
Modulation of Volatile Thiol Release during Fermentation of Red Musts by Wine Yeast

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Microorganisms and Culture Conditions
2.3. Laboratory-Scale Fermentation of Pinot Noir
2.4. Small Scale Winemaking of Grenache
2.5. Assay of Cysteine-S-Conjugate β-Lyase Activity in Yeast Cell-Free Protein Extracts
2.6. Analysis of Principal Non-Volatile Compounds by HPLC
2.7. Analysis of Aroma Compounds
2.8. Sensory Evaluation
2.9. Statistical Analysis
3. Results
3.1. Enzymatic Assay for Cysteine-S-Conjugate β-Lyase Activity in Yeast Cell-Free Protein Extracts
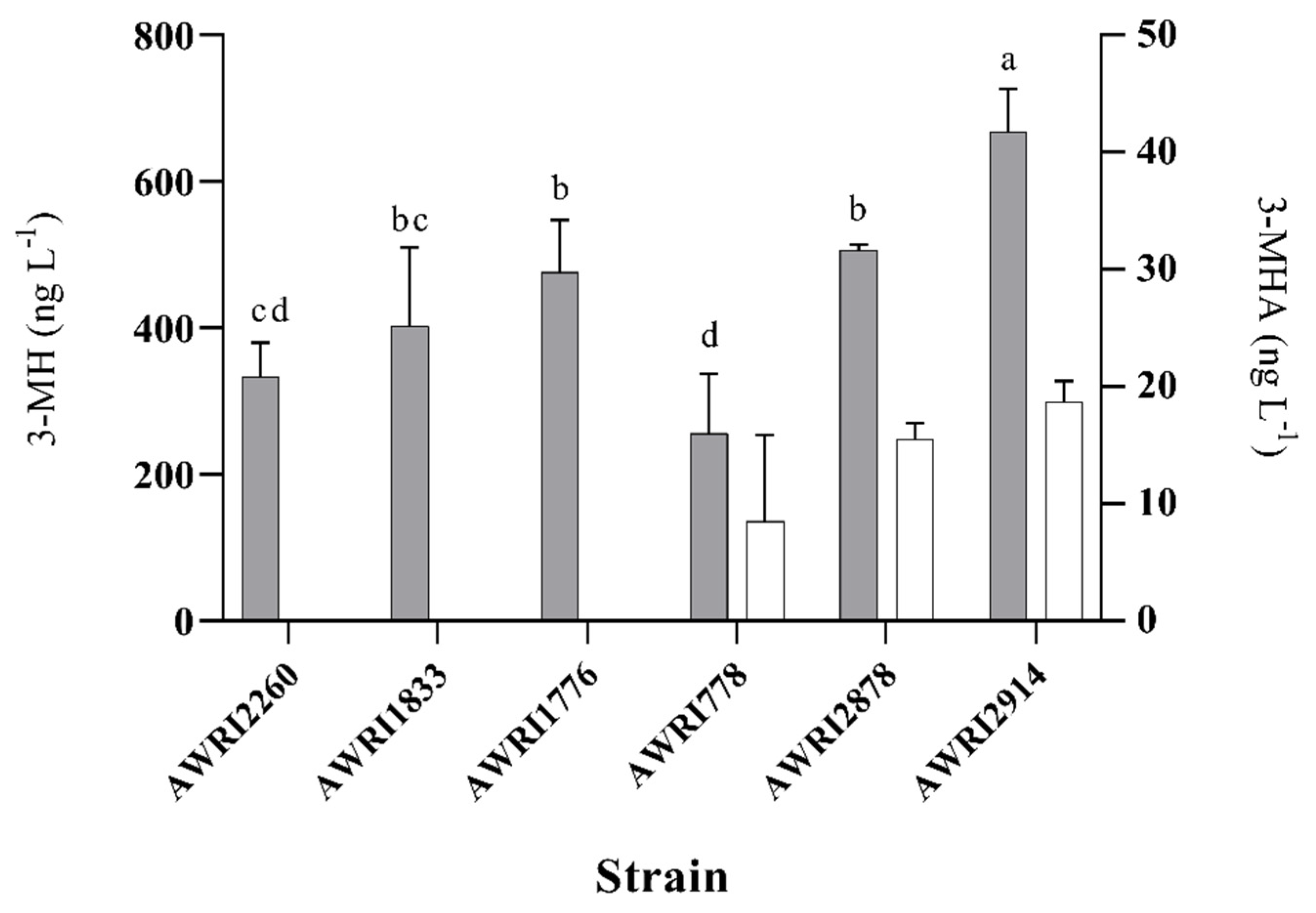
3.2. Laboratory-Scale Fermentations in Pinot Noir Must
4. Pilot-Scale Winemaking with Grenache
5. Quantitative Sensory Descriptive Analysis of the Grenache Wines
6. Linking Chemical Composition to Sensory Properties
7. Discussion
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Darriet, P.; Tominaga, T.; Lavigne, V.; Boidron, J.-N.; Dubourdieu, D. Identification of a powerful aromatic component of Vitis vinifera L. var. sauvignon wines: 4-mercapto-4-methylpentan-2-one. Flavour Fragr. J. 1995, 10, 385–392. [Google Scholar] [CrossRef]
- Tominaga, T.; Furrer, A.; Henry, R.; Dubourdieu, D. Identification of new volatile thiols in the aroma of Vitis vinifera L. var. Sauvignon blanc wines. Flavour Fragr. J. 1998, 13, 159–162. [Google Scholar] [CrossRef]
- Tominaga, T.; Darriet, P.; Dubourdieu, D. Identification de l’acétate de 3-mercaptohexanol, composé à forte odeur de buis, intervenant dans l’arôme des vins de Sauvignon. Vitis 1996, 35, 207–210. [Google Scholar]
- Capone, D.L.; Barker, A.; Williamson, P.O.; Francis, I.L. The role of potent thiols in Chardonnay wine aroma. Aust. J. Grape Wine Res. 2017, 24, 38–50. [Google Scholar] [CrossRef]
- Guth, H. Quantitation and sensory studies of character impact odorants of different white wine varieties. J. Agric. Food Chem. 1997, 45, 3027–3032. [Google Scholar] [CrossRef]
- Capone, D.L.; Ristic, R.; Pardon, K.H.; Jeffery, D.W. Simple quantitative determination of potent thiols at ultratrace levels in wine by derivatization and high-performance liquid chromatography-tandem mass spectrometry (HPLC-MS/MS) analysis. Anal. Chem. 2015, 87, 1226–1231. [Google Scholar] [CrossRef] [PubMed]
- Rigou, P.; Triay, A.; Razungles, A. Influence of volatile thiols in the development of blackcurrant aroma in red wine. Food Chem. 2014, 142, 242–248. [Google Scholar] [CrossRef]
- Culleré, L.; Escudero, A.; Cacho, J.; Ferreira, V. Gas chromatography-olfactometry and chemical quantitative study of the aroma of six premium quality spanish aged red wines. J. Agric. Food Chem. 2004, 52, 1653–1660. [Google Scholar] [CrossRef] [PubMed]
- Siebert, T.E.; Francis, I.L.; Pisaniello, L.; Melzer, S.; Bey, L.; Watson, F.; Cordente, T. Do varietal thiols matter in red wine? AWRI Technol. Rev. 2019, 183, 10–15. [Google Scholar]
- Fang, Y.; Qian, M. Aroma compounds in Oregon Pinot Noir wine determined by aroma extract dilution analysis (AEDA). Flavour Fragr. J. 2005, 20, 22–29. [Google Scholar] [CrossRef]
- Smith, M.E.; Bekker, M.Z.; Smith, P.A.; Wilkes, E.N. Sources of volatile sulfur compounds in wine. Aust. J. Grape Wine Res. 2015, 21, 705–712. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.; Swiegers, J.H.; Elsey, G.M.; Siebert, T.E.; Bartowsky, E.J.; Fleet, G.H.; Pretorius, I.S.; de Barros Lopes, M.A. Variation in 4-mercapto-4-methyl-pentan-2-one release by Saccharomyces cerevisiae commercial wine strains. FEMS Microbiol. Lett. 2004, 240, 125–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, E.S.; Swiegers, J.H.; Travis, B.; Francis, I.L.; Bastian, S.E.; Pretorius, I.S. Coinoculated Fermentations Using Saccharomyces Yeasts Affect the Volatile Composition and Sensory Properties of Vitis vinifera L. cv. Sauvignon Blanc Wines. J. Agric. Food Chem. 2008, 56, 10829–10837. [Google Scholar] [CrossRef] [PubMed]
- Murat, M.-L.; Masneuf, I.; Darriet, P.; Lavigne, V.; Tominaga, T.; Dubourdieu, D. Effect of Saccharomyces cerevisiae Yeast Strains on the Liberation of Volatile Thiols in Sauvignon blanc Wine. Am. J. Enol. Vitic. 2001, 52, 136. [Google Scholar]
- Cordente, A.G.; Borneman, A.R.; Bartel, C.; Capone, D.; Solomon, M.; Roach, M.; Curtin, C.D. Inactivating Mutations in Irc7p Are Common in Wine Yeasts, Attenuating Carbon-Sulfur β-Lyase Activity and Volatile Sulfur Compound Production. Appl. Environ. Microbiol. 2019, 85, e02684-18. [Google Scholar] [CrossRef] [Green Version]
- Tofalo, R.; Perpetuini, G.; Battistelli, N.; Tittarelli, F.; Suzzi, G. Correlation between IRC7 gene expression and 4-mercapto-4-methylpentan-2-one production in Saccharomyces cerevisiae strains. Yeast 2020, 37, 487–495. [Google Scholar] [CrossRef]
- Capone, D.L.; Sefton, M.A.; Hayasaka, Y.; Jeffery, D.W. Analysis of precursors to wine odorant 3-mercaptohexan-1-ol using HPLC-MS/MS: Resolution and quantitation of diastereomers of 3-S-cysteinylhexan-1-ol and 3-S-glutathionylhexan-1-ol. J. Agric. Food Chem. 2010, 58, 1390–1395. [Google Scholar] [CrossRef]
- Fedrizzi, B.; Pardon, K.H.; Sefton, M.A.; Elsey, G.M.; Jeffery, D.W. First identification of 4-S-glutathionyl-4-methylpentan-2-one, a potential precursor of 4-mercapto-4-methylpentan-2-one, in Sauvignon Blanc juice. J. Agric. Food Chem. 2009, 57, 991–995. [Google Scholar] [CrossRef]
- Capone, D.L.; Pardon, K.H.; Cordente, A.G.; Jeffery, D.W. Identification and quantitation of 3-S-cysteinylglycinehexan-1-ol (Cysgly-3-MH) in Sauvignon blanc grape juice by HPLC-MS/MS. J. Agric. Food Chem. 2011, 59, 11204–11210. [Google Scholar] [CrossRef]
- Cordente, A.G.; Capone, D.L.; Curtin, C.D. Unravelling glutathione conjugate catabolism in Saccharomyces cerevisiae: The role of glutathione/dipeptide transporters and vacuolar function in the release of volatile sulfur compounds 3-mercaptohexan-1-ol and 4-mercapto-4-methylpentan-2-one. Appl. Microbiol. Biotechnol. 2015, 99, 9709–9722. [Google Scholar] [CrossRef] [PubMed]
- Bonnaffoux, H.; Roland, A.; Remond, E.; Delpech, S.; Schneider, R.; Cavelier, F. First identification and quantification of S-3-(hexan-1-ol)-gamma-glutamyl-cysteine in grape must as a potential thiol precursor, using UPLC-MS/MS analysis and stable isotope dilution assay. Food Chem. 2017, 237, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.H.; Willmott, R.; Hill-Ling, A.; Capone, D.L.; Pardon, K.H.; Elsey, G.M.; Howell, K.S.; de Barros Lopes, M.A.; Sefton, M.A.; Lilly, M.; et al. Modulation of volatile thiol and ester aromas by modified wine yeast. In Developments in Food Science; Bredie, W.L.P., Petersen, M.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 43, pp. 113–116. [Google Scholar]
- Lilly, M.; Lambrechts, M.G.; Pretorius, I.S. Effect of increased yeast alcohol acetyltransferase activity on flavor profiles of wine and distillates. Appl. Environ. Microbiol. 2000, 66, 744–753. [Google Scholar] [CrossRef] [Green Version]
- Roncoroni, M.; Santiago, M.; Hooks, D.O.; Moroney, S.; Harsch, M.J.; Lee, S.A.; Richards, K.D.; Nicolau, L.; Gardner, R.C. The yeast IRC7 gene encodes a beta-lyase responsible for production of the varietal thiol 4-mercapto-4-methylpentan-2-one in wine. Food Microbiol. 2011, 28, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Cordente, A.G.; Williams, S.J.; Capone, D.L.; Jitjaroen, W.; Menz, I.R.; Curtin, C.; Anderson, P.A. Engineering Saccharomyces cerevisiae to release 3-mercaptohexan-1-ol during fermentation through overexpression of an S. cerevisiae gene, STR3, for improvement of wine aroma. Appl. Environ. Microbiol. 2011, 77, 3626–3632. [Google Scholar] [CrossRef] [Green Version]
- Zha, M.; Yin, S.; Sun, B.; Wang, X.; Wang, C. STR3 and CYS3 Contribute to 2-Furfurylthiol Biosynthesis in Chinese Sesame-Flavored Baijiu Yeast. J. Agric. Food Chem. 2017, 65, 5503–5511. [Google Scholar] [CrossRef]
- Howell, K.; Klein, M.; Swiegers, J.H.; Hayasaka, Y.; Elsey, G.M.; Fleet, G.H.; de Barros Lopes, M.A.; Høj, P.B.; Pretorius, I.S. Genetic determinants of volatile-thiol release by Saccharomyces cerevisiae during wine fermentation. Appl. Environ. Microbiol. 2005, 71, 5420–5426. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Ruiz, J.; Navascues, E.; Marquina, D.; Santos, A. Improvement of aromatic thiol release through the selection of yeasts with increased beta-lyase activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef]
- Ruiz, J.; de Celis, M.; Martín-Santamaría, M.; Benito-Vázquez, I.; Pontes, A.; Lanza, V.F.; Sampaio, J.P.; Santos, A.; Belda, I. Global distribution of IRC7 alleles in Saccharomyces cerevisiae populations: A genomic and phenotypic survey within the wine clade. Environ. Microbiol. 2021, 23, 3182–3195. [Google Scholar] [CrossRef]
- Dufour, M.; Zimmer, A.; Thibon, C.; Marullo, P. Enhancement of volatile thiol release of Saccharomyces cerevisiae strains using molecular breeding. Appl. Microbiol. Biotechnol. 2013, 97, 5893–5905. [Google Scholar] [CrossRef]
- Kotseridis, Y.; Ray, J.-L.; Augier, C.; Baumes, R. Quantitative determination of sulfur containing wine odorants at sub-ppb levels. 1. Synthesis of the deuterated analogues. J. Agric. Food Chem. 2000, 48, 5819–5823. [Google Scholar] [CrossRef] [PubMed]
- Pardon, K.H.; Graney, S.D.; Capone, D.L.; Swiegers, J.H.; Sefton, M.A.; Elsey, G.M. Synthesis of the individual diastereomers of the cysteine conjugate of 3-mercaptohexanol (3-MH). J. Agric. Food Chem. 2008, 56, 3758–3763. [Google Scholar] [CrossRef] [PubMed]
- Grant-Preece, P.A.; Pardon, K.H.; Capone, D.L.; Cordente, A.G.; Sefton, M.A.; Jeffery, D.W.; Elsey, G.M. Synthesis of wine thiol conjugates and labeled analogues: Fermentation of the glutathione conjugate of 3-mercaptohexan-1-ol yields the corresponding cysteine conjugate and free thiol. J. Agric. Food Chem. 2010, 58, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Cordente, A.G.; Heinrich, A.; Pretorius, I.S.; Swiegers, J.H. Isolation of sulfite reductase variants of a commercial wine yeast with significantly reduced hydrogen sulfide production. FEMS Yeast Res. 2009, 9, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Siebert, T.E.; Solomon, M.R.; Pollnitz, A.P.; Jeffery, D.W. Selective determination of volatile sulfur compounds in wine by gas chromatography with sulfur chemiluminescence detection. J. Agric. Food Chem. 2010, 58, 9454–9462. [Google Scholar] [CrossRef]
- Winter, G.; Henschke, P.A.; Higgins, V.J.; Ugliano, M.; Curtin, C.D. Effects of rehydration nutrients on H2S metabolism and formation of volatile sulfur compounds by the wine yeast VL3. AMB Express 2011, 1, 36. [Google Scholar] [CrossRef] [Green Version]
- Cordente, A.G.; Espinase Nandorfy, D.; Solomon, M.; Schulkin, A.; Kolouchova, R.; Francis, I.L.; Schmidt, S.A. Aromatic Higher Alcohols in Wine: Implication on Aroma and Palate Attributes during Chardonnay Aging. Molecules 2021, 26, 4979. [Google Scholar] [CrossRef]
- Heymann, H.; King, E.; Hopfer, H. Classical Descriptive Analysis. In Novel Techniques in Sensory Characterization and Consumer Profiling, 1st ed.; Varela, P., Ares, G., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 9–40. [Google Scholar]
- Day, M.; Espinase Nandorfy, D.; Bekker, M.; Bindon, K.; Solomon, M.; Smith, P.; Schmidt, S. Aeration of Vitis vinifera Shiraz fermentation and its effect on wine chemical composition and sensory attributes. Aust. J. Grape Wine Res. 2021, 27, 360–377. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Capone, D.L.; Pardon, K.H.; Elsey, G.M.; Sefton, M.A.; Francis, I.L.; Pretorius, I.S. Engineering volatile thiol release in Saccharomyces cerevisiae for improved wine aroma. Yeast 2007, 24, 561–574. [Google Scholar] [CrossRef]
- Santiago, M.; Gardner, R.C. TheIRC7gene encodes cysteine desulphydrase activity and confers on yeast the ability to grow on cysteine as a nitrogen source. Yeast 2015, 32, 519–532. [Google Scholar] [CrossRef]
- Santiago, M.; Gardner, R.C. Yeast genes required for conversion of grape precursors to varietal thiols in wine. FEMS Yeast Res. 2015, 15, fov034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, L.D.; Vannevel, S.; Buica, A.; Callerot, S.; Fedrizzi, B.; Kilmartin, P.; du Toit, W.J. Indications of the prominent role of elemental sulfur in the formation of the varietal thiol 3-mercaptohexanol in Sauvignon blanc wine. Food Res. Int. 2017, 98, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Harsch, M.J.; Benkwitz, F.; Frost, A.; Colonna-Ceccaldi, B.; Gardner, R.C.; Salmon, J.-M. New precursor of 3-mercaptohexan-1-ol in grape juice: Thiol-forming potential and kinetics during early stages of must fermentation. J. Agric. Food Chem. 2013, 61, 3703–3713. [Google Scholar] [CrossRef] [PubMed]
- Murat, M.-L.; Tominaga, T.; Dubourdieu, D. Assessing the Aromatic Potential of Cabernet Sauvignon and Merlot Musts Used to Produce Rose Wine by Assaying the Cysteinylated Precursor of 3-Mercaptohexan-1-ol. J. Agric. Food Chem. 2001, 49, 5412–5417. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, L.; Capone, D.L.; Roland, A.; Jeffery, D.W. Evolution and Correlation of cis-2-Methyl-4-propyl-1,3-oxathiane, Varietal Thiols, and Acetaldehyde during Fermentation of Sauvignon blanc Juice. J. Agric. Food Chem. 2020, 68, 8676–8687. [Google Scholar] [CrossRef] [PubMed]
- Capone, D.L.; Sefton, M.A.; Jeffery, D.W. Application of a modified method for 3-mercaptohexan-1-ol determination to investigate the relationship between free thiol and related conjugates in grape juice and wine. J. Agric. Food Chem. 2011, 59, 4649–4658. [Google Scholar] [CrossRef]
- Pinu, F.R.; Jouanneau, S.; Nicolau, L.; Gardner, R.C.; Villas-Boas, S.G. Concentrations of the Volatile Thiol 3-Mercaptohexanol in Sauvignon blanc Wines: No Correlation with Juice Precursors. Am. J. Enol. Vitic. 2012, 63, 407–412. [Google Scholar] [CrossRef]
- Peyrot des Gachons, C.; Tominaga, T.; Dubourdieu, D. Localization of S-cysteine conjugates in the berry: Effect of skin contact on aromatic potential of Vitis vinifera L. cv. Sauvignon Blanc must. Am. J. Enol. Vitic. 2002, 53, 144. [Google Scholar]
- Roland, A.; Schneider, R.; Charrier, F.; Cavelier, F.; Rossignol, M.; Razungles, A. Distribution of varietal thiol precursors in the skin and the pulp of Melon B. and Sauvignon Blanc grapes. Food Chem. 2011, 125, 139–144. [Google Scholar] [CrossRef]
- Maggu, M.; Winz, R.; Kilmartin, P.A.; Trought, M.C.T.; Nicolau, L. Effect of Skin Contact and Pressure on the Composition of Sauvignon Blanc Must. J. Agric. Food Chem. 2007, 55, 10281–10288. [Google Scholar] [CrossRef]
- Ferreira, V.; Ortín, N.; Escudero, A.; López, R.; Cacho, J. Chemical Characterization of the Aroma of Grenache Rosé Wines: Aroma Extract Dilution Analysis, Quantitative Determination, and Sensory Reconstitution Studies. J. Agric. Food Chem. 2002, 50, 4048–4054. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V. Volatile aroma compounds and wine sensory attributes. In Managing Wine Quality; Reynolds, A.G., Ed.; Woodhead Publishing: New York, NY, USA, 2010; pp. 3–28. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main IRC7 Genotype a | Additional SNPs b | Strains (n) | β-Lyase Activity (nmol min−1 mg prot −1) c | Strains Used for Fermentations d |
|---|---|---|---|---|
| IRC7S/IRC7S | - | 15 | 1.99 ± 0.31 d | AWRI1833, AWRI2865 |
| IRC7S/IRC7L | - | 5 | 6.42 ± 1.3 b | AWRI2878 |
| IRC7L T185A | 5 | 2.96 ± 1.28 cd | AWRI2260 | |
| IRC7L T185A P146R | 3 | 2.6 ± 0.8 cd | AWRI1776 | |
| IRC7L/IRC7L | - | 5 | 10.6 ± 2.35 a | AWRI2914, AWRI1082 |
| IRC7L/IRC7L T185A | 1 | 7.88 ab | AWRI2861 | |
| IRC7L T185A/IRC7L T185A | 2 | 5.62 ± 0.53 bc | AWRI1688 | |
| IRC7L T185A K43R/IRC7L T185A K43R | 2 | 3.94 ± 1.06 bcd | AWRI1493 | |
| IRC7L T185A/IRC7L T185A G321D | 1 | 3.33 bcd | AWRI778 |
| Precursor Concentration (µg L−1) | % Precursor Consumption | % Maximum Molar Conversion a | |||
|---|---|---|---|---|---|
| (R/S)-GSH-3-MH | (R/S)-Cys-Gly-3-MH | (R/S)-Cys-3-MH | |||
| Grape must | 1176 | 24.9 | 129.7 | ||
| Wines | |||||
| AWRI1082 | 13.2 ± 0.8 | 28.1 ± 2.7 | 0.6 ± 0.1 | 96.2 | 0.66 |
| AWRI1493 | 627 ± 26.2 | 32.1 ± 4.2 | 58.9 ± 0.9 | 46.1 | 0.10 |
| AWRI1688 | 33.9 ± 4.6 | 5.2 ± 0.6 | 1.3 ± 0.2 | 97.0 | 0.30 |
| AWRI1776 | 220 ± 9.0 | 19.7 ± 2.1 | 1.1 ± 0.2 | 82.7 | 0.14 |
| AWRI1833 | 288 ± 1.1 | 12.8 ± 0.7 | 1.3 ± 0.1 | 78.7 | 0.17 |
| AWRI2260 | nd | 3.7 ± 0.2 | 0.9 ± 0.1 | 99.5 | 0.13 |
| AWRI2861 | 18.5 ± 0.1 | 7.7 ± 0.9 | 2.5 ± 0.3 | 97.6 | 0.36 |
| AWRI2865 | 1085 ± 23 | 59.7 ± 1.6 | 120.1 ± 8 | 4.0 | 0.03 |
| AWRI2878 | nd | nd | 3.7 ± 0.4 | 99.5 | 0.22 |
| AWRI2914 | 9.4 ± 0.8 | 4.4 ± 0.6 | 58.0 ± 5.8 | 91.5 | 0.62 |
| AWRI778 | 531 ± 71 | 12.1 ± 1.5 | 127.3 ± 13 | 46.0 | 0.14 |
| Strain | VSC (µg L−1) | H2S Released (µg) a | |||
|---|---|---|---|---|---|
| H2S | MeSH | EtSH | MeSAc | ||
| AWRI1082 | 2.2 ± 0.2 | 0 | 0 | 5.5 ± 0.8 | 112 ± 12 |
| AWRI1493 | 1.3 ± 0.1 | 0.9 ± 0.1 | 0 | 7.9 ± 2.3 | 66 ± 3.5 |
| AWRI1688 | 1.7 ± 0.2 | 1.0 ± 0.1 | 0 | 9.2 ± 2.2 | 124 ± 2.1 |
| AWRI1776 | 2.6 ± 0.4 | 0 | 0 | 17.4 ± 2.6 | 60 ± 1.4 |
| AWRI1833 | 4.0 ± 0.4 | 0.8 ± 0.1 | 1.2 ± 0.1 | 6.0 ± 1.1 | 67 ± 2.9 |
| AWRI2260 | 2.4 ± 0.1 | 0.9 ± 0.1 | 0 | 17.4 ± 2.4 | 55 ± 0.8 |
| AWRI2861 | 2.0 ± 0.3 | 1.1 ± 0.2 | 0 | 19.9 ± 16.8 | 57 ± 6.6 |
| AWRI2865 | 1.9 ± 0.5 | 0 | 0 | 0 | 6.7 ± 0.4 |
| AWRI2878 | 0.7 ± 0.6 | 0 | 0 | 0 | 0 |
| AWRI2914 | 2.3 ± 0.5 | 1.0 ± 0.3 | 0 | 17.9 ± 4.5 | 163 ± 1.6 |
| AWRI778 | 0.9 ± 0.8 | 0 | 0 | 7.8 ± 0.9 | 113 ± 3.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordente, A.G.; Curtin, C.D.; Solomon, M.; Kulcsar, A.C.; Watson, F.; Pisaniello, L.; Schmidt, S.A.; Espinase Nandorfy, D. Modulation of Volatile Thiol Release during Fermentation of Red Musts by Wine Yeast. Processes 2022, 10, 502. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10030502
Cordente AG, Curtin CD, Solomon M, Kulcsar AC, Watson F, Pisaniello L, Schmidt SA, Espinase Nandorfy D. Modulation of Volatile Thiol Release during Fermentation of Red Musts by Wine Yeast. Processes. 2022; 10(3):502. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10030502
Chicago/Turabian StyleCordente, Antonio G., Christopher D. Curtin, Mark Solomon, Allie C. Kulcsar, Flynn Watson, Lisa Pisaniello, Simon A. Schmidt, and Damian Espinase Nandorfy. 2022. "Modulation of Volatile Thiol Release during Fermentation of Red Musts by Wine Yeast" Processes 10, no. 3: 502. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10030502