

Biodegradation of Chlorantraniliprole and Flubendiamide by Some Bacterial Strains Isolated from Different Polluted Sources
 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Pesticides and Media
2.2. Sampling, Enrichment, and Bacteria Isolation
2.3. Characterization of Bacterial Isolates
2.4. Molecular Identification of Isolates
2.5. Optimization of Growth Culture Conditions and Pesticide Biodegradation
2.6. Laboratory-Scale Bioremediation by Determine Microbial CO2 Production
2.7. Biodegradation of Different Pesticide Groups
2.8. Statistical Examinations
3. Results
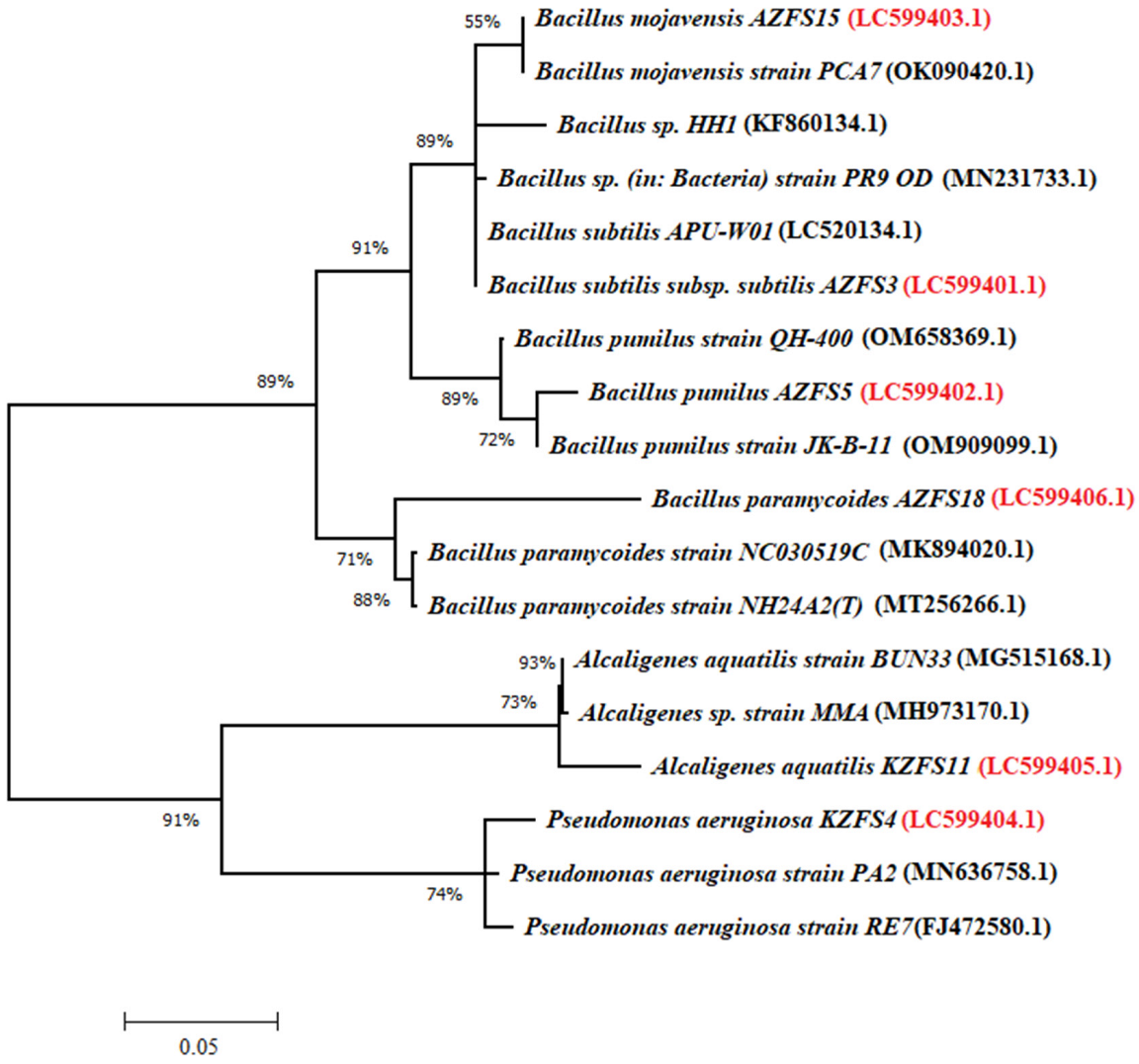
3.1. Identification of Bacterial Strains
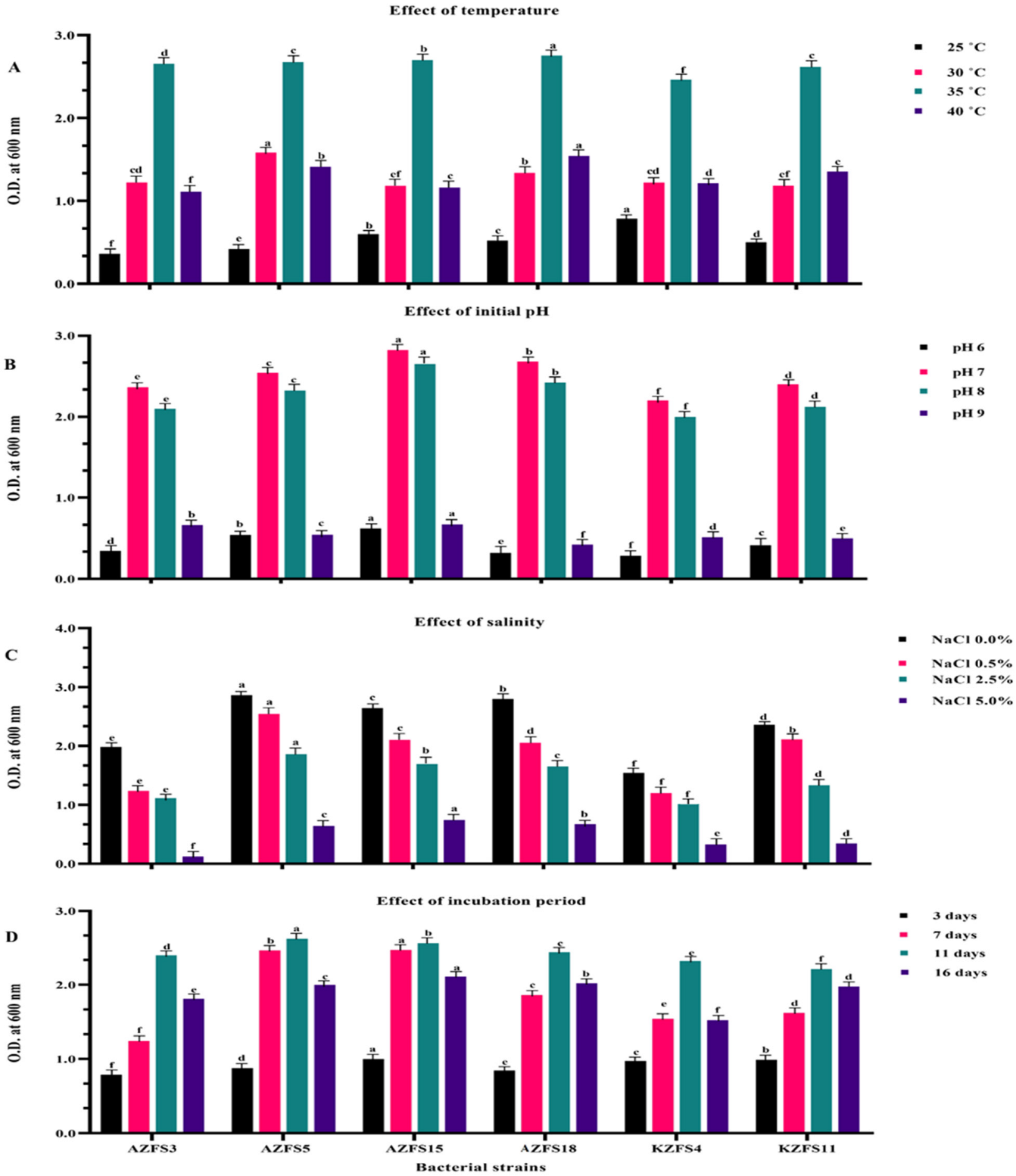
3.2. Factors Affecting Bacterial Growth through Biodegradation of Diamide Pesticides
3.2.1. Effect of Temperature
3.2.2. Effect of Initial pH
3.2.3. Salinity Effect
3.2.4. Effect of Incubation Period
3.3. Changes in the Amount of CO2 Evolution
3.4. Biodegradation of Diamide Pesticides
4. Conclusions
Author Contributions
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raffa, C.M.; Chiampo, F. Bioremediation of agricultural soils polluted with pesticides: A review. Bioengineering 2021, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. Rome. 2018. Available online: http://faostat.fao.org (accessed on 12 March 2018).
- Faostat, F. Food and Agriculture Organization of the United Nations-Statistic Division. 2019. Available online: https://www.fao.org/faostat/en/#data (accessed on 15 November 2022).
- Gonçalves, C.R.; da Silva Delabona, P. Bioremediation of pesticides in Brazil: A brief overview. Environ. Adv. 2022, 8, 100220. [Google Scholar] [CrossRef]
- Boedeker, W.; Watts, M.; Clausing, P.; Marquez, E. The global distribution of acute unintentional pesticide poisoning: Estimations based on a systematic review. BMC Public Health 2020, 20, 1875. [Google Scholar] [CrossRef] [PubMed]
- Calvert, G.M.; Mehler, L.N.; Alsop, J.; De Vries, A.L.; Besbelli, N. Surveillance of pesticide-related illness and injury in humans. In Hayes’ Handbook of Pesticide Toxicology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 1313–1369. [Google Scholar]
- Xing, H.; Li, S.; Wang, Z.; Gao, X.; Xu, S.; Wang, X. Histopathological changes and antioxidant response in brain and kidney of common carp exposed to atrazine and chlorpyrifos. Chemosphere 2012, 88, 377–383. [Google Scholar] [CrossRef] [PubMed]
- David, M.; Marigoudar, S.R.; Patil, V.K.; Halappa, R. Behavioral, morphological deformities and biomarkers of oxidative damage as indicators of sublethal cypermethrin intoxication on the tadpoles of D. melanostictus (Schneider, 1799). Pestic. Biochem. Phys. 2012, 103, 127–134. [Google Scholar] [CrossRef]
- Ellgehausen, H.; Guth, J.A.; Esser, H.O. Factors determining the bioaccumulation potential of pesticides in the individual compartments of aquatic food chains. Ecotoxicol. Environ. Saf. 1980, 4, 134–157. [Google Scholar] [CrossRef]
- David, M.; Sangeetha, J.; Shrinivas, J.; Harish, E.; Naik, V. Effects of deltamethrin on haematological indices of indian major carp, Cirrhinus mrigala (Hamilton). Int. J. Pure Appl. Zool. 2015, 3, 37–43. [Google Scholar]
- Majumdar, B.B.; Guha, G.; Ray, A.N.; Bala, B. Toxic intracerebral demyelination in a case of suicidal Cypermethrin poisoning. Ann. Trop. Med. Public Health 2012, 5, 615. [Google Scholar] [CrossRef]
- Özkara, A.; Akyıl, D.; Konuk, M. Pesticides, environmental pollution, and health. In Environmental Health Risk-Hazardous Factors to Living Species; IntechOpen: London, UK, 2016. [Google Scholar]
- Ishaaya, I. Biochemical Sites of Insecticide Action and Resistance; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef] [Green Version]
- Aghris, S.; Alaoui, O.T.; Laghrib, F.; Farahi, A.; Bakasse, M.; Saqrane, S.; Lahrich, S.; Mhammedi, E. Extraction and determination of flubendiamide insecticide in food samples: A review. Curr. Res. Food Sci. 2022, 5, 401–413. [Google Scholar] [CrossRef]
- Ortiz-Hernández, M.L.; Rodríguez, A.; Sánchez-Salinas, E.; Castrejón-Godínez, M.L. Bioremediation of soils contaminated with pesticides: Experiences in Mexico. In Bioremediation in Latin America; Springer: Berlin/Heidelberg, Germany, 2014; pp. 69–99. [Google Scholar]
- Bhatt, P.; Bhatt, K.; Sharma, A.; Zhang, W.; Mishra, S.; Chen, S. Biotechnological basis of microbial consortia for the removal of pesticides from the environment. Crit. Rev. Biotechnol. 2021, 41, 317–338. [Google Scholar] [CrossRef]
- Hussain, S.; Siddique, T.; Arshad, M.; Saleem, M. Bioremediation and phytoremediation of pesticides: Recent advances. Crit. Rev. Environ. Sci. Technol. 2009, 39, 843–907. [Google Scholar] [CrossRef]
- Obaid, R.J. Synthesis and biological evaluation of some new imidazo[1,2-c]pyrimido [5,4-e]pyrimidin-5-amine derivatives. J. Umm Al-Qura Univ. Appl. Sci. 2021, 7, 16–22. [Google Scholar]
- Gao, W.; Li, D.; You, H. Functional characterization and genomic analysis of the chlorantraniliprole-degrading strain Pseudomonas sp. GW13. Bioengineering 2019, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Atlas, R.M.; Synder, J.W. Reagents, stains, and media: Bacteriology. In Manual of Clinical Microbiology, 11th ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2011; pp. 272–303. [Google Scholar]
- Setlhare, B.; Kumar, A.; Aregbesola, O.; Mokoena, M.; Olaniran, A. 2, 4-dichlorophenol degradation by indigenous Pseudomonas sp. PKZNSA and Klebsiella pneumoniae KpKZNSA: Kinetics, enzyme activity and catabolic gene detection. Appl. Biochem. Microbiol. 2021, 57, 656–665. [Google Scholar] [CrossRef]
- Logan, N.A.; De Vos, P. Genus I. bacillus. Bergey’s Man. Syst. 2009, 3, 21–128. [Google Scholar]
- Kersters, K.; De Ley, J. Genus Alcaligenes Castellani and Chalmers 1919, 936AL. In Bergey’s Manual of Systematic Bacteriology, 9th ed.; LWW Publication: Philadelphia, PA, USA, 1992; Volume 1, pp. 361–373. [Google Scholar]
- Garrity, G.M.; Brenner, D.J.; Krieg, N.; Staley, J. Bergey’s Manual of Systematic Bacteriology. The Proteobacteria, Part C: The Alpha-, Beta-, Delta-, and Epsilonproteobacteria, Springer: New York, NY, USA, 2005.
- Nacef, M.; Chevalier, M.; Chollet, S.; Drider, D.; Flahaut, C. MALDI-TOF mass spectrometry for the identification of lactic acid bacteria isolated from a French cheese: The maroilles. Int. J. Food Microbiol. 2017, 247, 2–8. [Google Scholar] [CrossRef]
- Sauer, S.; Kliem, M. Mass spectrometry tools for the classification and identification of bacteria. Nat. Rev. Microbiol. 2010, 8, 74–82. [Google Scholar] [CrossRef]
- Biswas, S.; Rolain, J.-M. Use of MALDI-TOF mass spectrometry for identification of bacteria that are difficult to culture. J. Microbiol. Methods 2013, 92, 14–24. [Google Scholar] [CrossRef]
- Stevenson, L.G.; Drake, S.K.; Murray, P.R. Rapid identification of bacteria in positive blood culture broths by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2010, 48, 444–447. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.; Maniatis, T. Molecular Cloning. A Laboratory Manual, 2nd ed.; Cold Spring Harbor, Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Srivastava, S.; Singh, V.; Kumar, V.; Verma, P.C.; Srivastava, R.; Basu, V.; Gupta, V.; Rawat, A.K. Identification of regulatory elements in 16S rRNA gene of Acinetobacter species isolated from water sample. Bioinformation 2008, 3, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: A justification. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ouided, B.; Abderrahmane, B. Isolation and characterization of glyphosate-degrading bacteria from different soils of Algeria. Afr. J. Microbiol. Res. 2013, 7, 5587–5595. [Google Scholar] [CrossRef] [Green Version]
- John, E.M.; Sreekumar, J.; Jisha, M. Optimization of chlorpyrifos degradation by assembled bacterial consortium using response surface methodology. Soil Sediment Contam Int. J. 2016, 25, 668–682. [Google Scholar] [CrossRef]
- Dubey, S.; Upadhyay, S.N.; Sharma, Y.C. Optimization of removal of Cr by γ-alumina nano-adsorbent using response surface methodology. Ecol. Eng. 2016, 97, 272–283. [Google Scholar] [CrossRef]
- Isermeyer, H. Eine einfache Methode zur bestimmung der bodenatmung und der karbonate im boden. Z. Für. Pflanz. Düngung Bodenkd. 1952, 56, 26–38. [Google Scholar] [CrossRef]
- Mukherjee, I.; Das, S.K.; Kumar, A. Degradation of flubendiamide as affected by elevated CO2, temperature, and carbon mineralization rate in soil. Environ. Sci. Pollut. Res. 2016, 23, 19931–19939. [Google Scholar] [CrossRef]
- Wu, M.; Li, G.; Chen, X.; Liu, J.; Liu, M.; Jiang, C.; Li, Z. Rational dose of insecticide chlorantraniliprole displays a transient impact on the microbial metabolic functions and bacterial community in a silty-loam paddy soil. Sci. Total Environ. 2018, 616, 236–244. [Google Scholar] [CrossRef]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H. Methods of Soil Analysis, Part 3: Chemical Methods; John Wiley & Sons: Hoboken, NJ, USA, 2020; Volume 14. [Google Scholar]
- Page, A.; Miller, R.; Keeney, D. Methods of Soil Analysis, part 2. Chem. Microbiol. Prop. 1982, 2, 643–698. [Google Scholar]
- Malatova, K. Isolation and Characterization of Hydrocarbon Degrading Bacteria from Environmental Habitats in Western New York State. Master’s Thesis, Rochester Institute of Technology, Rochester, NY, USA, November 2005. [Google Scholar]
- Jadhav, S.S.; David, M. Biodegradation of flubendiamide by a newly isolated Chryseobacterium sp. strain SSJ1. 3 Biotech 2016, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Geed, S.; Kureel, M.; Shukla, A.; Singh, R.; Rai, B. Biodegradation of malathion and evaluation of kinetic parameters using three bacterial species. Resour.-Effic. Technol. 2016, 2, S3–S11. [Google Scholar] [CrossRef]
- Bille, E.; Dauphin, B.; Leto, J.; Bougnoux, M.-E.; Beretti, J.-L.; Lotz, A.; Suarez, S.; Meyer, J.; Join-Lambert, O.; Descamps, P. MALDI-TOF MS Andromas strategy for the routine identification of bacteria, mycobacteria, yeasts, Aspergillus spp. and positive blood cultures. Clin. Microbiol. Infect 2012, 18, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Dubois, D.; Leyssene, D.; Chacornac, J.P.; Kostrzewa, M.; Schmit, P.O.; Talon, R.; Bonnet, R.; Delmas, J. Identification of a variety of Staphylococcus species by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2010, 48, 941–945. [Google Scholar] [CrossRef] [Green Version]
- William, B.W.; Paul, D.V.; George, M.G.; Dorothy, J.; Noel, R.K.; Wolfgang, L.; Fred, A.R.; Karl, S. Bergeys Manual of Systematic Bacteriology; LWW Publication: Philadelphia, PA, USA, 2012; Volume 171, p. 190. [Google Scholar]
- Moussaoui, W.; Jaulhac, B.; Hoffmann, A.-M.; Ludes, B.; Kostrzewa, M.; Riegel, P.; Prévost, G. Matrix-assisted laser desorption ionization time-of-flight mass spectrometry identifies 90% of bacteria directly from blood culture vials. Clin. Microbiol. Infect. 2010, 16, 1631–1638. [Google Scholar] [CrossRef] [Green Version]
- Bizzini, A.; Durussel, C.; Bille, J.; Greub, G.; Prod’Hom, G. Performance of matrix-assisted laser desorption ionization-time of flight mass spectrometry for identification of bacterial strains routinely isolated in a clinical microbiology laboratory. J. Clin. Microbiol. 2010, 48, 1549–1554. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.R.; Qian, C.; Cui, S.H.; Li, F.Q. Characterization of Staphylococcus aureus isolated from clinical specimens by matrix assisted laser desorption/ionization time-of-flight mass spectrometry. Biomed. Environ. Sci. 2013, 26, 430–436. [Google Scholar]
- Teramoto, K.; Kitagawa, W.; Sato, H.; Torimura, M.; Tamura, T.; Tao, H. Phylogenetic analysis of Rhodococcus erythropolis based on the variation of ribosomal proteins as observed by matrix-assisted laser desorption ionization-mass spectrometry without using genome information. J. Biosci. Bioeng. 2009, 108, 348–353. [Google Scholar] [CrossRef]
- Odukkathil, G.; Vasudevan, N. Toxicity and bioremediation of pesticides in agricultural soil. Rev. Environ. Sci. Biotechnol. 2013, 12, 421–444. [Google Scholar] [CrossRef]
- Mali, H.; Shah, C.; Patel, D.H.; Trivedi, U.; Subramanian, R. Degradation insight of organophosphate pesticide chlorpyrifos through novel intermediate 2, 6-dihydroxypyridine by Arthrobacter sp. HM01. Bioresour. Bioprocess. 2022, 9, 31. [Google Scholar] [CrossRef]
- Lin, Q.; Chen, S.; Hu, M.; Haq, M.; Yang, L.; Li, H. Biodegradation of cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge. Int. J. Environ. Sci. Technol. 2011, 8, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Siddique, T.; Okeke, B.C.; Arshad, M.; Frankenberger, W.T. Biodegradation kinetics of endosulfan by Fusarium ventricosum and a Pandoraea species. J. Agric. Food Chem. 2003, 51, 8015–8019. [Google Scholar] [CrossRef] [PubMed]
- Philip, L. Bioremediation of endosulfan contaminated soil and water—Optimization of operating conditions in laboratory scale reactors. J. Hazard. Mater. 2006, 136, 354–364. [Google Scholar]
- Arshad, M.; Hussain, S.; Saleem, M. Optimization of environmental parameters for biodegradation of alpha and beta endosulfan in soil slurry by Pseudomonas aeruginosa. J. Appl. Microbiol. 2008, 104, 364–370. [Google Scholar] [CrossRef]
- Singh, B.K.; Walker, A.; Wright, D.J. Bioremedial potential of fenamiphos and chlorpyrifos degrading isolates: Influence of different environmental conditions. Soil Biol. Biochem. 2006, 38, 2682–2693. [Google Scholar] [CrossRef]
- Tao, L.; Yang, H. Fluroxypyr biodegradation in soils by multiple factors. Environ. Monit. Assess. 2011, 175, 227–238. [Google Scholar] [CrossRef]
- Farhan, M.; Ahmad, M.; Kanwal, A.; Butt, Z.A.; Khan, Q.F.; Raza, S.A.; Qayyum, H.; Wahid, A. Biodegradation of chlorpyrifos using isolates from contaminated agricultural soil, its kinetic studies. Sci. Rep. 2021, 11, 10320. [Google Scholar] [CrossRef]
- Vidali, M. Bioremediation. An overview. Pure Appl. Chem. 2001, 73, 1163–1172. [Google Scholar] [CrossRef]
- Karpouzas, D.G.; Walker, A. Factors influencing the ability of Pseudomonas putida epI to degrade ethoprophos in soil. Soil Biol. Biochem. 2000, 32, 1753–1762. [Google Scholar] [CrossRef]
- Mohan, S.V.; Sirisha, K.; Rao, R.S.; Sarma, P. Bioslurry phase remediation of chlorpyrifos contaminated soil: Process evaluation and optimization by Taguchi design of experimental (DOE) methodology. Ecotoxicol. Environ. Saf. 2007, 68, 252–262. [Google Scholar] [CrossRef]
- Sidal, U.; Yilmaz, E.S. Production of rhamnolipid (A Biosurfactant) using free and immobilized cells of Pseudomonas sp. Kafkas Univ. Vet. Fak. Derg. 2012, 18, 285–289. [Google Scholar]
- Vecino Bello, X.; Devesa-Rey, R.; Cruz, J.M.; Moldes, A.B.n. Study of the synergistic effects of salinity, pH, and temperature on the surface-active properties of biosurfactants produced by Lactobacillus pentosus. J. Agric. Food Chem. 2012, 60, 1258–1265. [Google Scholar] [CrossRef]
- Saikia, R.R.; Deka, S.; Deka, M.; Sarma, H. Optimization of environmental factors for improved production of rhamnolipid biosurfactant by Pseudomonas aeruginosa RS29 on glycerol. J. Basic Microbiol. 2012, 52, 446–457. [Google Scholar] [CrossRef]
- Al-Araji, L.I.Y.; Issa, L. Biosurfactant Production by Pseudomonas aeruginosa 181. Ph.D. Thesis, Universiti Putra Malaysia, Seri Kembangan, Malaysia, 2004. [Google Scholar]
- Cycoń, M.; Wójcik, M.; Piotrowska-Seget, Z. Biodegradation of the organophosphorus insecticide diazinon by Serratia sp. and Pseudomonas sp. and their use in bioremediation of contaminated soil. Chemosphere 2009, 76, 494–501. [Google Scholar] [CrossRef]
- Abo-Amer, A.E. Biodegradation of diazinon by Serratia marcescens DI101 and its use in bioremediation of contaminated environment. J. Microbiol. Biotechnol. 2011, 21, 71–80. [Google Scholar] [CrossRef]
- Jing, X.; Yang, J.; Wang, T. Effects of salinity on herbicide lactofen residues in soil. Water Air Soil Pollut. 2018, 229, 3. [Google Scholar] [CrossRef]
- Yun, E.-Y.; Ro, H.-M.; Lee, G.-T.; Choi, W.-J. Salinity effects on chlorpyrifos degradation and phosphorus fractionation in reclaimed coastal tideland soils. Geosci. J. 2010, 14, 371–378. [Google Scholar] [CrossRef]
- de Souza Silva, C.M.M.; Fay, E.F. Effect of salinity on soil microorganisms. Soil Health Land Use Manag. 2012, 10, 177–198. [Google Scholar]
- Minai-Tehrani, D.; Herfatmanesh, A.; Azari-Dehkordi, F.; Minuoi, S. Effect of salinity on biodegradation of aliphatic fractions of crude oil in soil. Pak. J. Biol. Sci. 2006, 9, 1531–1535. [Google Scholar] [CrossRef] [Green Version]
- Mingelgrin, U.; Yaron, B. The effect of calcium salts on the degradation of parathion in sand and soil. Soil Sci. Soc. Am. J. 1974, 38, 914–917. [Google Scholar] [CrossRef]
- Hussain, S.; Arshad, M.; Saleem, M.; Khalid, A. Biodegradation of α-and β-endosulfan by soil bacteria. Biodegradation 2007, 18, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Saxena, S.; Datta, A.; Arora, S. Spectrophotometric analysis of degradation of chlorpyrifos pesticide by indigenous microorganisms isolated from affected soil. Int. J. Curr. Microbiol. App. Sci. 2016, 5, 742–749. [Google Scholar] [CrossRef]
- Doolotkeldieva, T.; Konurbaeva, M.; Bobusheva, S. Microbial communities in pesticide-contaminated soils in Kyrgyzstan and bioremediation possibilities. Environ. Sci. Pollut. Res. 2018, 25, 31848–31862. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Dong, F.; Lei, X. Microbial resources and ecology-microbial degradation of pesticides. Nat. Res. Conserv. Res. 2018, 1, 242. [Google Scholar] [CrossRef]
- Gilani, R.A.; Rafique, M.; Rehman, A.; Munis, M.F.H.; ur Rehman, S.; Chaudhary, H.J. Biodegradation of chlorpyrifos by bacterial. J. Basic Microbiol. 2015, 55, 1–15. [Google Scholar]
- Feng, Y.; Minard, R.D.; Bollag, J.M. Photolytic and microbial degradation of 3,5,6-trichloro-2-pyridinol. Environ. Toxicol. Chem. Int. J. 1998, 17, 814–819. [Google Scholar] [CrossRef]
- Voos, G.; Groffman, P. Relationships between microbial biomass and dissipation of 2,4-D and dicamba in soil. Biol. Fertil. Soils 1997, 24, 106–110. [Google Scholar] [CrossRef]
- Anderson, J. Herbicide degradation in soil: Influence of microbial biomass. Soil Biol. Biochem. 1984, 16, 483–489. [Google Scholar] [CrossRef]
- Sabaté, J.; Vinas, M.; Solanas, A. Laboratory-scale bioremediation experiments on hydrocarbon-contaminated soils. Int. Biodeterior. Biodegrad. 2004, 54, 19–25. [Google Scholar] [CrossRef]
- Xu, B.; Xue, R.; Zhou, J.; Wen, X.; Shi, Z.; Chen, M.; Xin, F.; Zhang, W.; Dong, W.; Jiang, M. Characterization of acetamiprid biodegradation by the microbial consortium ACE-3 enriched from contaminated soil. Front. Microbiol. 2020, 11, 1429. [Google Scholar] [CrossRef]
- Pino, N.; Peñuela, G. Simultaneous degradation of the pesticides methyl parathion and chlorpyrifos by an isolated bacterial consortium from a contaminated site. Int. Biodeterior. Biodegrad. 2011, 65, 827–831. [Google Scholar] [CrossRef]
- Randika, J.; Bandara, P.; Soysa, H.; Ruwandeepika, H.; Gunatilake, S. Bioremediation of pesticide-contaminated soil: A review on indispensable role of soil bacteria. J. Agric. Sci.-Sri Lanka 2022, 17, 19–43. [Google Scholar] [CrossRef]
- Góngora-Echeverría, V.R.; García-Escalante, R.; Rojas-Herrera, R.; Giácoman-Vallejos, G.; Ponce-Caballero, C. Pesticide bioremediation in liquid media using a microbial consortium and bacteria-pure strains isolated from a biomixture used in agricultural areas. Ecotoxicol. Environ. Saf. 2020, 200, 110734. [Google Scholar] [CrossRef]
- McAllister, K.A.; Lee, H.; Trevors, J.T. Microbial degradation of pentachlorophenol. Biodegradation 1996, 7, 1–40. [Google Scholar] [CrossRef]
- Geetha, M.; Fulekar, M. Bioremediation of pesticides in surface soil treatment unit using microbial consortia. Afr. J. Environ. Sci. Technol. 2008, 2, 036–045. [Google Scholar]
- Werren, J.H. Symbionts provide pesticide detoxification. Proc. Natl. Acad. Sci. USA 2012, 109, 8364–8365. [Google Scholar] [CrossRef] [Green Version]
- Naphade, S.R.; Durve, A.A.; Bhot, M.; Varghese, J.; Chandra, N. Isolation, characterization and identification of pesticide tolerating bacteria from garden soil. Eur. J. Exp. Biol. 2012, 2, 1943–1951. [Google Scholar]
- Javaid, M.K.; Ashiq, M.; Tahir, M. Potential of biological agents in decontamination of agricultural soil. Scientifica 2016, 2016, 1598325. [Google Scholar] [CrossRef] [Green Version]
- Mulbry, W.W.; Karns, J.S.; Kearney, P.; Nelson, J.; McDaniel, C.; Wild, J. Identification of a plasmid-borne parathion hydrolase gene from Flavobacterium sp. by southern hybridization with opd from Pseudomonas diminuta. Appl. Environ. Microbiol. 1986, 51, 926–930. [Google Scholar] [CrossRef] [PubMed]
- Malghani, S.; Chatterjee, N.; Yu, H.X.; Luo, Z. Isolation and identification of profenofos degrading bacteria. Braz. J. Microbiol. 2009, 40, 893–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. The WHO Recommended Classification of Pesticides by Hazard and Guidelines to Classification 2009; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Ghani, M.U.; Asghar, H.N.; Niaz, A.; Ahmad Zahir, Z.; Nawaz, M.F.; Häggblom, M.M. Efficacy of rhizobacteria for degradation of profenofos and improvement in tomato growth. Int. J. Phytoremediation 2022, 24, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Kadhim, F.; Rabee, A.M.; Abdalraheem, E. Biodegradation of malathion by selected bacterial isolates. J. Int. Environ. Appl. Sci. 2015, 10, 361–366. [Google Scholar]



{kind=link}
{kind=link}
| Characters | Values |
|---|---|
| Physical analysis | |
| Clay% | 12.1 |
| Silt% | 10.9 |
| Sand% | 77.0 |
| Textural class | Sandy loam |
| CaCO3 (g kg−1) | 11.5 |
| Chemical analysis | |
| pH (1:2.5 at 25 °C) | 8.09 |
| EC dSm−1 (1:5 at 25 °C) * | 0.52 |
| Na+ | 0.30 |
| K+ | 1.30 |
| Ca2+ | 1.60 |
| Mg2+ | 2.22 |
| CO32− | Nil |
| HCO3 | 2.30 |
| Cl− | 0.66 |
| SO42− | 2.44 |
| Organic matter (g kg−1) | 9.40 |
| Available nutrients (mg kg−1 soil) | |
| N | 60.50 |
| P | 18.6 |
| K | 90.5 |
| Isolates Code | AZFS3 | AZFS5 | AZFS15 | AZFS18 | KZFS4 | KZFS11 |
|---|---|---|---|---|---|---|
| 1-Gram reaction | + | + | + | + | − | − |
| 2-Cell shape | L-Rod | L-Rod | L-Rod | L-Rod | S-rod | S-rod |
| 3-Motility | + | + | + | − | + | + |
| 4-Spore formation | + | + | + | NO | - | − |
| 5-Oxygen requirement Aerobic growth Anaerobic growth | + − | + − | + − | + +/− | + − | + − |
| 6-Voges-proskauer | + | + | + | + | − | − |
| 7-Catalase reaction | + | + | + | − | + | + |
| 8-Oxidase reaction | − | + | + | − | + | + |
| 9-Urease | − | − | − | − | − | − |
| 10-Nitrate reduction | + | − | + | + | + | + |
| 11-Uitlization of • Citrate • Propionate | + − | + − | + − | + + | + ND | − − |
| 12-Hydrolysis of -Casein -Gelatin -Starch | + + + | + + − | ND + | + + + | ND + ND | − − − |
| 13-Growth in media with • 0% NaCl • 2% NaCl • 5% NaCl • −7% NaCl • −10% NaCl | + + + + ND | + + + + + | + + + + + | + + + + + | + + + − − | + + + + + |
| 14-Growth at pH • 5.0 • 6.0 • 7.0 • 8.0 • 9.0 • 10.0 | + + + + + − | + + + + + − | + + + + + − | + + + + + − | + + + + + − | − + + + − |
| 15-Growth at • 20 °C • 30 °C • 40 °C • 50 °C • 60 °C | + + + −− | + + + −− | + + + + | + + + −− | + + + −− | + + + −− |
| 16-Acid production from (1) L-Arabinose (2) D-Glucose (3) Glycogen (4) D-Mannitol (5) D-Mannose (6) L-Rhamnose (7) Starch (8) D-Xylose (9) Trehalose (10) Inulin | + + + + + −+ + + − | + + + + + − | + + ND + + + ND + + ND | + + + + d + + − | −+ ND + − | + + + −ND + ND ND ND ND |
| No. | Isolate Code | Analyte Name | Organism (Best Match) | Score Value |
|---|---|---|---|---|
| 1 | AZFS3 | C1 (+++) | Bacillus subtilis subsp. subtilis DSM10 | 2.121 |
| 2 | AZFS5 | C2 (+++) | Bacillus pumilus BB08-1 | 2.332 |
| 3 | AZFS15 | C3 (++) | Bacillus mojavensis DSM 9205 | 2.141 |
| 4 | KZFS18 | C6 (+++) | Bacillus paramycoides BY9 | 2.323 |
| 5 | KZFS4 | C4 (++) | Pseudomonas aeruginosa F1 | 2.129 |
| 6 | KZFS11 | C5 (++) | Alcaligenes aquatilis QD168 | 2.120 |
| Source of Soil | ID Sample | Name of Strain | Accessions Numbers | Identity (%) |
|---|---|---|---|---|
| AZF | AZFS3 | Bacillus subtilis subsp. subtilis AZFS3 | LC599401.1 | 100 |
| AZFS5 | Bacillus pumilus AZFS5 | LC599402.1 | 100 | |
| AZFS15 | Bacillus mojavensis AZFS15 | LC599403.1 | 100 | |
| AZFS18 | Bacillus paramycoides AZFS18 | LC599406.1 | 100 | |
| KZF | KZFS4 | Pseudomonas aeruginosa KZFS4 | LC599404.1 | 100 |
| KZFS11 | Alcaligenes aquatilis KZFS11 | LC599405.1 | 100 |
| Bacterial Strain | Incubation at 28 °C for 16 Days | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 Days | 7 Days | 11 Days | 16 Days | mg CO2/16 Day | % *** | ||||||
| T. * | FBD-CAP ** | T. | FBD-CAP | T. | FBD-CAP | T. | FBD-CAP | T. | FBD-CAP | FBD-CAP | |
| AZFS3 1 | 10.103 i | 0.202 i | 14.443 e | 0.388 e | 8.021 e | 0.210 e | 1.101 a | 0.101 f | 33.668 e | 0.901 e | 57.7 |
| AZFS5 2 | 11.231 h | 0.244 h | 13.571 f | 0.368 f | 6.401 f | 0.129 f | 0.202 e | 0.122 c | 31.405 f | 0.863 g | 55.4 |
| AZFS15 3 | 15.525 f | 0.301 f | 8.122 i | 0.265 i | 5.111 i | 0.115 i | 0.233 b | 0.129 a | 28.991 i | 0.810 i | 51.9 |
| AZFS18 4 | 14.362 g | 0.298 g | 10.703 g | 0.299 g | 5.959 h | 0.121 h | 0.229 d | 0.121 d | 31.253 h | 0.839 h | 53.8 |
| KZFS4 5 | 19.125 d | 0.481 d | 17.021 d | 0.404 d | 10.005 c | 0.230 g | 0.201 f | 0.111 e | 46.352 d | 1.226 d | 78.6 |
| KZFS11 6 | 16.212 e | 0.322 e | 8.613 h | 0.277 h | 6.213 g | 0.122 f | 0.231 c | 0.123 b | 31.269 g | 0.844 f | 54 |
| Consortia (No.1) 7 | 22.125 c | 0.665 c | 20.021 c | 0.499 c | 9.005 d | 0.225 c | 0.011 h | 0.008 h | 51.162 c | 1.397 c | 89.6 |
| Consortia (No.2) 8 | 25.125 b | 0.688 b | 22.021 b | 0.515 b | 10.006 b | 0.231 b | 0.044 g | 0.012 g | 57.195 b | 1.446 b | 92.8 |
| Consortia (No.3) 9 | 29.891 a | 0.712 a | 23.912 a | 0.535 a | 16.218 a | 0.306 a | 0.0 | 0.0 | 70.021 a | 1.553 a | 99.6 |
| Bacterial Strains | Various Pesticides Types | ||||||
|---|---|---|---|---|---|---|---|
| Bacterial Growth (O.D. 600 nm) | |||||||
| Diamide Pesticide Group (1) | Other Different Pesticides Group (2) | ||||||
| FBD-CAP * | FBD | CAP | PFS | CYP | CFN | MLN | |
| Bacillus subtilis subsp. subtilis AZFS3 | 1.300 e | 1.320 e | 1.31 e | 1.33 c | 1.251 f | 1.219 g | 1.321 e |
| Bacillus pumilus AZFS5 | 1.233 f | 1.306 f | 1.356 d | 1.133 g | 1.244 g | 1.333 e | 1.245 f |
| Bacillus mojavensis AZFS15 | 1.366 d | 1.359 d | 1.300 f | 1.301 e | 1.356 e | 1.367 d | 1.131 g |
| Bacillus paramycoides AZFS18 | 1.300 e | 1.216 g | 1.311 e | 1.260 f | 1.359 d | 1.300 f | 1.329 d |
| Pseudomonas aeruginosa KZFS4 | 1.610 b | 1.701 b | 1.601 b | 1.599 b | 1.542 b | 1.621 b | 1.701 b |
| Alcaligenes aquatilis KZFS11 | 1.521 c | 1.455 c | 1.483 c | 1.322 d | 1.479 c | 1.433 c | 1.494 c |
| Consortium (No.3) | 2.365 a | 2.119 a | 2.234 a | 2.122 a | 2.222 a | 2.324 a | 2.311 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahmy, M.A.; Salem, S.H.; Qattan, S.Y.A.; Abourehab, M.A.S.; Ashkan, M.F.; Al-Quwaie, D.A.; Abd El-Fattah, H.I.; Akl, B.A. Biodegradation of Chlorantraniliprole and Flubendiamide by Some Bacterial Strains Isolated from Different Polluted Sources. Processes 2022, 10, 2527. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10122527
Fahmy MA, Salem SH, Qattan SYA, Abourehab MAS, Ashkan MF, Al-Quwaie DA, Abd El-Fattah HI, Akl BA. Biodegradation of Chlorantraniliprole and Flubendiamide by Some Bacterial Strains Isolated from Different Polluted Sources. Processes. 2022; 10(12):2527. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10122527
Chicago/Turabian StyleFahmy, Mohamed A., Samir H. Salem, Shaza Y. A. Qattan, Mohammed A. S. Abourehab, Mada F. Ashkan, Diana A. Al-Quwaie, Hassan I. Abd El-Fattah, and Behairy A. Akl. 2022. "Biodegradation of Chlorantraniliprole and Flubendiamide by Some Bacterial Strains Isolated from Different Polluted Sources" Processes 10, no. 12: 2527. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10122527