, 1491. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.K.; Bansal, M.; Soni, G.; Bhatnagar, D. N-nitrosodiethylamine induced oxidative stress in rat liver. Chem. Biol. Interact. 2005, 156, 101–111. [Google Scholar] [CrossRef]
- Stocker, R. Dietary and pharmacological antioxidants in atherosclerosis. Curr. Opin. Lipidol. 1999, 10, 589–597. [Google Scholar] [CrossRef]
- Gonthier, M.P.; Verny, M.A.; Besson, C.; Rémésy, C.; Scalbert, A. Chlorogenic acid bioavailability largely depends on its metabolism by the gut microflora in rats. J. Nutr. 2003, 133, 1853–1859. [Google Scholar] [CrossRef] [Green Version]
- Clifford, M.N. Chlorogenic acids and other cinnamates—nature, occurrence and dietary burden. J. Sci. Food Agric. 1999, 79, 362–372. [Google Scholar] [CrossRef]
- Konishi, Y.; Kobayashi, S. Transepithelial transport of chlorogenic acid, caffeic acid, and their colonic metabolites in intestinal Caco-2 cell monolayers. J. Agric. Food Chem. 2004, 52, 2518–2526. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Lindsay, J.; Laurin, D.; Verreault, R.; Hebert, R.; Helliwell, B.; Hill, G.B.; McDowell, I. Risk factors for Alzheimer’s disease: A prospective analysis from the Canadian Study of Health and Aging. Am. J. Epidemiol. 2002, 156, 445–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar-Martinez, E.; Willett, W.C.; Ascherio, A.; Manson, J.E.; Leitzmann, M.F.; Stampfer, M.J.; Hu, F.B. Coffee consumption and risk for type 2 diabetes mellitus. Ann. Intern. Med. 2004, 140, 1–8. [Google Scholar] [CrossRef]
- Ranheim, T.; Halvorsen, B. Coffee consumption and human health: Beneficial or detrimental? Mechanisms for effects of coffee consumption on different risk factors for cardiovascular disease and type 2 diabetes mellitus. Mol. Nutr. Food Res. 2005, 49, 274–284. [Google Scholar] [CrossRef]
- Almeida, A.A.; Farah, A.; Silva, D.A.M.; Nunam, E.A.; Glória, M.B.A. Antibacterial activity of coffee extracts and selected coffee chemical compounds against enterobacteria. J. Agric. Food Chem. 2006, 54, 8738–8743. [Google Scholar] [CrossRef]
- Santos, M.D.; Almeida, M.C.; Lopes, N.P.; Souza, G.E.P. Evaluation of the antiinflamatory, analgesic and antypiretic activity of the natural polyphenol chlorogenic acid. Biol. Pharm. Bull. 2006, 29, 2236–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, J.; Farah, A.; de Paulis, T.; Bracy, D.P.; Pencek, R.R.; Graham, T.E.; Wasserman, D.H. Quinides of roasted coffee enhance insulin action in conscious rats. J. Nutr. 2003, 133, 3529–3532. [Google Scholar] [CrossRef]
- Berton, T.R.; Conti, C.J.; Mitchell, D.L.; Aldaz, C.M.; Lubet, R.A.; Fischer, S.M. The effect of vitamin E acetate on ultravioletinduced mouse skin carcinogenesis. Mol. Carcinog. 1998, 23, 175–184. [Google Scholar] [CrossRef]
- Goodla, L.; Manubolu, M.; Pathakoti, K.; Jayakumar, T.; Sheu, J.R.; Fraker, M.; Tchounwou, P.B.; Poondamalli, P.R. Protective effects of Ammannia baccifera against CCl4-induced oxidative stress in rats. Int. J. Environ. Res. Public Health 2019, 16, 1440. [Google Scholar] [CrossRef] [Green Version]
- Brattin, W.J.; Glende, E.A., Jr.; Recknagel, R.O. Pathological mechanisms in carbon tetrachloride hepatotoxicity. J. Free Radic. Biol. Med. 1985, 1, 27–38. [Google Scholar] [CrossRef]
- Hattori, T.; Ito, M.; Suzuki, Y. Studies on antinephritic effects of plant components in rats (1). Effects of saikosaponins originaltype anti-GBM nephritis in rats and its mechanisms. Nippon. Yakurigaku Zasshi 1991, 97, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.R.; Panda, V.S. Antioxidant and hepatoprotective effects of Ginkgo biloba phytosomes in carbon tetrachloride-induced liver injury in rodents. Liver Int. 2007, 27, 393–399. [Google Scholar] [CrossRef]
- Sodergren, E.; Cederberg, J.; Vessby, B.; Basu, S. Vitamin E reduces lipid peroxidation in experimental hepatotoxicity in rats. Eur. J. Nutr. 2001, 40, 10–16. [Google Scholar] [CrossRef]
- Dai, C.; Li, H.; Wang, Y.; Tang, S.; Velkov, T.; Shen, J. Inhibition of oxidative stress and ALOX12 and NF-κB pathways contribute to the protective effect of baicalein on carbon tetrachloride-induced acute liver injury. Antioxidants 2021, 10, 976. [Google Scholar] [CrossRef]
- Tsai, J.C.; Chiu, C.S.; Chen, Y.C.; Lee, M.S.; Hao, X.Y.; Hsieh, M.T.; Kao, C.P.; Peng, W.H. Hepatoprotective effect of Coreopsis tinctoria flowers against carbon tetrachloride-induced liver damage in mice. BMC Complement. Altern. Med. 2017, 17, 139. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Yang, Z.; Zhang, J.; Mu, J.; Zhou, X.; Zhao, X. Liver Injury Induced by Carbon Tetrachloride in Mice Is Prevented by the Antioxidant Capacity of Anji White Tea Polyphenols. Antioxidants 2019, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.F.; Hsu, Y.W.; Ting, H.C.; Huang, C.F.; Yen, C.C. The in vivo antioxidant and antifibrotic properties of green tea (Camellia sinensis, Theaceae). Food Chem. 2013, 136, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Demiroren, K.; Basunlu, M.T.; Erten, R.; Cokluk, E. A comparison of the effects of thymoquinone, silymarin and N-acetylcysteine in an experimental hepatotoxicity. Biomed. Pharmacother. 2018, 106, 1705–1712. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.W.; Tsai, C.F.; Chen, W.K.; Lu, F.J. Protective effects of seabuckthorn (Hippophae rhamnoides L.) seed oil against carbon tetrachloride- induced hepatotoxicity in mice. Food Chem. Toxicol. 2009, 47, 2281–2288. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Role of free radicals and catalytic metal irons in human disease: An overview. Meth. Enzymol. 1990, 186, 59–85. [Google Scholar]
- Wang, B.J.; Liu, C.T.; Tseng, C.Y.; Wu, C.P.; Yu, Z.R. Hepatoprotective and antioxidant effects of Bupleurum kaoi Liu (Chao et Chuang) extract and its fractions fractionated using supercritical CO2 on CCl4-induced liver damage. Food Chem. Toxicol. 2004, 42, 609–617. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.; Osuna, C.; Gitto, E. Actions of melatonin in the reduction of oxidative stress. J. Biomed. Sci. 2000, 7, 444–458. [Google Scholar] [CrossRef]
- Baudrimont, I.; Ahouandjivo, R.; Creppy, E.E. Prevention of lipid peroxidation induced by ochratoxin A in Vero cells in culture by several agents. Chem. Biol. Interact. 1997, 104, 29–40. [Google Scholar] [CrossRef]
- Yang, Y.S.; Ahn, T.H.; Lee, J.C.; Moon, C.J.; Kim, S.H.; Jun, W.; Park, S.C.; Kim, H.C.; Kim, J.C. Protective effects of Pycnogenol® on carbon tetrachloride-induced hepatotoxicity in Sprague–Dawley rats. Food Chem. Toxicol. 2008, 46, 380–387. [Google Scholar] [CrossRef]
- Kadiska, M.B.; Gladen, B.C.; Baird, D.D.; Dikalov, A.E.; Sohal, R.S.; Hatch, G.B.; Jones, D.P.; Mason, R.P.; Barret, J.C. Biomarkers of oxidative stress study: Are plasma antioxidants markers of CCl4 poisoning? J. Free Radic. Biol. Med. 2000, 28, 838–845. [Google Scholar] [CrossRef]
- Dambach, D.M.; Durham, S.K.; Laskin, J.D.; Laskin, D.L. Distinct roles of NFkappaB p50 in the regulation of acetaminophen-induced inflammatory mediator production and hepatotoxicity. Toxicol. Appl. Pharmacol. 2006, 211, 157–165. [Google Scholar] [CrossRef]
- Cantin, A.M.; White, T.B.; Cross, C.E.; Forman, H.J.; Sokol, R.J.; Borowitz, D. Antioxidants in cystic fibrosis conclusions from the CF antioxidant workshop, Bethesda, Maryland, November 11–12, 2003. J. Free Radic. Biol. Med. 2007, 42, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castor, J.A.; Ferrya, G.C.; Castro, C.R.; Sasame, S.; Fenes, O.M.; Gilette, J.R. Prevention of carbon tetrachloride-induced necrosis by inhibitors of drug metabolism. Further studies on the mechanism of their action. Biochem. Pharmacol. 1974, 23, 295–302. [Google Scholar] [CrossRef]
- Halliwell, B. Antioxidants and human disease: A general introduction. Nutr. Rev. 1997, 55, 44–52. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of chlorogenic acid.

Figure 2.
The morphological analysis of hepatoprotective effect of the CGA in CCl4-intoxicated mice. Dissected liver tissues were stained with haematoxylin–eosin by standard techniques (400×). (A) Normal control; (B) CCl4 control; (C) silymarin (200 mg/kg) + CCl4; (D) CGA (60 mg/kg) + CCl4; (E) CGA (100 mg/kg) + CCl4; and (F) CGA (200 mg/kg) + CCl4.
Figure 2.
The morphological analysis of hepatoprotective effect of the CGA in CCl4-intoxicated mice. Dissected liver tissues were stained with haematoxylin–eosin by standard techniques (400×). (A) Normal control; (B) CCl4 control; (C) silymarin (200 mg/kg) + CCl4; (D) CGA (60 mg/kg) + CCl4; (E) CGA (100 mg/kg) + CCl4; and (F) CGA (200 mg/kg) + CCl4.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of CGA on serum AST, ALT, cholesterol, and TG in CCl4 intoxicated mice.
| Design of Treatment | AST (Units/L) | ALT (Units/L) | Cholesterol (mg/dL) | TG (mg/dL) | Total Albumin (mg/dL) |
|---|---|---|---|---|---|
| Normal control | 15.9 ± 3.45 e | 33.1 ± 6.22 e | 223 ± 14.4 b | 162 ± 12.3 c | 38.7 ± 3.52 b |
| CCl4 (3 mL/kg i.p.) | 1230 ± 104 a | 1253 ± 115 a | 313 ± 22.3 a | 261 ± 15.5 a | 22.5 ± 0.77 a |
| Silymarin + CCl4 | 7.37 ± 2.01 f | 16.5 ± 2.60 f | 238 ± 12.9 b | 181 ± 16.0 b | 38.9 ± 1.81 b |
| CGA (60 mg/kg) + CCl4 | 895 ± 29.7 b | 827 ± 71.2 b | 265 ± 20.2 b | 209 ± 11.7 b | 37.2 ± 0.82 b |
| CGA (100 mg/kg) + CCl4 | 390 ± 33.6 c | 317 ± 10.4 c | 249 ± 13.8 b | 188 ± 12.5 b | 38.5 ± 0.67 b |
| CGA (200 mg/kg) + CCl4 | 42.6 ± 4.99 d | 58.5 ± 6.50 d | 197 ± 15.5 b | 155 ± 9.81 c | 41.4 ± 1.12 b |
| p-Value | 0.000 | 0.000 | 0.010 | 0.016 | 0.010 |
Data are mean ± SD, n = 10. a–f Mean values with different letters in the same row are significantly different (p < 0.05) according to Duncan’s multiple-range test.
Table 2.
Effects of CGA on hepatic catalase, SOD, GSH-Rd, and GSH-Px in CCl4 intoxicated mice.
| Design of Treatment | Catalase (Units/mg Protein) | SOD (Units/mg Protein) | GSH-Rd (nmole NADPH/min/mg Protein) | GSH-Px (nmole NADPH/min/mg Protein) |
|---|---|---|---|---|
| Normal control | 18.9 ± 1.63 b | 22.1 ± 2.28 d | 99.5 ± 1.22 c | 493 ± 53.4 d |
| CCl4 (3 mL/kg i.p.) | 5.62 ± 0.98 a | 10.9 ± 1.24 a | 72.3 ± 5.09 a | 277 ± 23.2 a |
| Silymarin + CCl4 | 21.0 ± 3.04 b | 15.4 ± 1.11 c | 85.2 ± 2.24 b | 353 ± 30.1 b,c |
| CGA (60 mg/kg) + CCl4 | 15.3 ± 2.58 b | 12.4 ± 1.32 b | 77.5 ± 1.16 a | 309 ± 15.2 b |
| CGA (100 mg/kg) + CCl4 | 19.9 ± 1.18 b | 15.2 ± 1.72 c | 95.9 ± 0.85 c | 453 ± 25.0 d |
| CGA (200 mg/kg) + CCl4 | 21.9 ± 3.37 b | 20.3 ± 1.53 d | 98.1 ± 7.00 c | 544 ± 31.7 e |
| p-Value | 0.008 | 0.020 | 0.041 | 0.011 |
Data are mean ± SD, n = 10. a–e Mean values with different letters in the same row are markedly different (p < 0.05) according to Duncan’s multiple-range test.
Table 3.
Effects of CGA on hepatic TBARS and GSH in CCl4 intoxicated mice.
| Design of Treatment | TBARS (nmol/mg Protein) | GSH (μmol/g wet Weight) |
|---|---|---|
| Normal control | 1.28 ± 0.18 d | 4.51 ± 0.58 c |
| CCl4 (3 mL/kg i.p.) | 3.08 ± 0.41 a | 2.60 ± 0.44 a |
| Silymarin + CCl4 | 1.37 ± 0.31 d | 4.47 ± 0.21 c |
| CGA (60 mg/kg) + CCl4 | 2.12 ± 0.14 b | 3.67 ± 0.52 b |
| CGA (100 mg/kg) + CCl4 | 1.75 ± 0.15 c | 4.01 ± 0.22 b,c |
| CGA (200 mg/kg) + CCl4 | 1.29 ± 0.09 d | 4.65 ± 0.19 c |
| p-Value | 0.032 | 0.010 |
Data are mean ± SD, n = 10. a–d Mean values with different letters in the same row are markedly different (p < 0.05) according to Duncan’s multiple-range test.
Table 4.
Effects of CGA on hepatic histopathology of liver injury in mice treated with CCl4.
| Parameter | Grades */Score ** | Design of Treatment | |||||
|---|---|---|---|---|---|---|---|
| Normal Control | CCl4 Control (3 mL/kg) | Silymarin (200 mg/kg) + CCl4 | CGA (60 mg/kg) + CCl4 | CGA (100 mg/kg) + CCl4 | CGA (200 mg/kg) + CCl4 | ||
| Hepatocyte necrosis | − | 6 | 0 | 0 | 0 | 0 | 0 |
| + | 4 | 0 | 3 | 1 | 4 | 3 | |
| ++ | 0 | 2 | 7 | 4 | 6 | 7 | |
| +++ | 0 | 8 | 0 | 5 | 0 | 0 | |
| Score | 0.40 ± 0.52 b,c | 2.80 ± 0.42 a,c | 1.70 ± 0.48 a,b | 2.4 ± 0.70 a | 1.60 ± 0.52 a,b | 1.70 ± 0.48 a,b | |
| Inflammatory cell infiltration | − | 7 | 0 | 0 | 0 | 0 | 0 |
| + | 3 | 0 | 0 | 0 | 0 | 0 | |
| ++ | 0 | 3 | 9 | 7 | 9 | 10 | |
| +++ | 0 | 7 | 1 | 3 | 1 | 0 | |
| Score | 0.30 ± 0.48 b,c | 2.70 ± 0.48 a,c | 2.10 ± 0.32 a,b | 2.30 ± 0.48 a | 2.10 ± 0.32 a,b | 2.00 ± 0.00 a,b | |
| Ballooning deterioration | − | 9 | 0 | 0 | 0 | 0 | 0 |
| + | 1 | 0 | 5 | 0 | 0 | 2 | |
| ++ | 0 | 2 | 4 | 7 | 8 | 7 | |
| +++ | 0 | 8 | 1 | 3 | 2 | 1 | |
| Score | 0.10 ± 0.32 b,c | 2.80 ± 0.42 a,c | 1.60 ± 0.70 a,b | 2.30 ± 0.48 a | 2.20 ± 0.42 a | 1.90 ± 0.57 a,b | |
* Grades are as follows: − = absent, + = trace (1–25%), ++ = weak (26–50%), and +++ = moderate (50–75%). ** The numerical score for histopathologic examinations were the result of adding the number per grade of affected animals and dividing by the total number of tested animals. a Remarkably different from normal control, p < 0.05. b Remarkably different from CCl4 control, p < 0.05. c Remarkably different from silymarin group, p < 0.05.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hsu, Y.-W.; Chen, Y.-Y.; Tsai, C.-F. Protective Effects of Chlorogenic Acid against Carbon Tetrachloride-Induced Hepatotoxicity in Mice. Processes 2022, 10, 31. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10010031
AMA Style
Hsu Y-W, Chen Y-Y, Tsai C-F. Protective Effects of Chlorogenic Acid against Carbon Tetrachloride-Induced Hepatotoxicity in Mice. Processes. 2022; 10(1):31. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10010031
Chicago/Turabian StyleHsu, Yu-Wen, Ya-Yu Chen, and Chia-Fang Tsai. 2022. "Protective Effects of Chlorogenic Acid against Carbon Tetrachloride-Induced Hepatotoxicity in Mice" Processes 10, no. 1: 31. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10010031
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.