

Production and Recovery of Ectoine: A Review of Current State and Future Prospects

1
Centre for Research and Graduate Studies, University of Cyberjaya, Persiaran Bestari, Cyberjaya 63000, Selangor, Malaysia
2
Biorefinery and Bioprocess Engineering Laboratory, Department of Chemical Engineering and Material Science, Yuan Ze University, Taoyuan 320, Taiwan
3
Department of Chemical Science and Engineering, Graduate School of Engineering, Kobe University, 1-1 Rokkodai, Nada 657-8501, Japan
4
Department of Chemical Engineering, National Cheng Kung University, Tainan 701, Taiwan
5
Department of Chemical and Materials Engineering, Tunghai University, Taichung 407, Taiwan
6
Research Center for Smart Sustainable Circular Economy, Tunghai University, Taichung 407, Taiwan
7
Graduate School of Biotechnology and Bioengineering, Yuan Ze University, Taoyuan 320, Taiwan
*
Author to whom correspondence should be addressed.
Processes 2023, 11(2), 339; https://0-doi-org.brum.beds.ac.uk/10.3390/pr11020339
Submission received: 31 October 2022
/
Revised: 9 January 2023
/
Accepted: 16 January 2023
/
Published: 20 January 2023
(This article belongs to the Special Issue Technologies for Production, Processing, and Extractions of Nature Product Compounds)
Abstract
:Ectoine (1,4,5,6-tetrahydro-2-methyl-4-pyrimidinecarboxylic acid) is a revolutionizing substance with vast applications in the cosmetic and food industries. Ectoine is often sourced from halobacteria. The increasing market demand for ectoine has urged the development of cost-effective and sustainable large-scale production of ectoine from microbial sources. This review describes the existing and potential microbial sources of ectoine and its derivatives, as well as microbial production and fermentation approaches for ectoine recovery. In addition, conventional methods and emerging technologies for enhanced production and recovery of ectoine from microbial fermentation with a focus on the aqueous biphasic system (ABS) are discussed. The ABS is a practically feasible approach for the integration of fermentation, cell disruption, bioconversion, and clarification of various biomolecules in a single-step operation. Nonetheless, the implementation of the ABS on an industrial-scale basis for the enhanced production and recovery of ectoine is yet to be exploited. Therefore, the feasibility of the ABS to integrate the production and direct recovery of ectoine from microbial sources is also highlighted in this review.
1. Introduction
True halophiles such as halophilic archaea and some halophilic bacteria are adapted to a high-salt environment, with well-adapted enzymes and structural components and the accumulation of inorganic ions in the cytoplasm [1]. Meanwhile, some halophilic and halotolerant microorganisms synthesize low-molecular-weight organic solutes in response to high osmotic stress, high turgor pressure, or a high-salt external environment to maintain osmotic equilibrium [1,2]. The accumulation of these organic compatible solutes can stabilize the cytoplasmic and the whole cells’ structural components against extreme conditions by enhanced osmotic activity [3]. Compatible solutes are widely applied in various industries due to their strong stabilizing capability, especially in the medicinal, pharmaceutical, cosmeceutical, and agricultural industries.
Ectoine (CAS No.: 96702-03-3), chemically known as 1,4,5,6-tetrahydro-2-methyl-4-pyrimidinecarboxylic acid, is a heterocyclic amino acid derivative of aspartate with a zwitterionic structure. Ectoine was first identified and isolated from a halophilic bacterium, Ectothiorhodospira halochloris [4]. In later years, ectoine was discovered in several naturally occurring aerobic chemoheterotrophic bacteria [5]. Ectoine’s biosynthetic gene was cloned and functionally expressed for the first time in Escherichia coli by Louis and Galinski in 1997 and later by He et al. (2006) with the advancement of genetic engineering, which has marked a great breakthrough in the sustainable production of ectoine [6,7].
Ectoine is synthesized by halophilic bacteria via the amino acid biosynthesis of the aspartate family with the enzymatic reactions of three main enzymes: l-2, 4-diaminobutyrate transaminase (ectB), l-2, 4-diaminobutyrate acetyltransferase (ectA), and ectoine synthase (ectC) [8,9]. Several halophilic bacteria inclusive of Chromohalobacter salexigens and Halomonas elongata tend to synthesize hydroxyectoine via a hydroxylation reaction in the presence of the enzyme ectoine hydroxylase (ectD) [10]. A high-salt environment often triggers the expression of these ectoine-synthesizing enzymes, resulting in the synthesis of high concentrations of ectoine.
Ectoine, with its strong stabilizing capability, often acts as a stress and thermal protectant for halophilic bacteria such as C. salexigens and Virgibacillus pantothenticus against extreme freezing, drying, and heating conditions [1]. Ectoine provides osmotic balance to microbial cells without disrupting the cellular metabolic processes of the microorganisms. The zwitterionic ectoine exerts a negligible effect on the ionic strength of the cytoplasm of the microorganism; therefore, there is no imperative modification on the intracellular enzymes and organelles [11,12]. The concentration of the ectoine accumulated intracellularly is often dependent upon the surrounding environmental osmolarity [12]. For instance, a hypo-osmotic environment induces the synthesis of ectoine through the unique efflux system which regulates the internal osmolarity of microorganisms by increasing the intracellular solutes pool and vice versa [12].
Ectoine is among the most extensively used compatible solutes for various commercial applications due to the intensely powerful stabilizing capability shown [13]. Ectoine is an excellent stabilizer for intracellular macromolecules enzymes, nucleic acids, and various proteins [12,14]. Ectoine alters the solvent properties by preferentially excluding the solutes from the enzyme surface, resulting in a more compact conformation of the enzyme [15]. In addition, the accumulation of ectoine also enhances the cellular hydration and the turgor maintenance under unfavorable conditions. In the context of stabilization of the nucleic acid structure, ectoine acts to trigger the structural change of the nucleic acids which can restrict the cleavage reactions by most of the endonucleases [12].
This review discusses the current state of ectoine production and the emerging technology for the improved downstream processing of ectoine sourced from microbial fermentation with the aqueous biphasic system (ABS). In addition, the microbial sources of ectoine and the vast applications of ectoine are also summarized.
There is a vast number of microorganisms synthesizing ectoine, which include the members of halobacteria, actinobacteria, and firmicutes under different stress conditions [12]. Ectoine is also found in some methylotrophic bacteria with a four-gene cluster of greater halotolerance, which synthesize ectoine independent of other amino acid biosynthetic pathways. For instance, methylotrophic bacteria which synthesizes ectoine include the members of the genera Methylomicrobium, Methylobacter, and Methylophaga [16,17,18]. Furthermore, several species of Archaea also produce ectoine and hydroxyectoine [19].
The most common ectoine-synthesizing halobacteria which are applied as industrial producers are of the members of Halomonas such as Halomonas salina [20], Halomonas boliviensis [21], and Halomonas elongata [22]. In addition, some of the ectoine-synthesizing bacteria such as Brevibacterium album, Marinococcus halotolerans, Virgibacillus salexigens, and Halomonas sp. also co-synthesize hydroxyectoine under certain stresses with the presence of the enzyme ectD [23]. Studies have also shown that ectoine is found to be produced in a smaller amount in some microalgae [24] and from biogas production [25,26]. Several other common microbial ectoine producers are summarized in Table 1. In recent years, the ectoine biosynthesis gene cluster was found and identified from Aestuariispira ectoiniformans sp. nov. which was isolated from seawater [27].
The unique properties of ectoine as a stabilizer biomolecule allows the vast applications of ectoine in the field of biotechnology. For instance, ectoine enhances the catalytic efficiency of the enzyme lipase in the production of biodiesel, which can alternately improve the overall yield of the biodiesel [28]. The yield of methyl ester using the solvent-free methanolysis system consisting of cottonseed oil was significantly enhanced by 20.9% when ectoine was added to the immobilized lipase. Ectoine acts as an enzyme stabilizer which can conserve the structural conformation of the lipase and thereby enhance the overall production efficiency of the methyl ester [28]. Ectoine as an osmolyte was also shown to have protective effects on several enzymes against freezing, heating, drying, and proteolysis which were triggered by trypsin and trypsinogen [12].
Ectoine has been widely applied in the cosmetic industry over the past decades because of its protective and stabilizing effects on human skin cell membranes against damaging external factors such as ultraviolet (UV) radiation, wind, humidity, and drastic temperatures [12,29]. Ectoine can prevent various skin diseases such as photocarcinogenesis, photodermatoses, and photoaging, which are often associated to long-term exposure to ultraviolet A (UV-A) by protecting the skin cells with its singlet oxygen-quenching properties and delaying the process of skin aging [30]. Ectoine was also shown to prevent water loss from the skin barrier and maintain skin hydration in previous examples in the literature [31]. As a result, Marini et al., 2014, have formulated an ectoine-containing cream as a topical treatment for patients with atopic dermatitis (AD), which was often associated to deteriorated skin barrier function and skin dryness. Results revealed that ectoine is a novel and potent substance to treat AD when compared to existing available reference products [32]. Ectoine was also found to disperse keratin bundles in human stratum corneum in the latest literature [33]. Following this discovery, the popularity and acceptance of ectoine as a skincare ingredient among dermatologists and scientists were further enhanced due to its excellent moisturizing and anti-UV properties. Table 2 shows several other significant applications of ectoine in the biomedical and cosmetic industries.
Furthermore, ectoine can protect and moisturize the epithelial tissues which can reduce the inflammation on cell membranes via the “preferential exclusion” mechanism [43]. This unique mechanism exhibited by ectoine can alternately stabilize the mucous membrane and thereby prevents the invasion of potential allergens and pathogens. Therefore, Eichel et al., 2013, have developed the ectoine-containing nasal spray to treat patients with acute rhinosinusitis in ears, nose, and throat [44]. Later, in 2014, ectoine-containing nasal spray and eye drops were shown to be as effective as conventional azelastine-containing nasal spray and eye drops in treating allergic rhinitis, which affects approximately 20% of the human population in Western countries [42]. In addition, there were no adverse effects detected when patients were treated topically with ectoine-containing medical devices as compared to azelastine-containing medical devices [42]. These novel findings have marked a new milestone of discovery for ectoine’s biomedical applications.
1.1. Current State of Ectoine Production
The diverse applications of ectoine in various industries as mentioned above have urged the development of the cost-effective large-scale production of ectoine to meet the increasing market demand. Ectoine is often produced by large-scale microbial fermentation for industrial applications. Microbial ectoine fermentation generally results in low yields and, therefore, demands multiple steps of downstream processes for the purification of ectoine in order to achieve acceptably high yields [45]. Moreover, the microbial production process of ectoine is generally performed in high-salt concentration fermentation media because halophilic microorganisms require a high-salinity environment for ectoine synthesis. In most cases, the ectoine concentration synthesized by the microbial sources directly corresponds to the concentration of the salt present in the fermentation media. Therefore, a high-salt environment is often the key determinant to the efficiency of large-scale production and refinery of ectoine [2]. Nevertheless, regular usage of high salt concentration in the fermentation medium is detrimental to the fermenter in the large-scale production of ectoine due to the corrosiveness of the high salinity, and subsequently reduces the microbial cell growth and the overall production yields of the ectoine [46].
The ectoine yields of microbial fermentation are also dependent upon the types of carbon and nitrogen sources utilized in the fermentation medium. Therefore, lab-scale experiments to determine the types of carbon and nitrogen sources, as well as the types and concentrations of the salts in the fermentation medium formulation, are imperative prior to large-scale ectoine fermentation [47]. Ectoine yields can be enhanced when the fermentation media and conditions are optimized, respectively. Several strategies of ectoine production established in the previous literature are summarized in Table 3.
1.2. Intracellular Ectoine Production
Most microbial ectoines are present in the intracellular compartment. Bacterial milking has been widely used to improve intracellular ectoine production in the past two decades [12]. Bacterial milking is a common fermentation technology to produce intracellular compatible solutes in the industrial-scale production of ectoine. Bacterial milking of ectoine requires a series of hyperosmotic or hypoosmotic culture media for the continual synthesis and production of ectoine [50].
Bacterial milking for ectoine production was first used on the fed-batch fermentation of Gram-negative, proteolytic wild-type Halomonas elongata DSM 142T strain to enhance the ectoine yields for commercialization purposes [50]. H. elongata DSM 142T cells were grown in medium containing 15% (w/v) NaCl, with glucose and ammonium chloride as the respective carbon and nitrogen sources. Membrane filtration was then used to retain the bacterial cells in the fermenter and, later, the bacteria cells were exposed to an osmotic downshock with 3% (w/v) of NaCl in the growth medium to promote the synthesis of ectoine. The cells were then re-suspended in a new medium containing a 1.2-fold higher salt concentration than the initial medium, and the fermentation process was repeated. An average biomass of 155 mg of ectoine/g of dry cell weight was achieved using this repetitive bacterial fermentation process of nine cycles [50].
Bacterial milking is the most widely applied method in large-scale production of ectoine, despite several drawbacks encountered along the fermentation process. For instance, bacterial milking is highly associated with high-salt media used for the cultivation of the microorganisms, which is corrosive to the materials, equipment, as well as the bioreactor [52]. The corrosion of the equipment and the instability of materials used in the bioreactors along the microbial fermentation process remain unresolved, which resulted in the increase in the overall production cost of ectoine [46].
Intracellular ectoine can be produced via conventional fed-batch fermentation. Brevibacterium epidermis strain DSM 20659 synthesizes ectoine intracellularly using glutamate and yeast extract as the primary carbon sources with a relatively low salt concentration of 1 M NaCl as compared to the above-mentioned bacterial milking method [48]. The lower salt concentration enhances the overall productivity of ectoine and minimizes the equipment corrosion with an comparable yield of 160 mg ectoine/g of dry cell weight produced [48].
In later years, Guzman et al., 2009, demonstrated that ectoine can be produced as a by-product in the production of poly(3-hydroxybutyrate) (PHB) via two-step fed-batch Halomonas boliviensis DSM 15516 fermentation. However, the final yield of ectoine (170 mg/g dry cell weight) obtained using this co-production strategy was not significantly higher when compared to other halophilic ectoine-excreting bacterial producers such as H. elongata, which synthesizes ectoine as a major bio-product [51]. The metabolic influx would result in the preferrable synthesis of PHB or ectoine in significantly different yields by H. boliviensis DSM 15516 fermentation, as the metabolic reaction required the intermediate acetyl-CoA. Therefore, the consistency of the yields and productivity of ectoine were difficult to monitor in a large-scale fermentation process. Furthermore, additional steps in separating the ectoine from the PHB are required, which often hampered the commercialization of this co-production strategy, considering the high production cost incurred [51].
2. Extracellular Ectoine Production
The recovery of extracellular ectoine from the fermentation medium is relatively simple when compared to the recovery of intracellular ectoine from microbial cells. The production cost of industrial-scale extracellular ectoine production can be reduced with minimal usage of high salt concentrations and elimination of additional steps to extract intracellular ectoine from microbial cells [46]. However, there are finite ectoine-excreting bacterial strains known to date, and extracellular ectoine is usually produced in small quantities in fermentation broth which often demands subsequent downstream processes to concentrate and purify the ectoine produced [40].
Halomonas salina DSM 5928 is an innate ectoine-excreting strain which can synthesize ectoine extracellularly with a low salt concentration of 0.5 M NaCl [52]. The presence of glucose and yeast extract in the fermentation medium adversely affected the production yield of the extracellular ectoine. A negative correlation between the yield of the extracellular ectoine and NaCl concentration in the fermentation medium was observed, which is in contrast to most of the bacterial strains that synthesized ectoine intracellularly [52]. A combined batch-fermentation approach utilizing both growing and resting cells of H. salina DSM 5928T was developed for enhanced extracellular ectoine production (11.75 g/L) [52]. The growing cells were cultured in the optimal fermentation medium containing monosodium glutamate as the primary carbon and nitrogen source. Upon reaching the late exponential growth phase, the cells were fed with phosphate-limited medium for optimum ectoine production.
Advances of Integrated Approaches for Ectoine Fermentation
Following the advances in fermentation technology, several integrated approaches on ectoine fermentation were introduced in order to enhance the production yields of ectoine. The advances in metabolic engineering also enhance the production of ectoine in microbial fermentation [53,54]. For instance, Van-Thuoc et al., 2010, have successfully integrated the two-step fed-batch fermentation and bacterial milking of H. boliviensis LC1T cells which resulted in enhanced yields of ectoine in the second fed-batch fermentation after a high cell density was recorded in the first fed-batch H. boliviensis LC1T cell cultivation [21]. Ectoine was produced in the fermentation broth with a rate of 9.1 g/L per day in this study. However, the overall ectoine fermentation process was dependent upon the selection of the salt medium because a relatively high salt concentration of 2.55 M NaCl was demanded for the cultivation of bacterial cells [21].
A cascade of two continuously operating bioreactors for intracellular ectoine production from the moderately halophilic C. salexigens was constructed later in 2010 by Fallet et al. The first bioreactor was designed for accumulation of a high cell density of C. salexigens with an extreme pH and salt concentration, whereas, a second coupled bioreactor was engineered for the synthesis of ectoine through the osmotic and thermal downshock methods [49]. The overall ectoine yield recovered was improved by the high cell density of 61 g/L using a cross-flow ultrafiltration. A total of 540 mg ectoine/g of dry cell weight was recorded, which has outperformed the ectoine yield demonstrated by the ectoine biofactory developed by Sauer and Galinski, 1998 (155 mg ectoine/g of dry cell weight).
The yield of ectoine can be further enhanced by uncoupling the high osmolarity from the ectoine biosynthesis pathway of microorganisms via the metabolic engineering approach. Heterologous producers of ectoine have been developed by incorporating the ectoine-synthesis gene into a non-halophilic microorganism. Ectoine can be synthesized and secreted extracellularly by the heterologous ectoine producers while maintaining their biological properties when they are exposed to a new abiotic stress-resistance environment [12]. The ectoine-synthesis gene cluster sourced from a variety of halophiles such as Marinococcus halophilus and C. salexigens has been transferred to Escherichia coli in most of the metabolic engineering studies [55].
In 2007, Schubert et al. successfully transferred the ectABC gene cassette from C. salexigens to E. coli DH5α host cells using the pASK-IBA7 vector. The resulting recombinant E. coli produced extracellular ectoine in low-salinity broth, which alternately simplified the overall ectoine recovery process during industrial-scale production [56]. Becker et al., 2013, later successfully developed a recombinant Gram-positive Corynebacterium glutamicum strain by integrating the ectABCD gene cluster sourced from halophilic Pseudomonas stutzeri A1501 into the genome of C. glutamicum. In contrast to the existing halophilic extracellular ectoine-producing microbial strains, the resulting recombinant C. glutamicum excreted ectoine into the fermentation medium irrespective of the high salt concentration in the medium [57].
With the recent advances in metabolic engineering, over-expression of the ectoine gene cluster ectABCD, which is sourced from H. elongata cells, was initiated in E. coli K12 BW25113 [58]. The recombinant E. coli produced a remarkably high yield of ectoine in a fermenter, as compared to the shake-flask cultures. Repetitive cycles of ectoine fermentation using whole-cell biocatalysts were feasible by using this transgenic E. coli strain in view of the high consistency of ectoine yields recorded at the second cycle (84%) and third cycles (69%) when compared to the first ectoine-production cycle [57].
Tanimura et al., 2013, further investigated the effects of different types of sugars as carbon sources on engineered H. elongata strains for enhanced ectoine production. H. elongata mutants could serve as potential hosts for ectoine synthesis with biomass-derived lignocellulose and rice straw hydrolysate as carbon sources [58]. Inedible lignocellulose is widely available worldwide; the utilization of the lignocellulose-derived sugars as carbon sources replacing glucose in the fermentation media has facilitated a biomass-oriented sustainable society [21]. In addition, the study revealed that the yield of ectoine could be enhanced by applying rice straw hydrolysate, xylose, cellobiose, and a mixture of glucose and xylose as the carbon source. The deletion of the ectD gene from the H. elongata mutant further promoted ectoine production [58].
Following their success in 2013, Tanimura et al., 2016, further improved the production of ectoine by over-expressing the sugar transporters in the H. elongata mutant. The sugar-consumption rate of the H. elongata cells was reduced during the ectoine-producing stage to maintain osmotic balance. The majority of the sugars was consumed during the bacterial growth phase after ectoine production and a higher rate of ectoine production was recorded in the ectoine-producing phase [59]. Ectoine productivity was enhanced with the facilitation of the sugar transport from the medium into the cells.
In the latest literature, a potential synthetic cell factory for ectoine production by transferring the ectoine-synthesis gene cluster, ectABC, from halophilic H. elongata to a non-halophilic E. coli host was established [60]. The performance of E. coli in ectoine production was further improved by the deletion of the thrA gene encoding bifunctional aspartokinase (Ask) to inhibit the competitive l-threonine branch along the ectoine biosynthetic pathway. To complement the enzyme deficiency due to thrA deletion, feedback resistance of lysC sourced from Corynebacterium glutamicium was incorporated to synthesize Ask [60]. The final engineered E. coli strain could produce a total of 25.1 g/L of ectoine with a productivity of 0.84 g/L per hour through fed-batch fermentation with low salinity. However, further research on improving the metabolic network and fermentation conditions is required, because only 17% of the predicted maximum ectoine yield was obtained in this study [60].
3. Recovery and Downstream Processes of Ectoine
The ectoine fermentation process is often coupled with the production of other bioproducts such as PHB and hydroxyectoine [51]. Moreover, ectoine is preferably synthesized in the intracellular compartment of microorganisms, which further complicates the downstream processes of ectoine production. Additional multi-step separation, concentration, and clarification downstream processes are required to obtain ectoine with high yield and purity. The current state of and biotechnological advances in the recovery and downstream processes of ectoine are discussed in the following sections.
3.1. Coupled Cation-Exchange Resin and Crystallization
Sauer and Galinski, 1998, first attempted the purification of ectoine using a two-step purification strategy which consisted of a coupled cation-exchange chromatography and crystallization. Ectoine and hydroxyectoine were successfully recovered in their cationic states at a pH of 1.4 using this purification strategy. Ectoine was purified via a cation-exchange resin (Na+ form). Due to differences in ionic charges, the contaminants were removed, while the ectoine was bound to the cation-exchange resin and recovered with the addition of sodium hydroxide (NaOH). Further separation of ectoine and hydroxyectoine were achieved using methanol evaporation, followed by a water crystallization step for the recovery of ectoine [50].
3.2. Ethanol Crystallization
The recovery and purification of intracellular ectoine from the fermentation broth of Brevibacterium epidermis was extended to a two-step hypo-osmotic extraction of ectoine using water and ethanol, respectively. However, further downstream processing was required to remove the impurities and contaminants such as the remaining NaCl and glutamate to obtain a high purity of ectoine. The crude ectoine was then concentrated using evaporation with a reduced pressure until a brownish precipitate was obtained. Ethanol was later used to re-crystallize the recovered ectoine [48].
Ethanol crystallization was applied as the initial purification step to recover ectoine. Wei et al., 2011, presented an ectoine recovery extraction method by mixing ethanol, chloroform, and water at a ratio of 1:1:1 (v/v/v) to eliminate most of the chloroform-soluble and lipid-soluble materials from the ectoine-containing bacterial cultures. The Marinococcus sp. ECT1 cells were harvested and centrifuged prior to the addition of the solvent at the above-mentioned ratio to the cell pellets. The mixture was then centrifuged again, and the pellet was subjected to oven drying. Ethanol was used to re-dissolve the dried pellet, and ectoine was purified using activated carbon to absorb the ectoine. The ethanolic ectoine extract was then crystallized by drying at 100 °C to obtain the purified ectoine [61]. The use of large amounts of organic solvents such as ethanol and chloroform are hazardous to the environment in industrial-scale ectoine production.
3.3. Ultrafiltration with Hollow Fibers, Ethanol Precipitation, and Ion-Exchange Chromatography
Multiple combined purification steps inclusive of hollow-fiber ultrafiltration, ethanol precipitation, and ion-exchange chromatography have been applied in the purification of extracellular ectoine from H. salina DSM 5928T fermentation broth [52]. Hollow-fiber ultrafiltration was performed to remove the bacterial cells and contaminant proteins present in the broth. The resultant mixture was then concentrated by depression vaporization. Ethanol precipitation was used to remove impurities such as monosodium glutamate and NaCl. Next, the supernatant was concentrated and crystallized again with depression vaporization. Ectoine was purified with ion-exchange chromatography using highly acidic hydrochloric acid. Notably, the rates of recovery (≥86%) and purity of ectoine (97.6%) obtained after a series of multiple steps of purification and recovery were quite promising [52]. However, this purification strategy can be further improved to reduce the time-consuming steps and overall production cost of ectoine.
The yield and purity of ectoine, overall production costs, and time consumption are often the major concerns for the industrial-scale production of ectoine. Ectoine production equipment is vulnerable to corrosion due to the high-salinity ectoine fermentation media and low yields of ectoine resulting from prolonged fermentation processes are among the factors contributing to the increase in the overall production cost of ectoine. Heterologous ectoine producers can overcome the drawbacks mentioned above; however, the biological activities and morphology of the host bacteria are not well-conserved in this context [46]. Furthermore, there is an urgent need to develop a novel and cost-effective alternative for industrial-scale ectoine production, with limited enhanced integrated approaches for the production and recovery of ectoine being reported to date. The aqueous biphasic system (ABS) can serve as a potential tool to integrate the recovery, concentration, clarification, and purification of ectoine into a single-step operation. In subsequent sections, the potential of using the ABS as an integrated approach to recover ectoine with high yields is discussed.
3.4. Applications of Aqueous Biphasic System in Recovery of Ectoine from Microbial Fermentation
The aqueous biphasic system (ABS) is a well-established separation method that is widely applied in the separation and purification of various biomaterials, including low-molecular-weight bioactive compounds [62], proteins [63], metabolites [64], antibodies [65], and cell organelles. The versatile applications of the ABS provide a promising alternative for the production and recovery of ectoine from microbial sources.
The ABS is an ideal liquid–liquid extraction technology that efficiently purifies and separates labile biomaterials by partitioning the target bioproducts between two distinct aqueous phases, based on their physiochemical interaction with the phase-forming components [66]. The basis of the separation of the target bioproduct in the ABS is associated with the types, hydrophobicity, concentration, and molecular weight of the phase-forming components, as well as the surface properties, sizes, shapes, charges, and specific binding sites of the target bioproduct [67]. Therefore, different ABS parameters such as pH, temperature, and concentration of the phase-forming components are often the main factors in determining the partition behavior of the target bioproduct in an ABS. The exclusive partition of a target bioproduct in an ABS can be enhanced by the addition of certain adjuvants such as neutral salt (e.g., NaCl) [45] and ionic liquids [68]. These additives can alter the resultant physiochemical interactions between the target bioproduct and the phase-forming components, and, therefore, enhance the exclusive partition of the target bioproduct to one specific phase of the ABS. An ABS with a high-water content (70–80%) provides a mild and biocompatible environment for the recovery of biomaterials and the growth of microorganisms. A green ABS is often coupled with non-toxic, biodegradable, and non-flammable phase-forming components [69]. Furthermore, the low interfacial surface tension between the two phases facilitates the efficient mass transfer of molecules in the ABS with a minimal loss of bioactivity.
Ectoine is a zwitterionic molecule which is anticipated to exhibit different partition behavior in different types of ABS owing to the different physiochemical properties featured by different types of ABS. Bacterial milking of ectoine is often associated with high salinity, which will result in corrosion. An ABS is regarded as a promising strategy for the downstream processing of ectoine, which can prevent the use of high salt concentrations in the fermentation process and the direct recovery of ectoine by immediate removal of ectoine via extractive fermentation. Moreover, the practical implementation of an ABS for large-scale industrial application is easy and simple to operate, with great potential for continuous and automated operation with minimal overall production costs and energy inputs [70].
The immediate removal of ectoine from the fermentation broth or cell mixture upon osmotic downshock is often demanded because the ectoine will be depleted over time to supplement the microbial cell growth [45]. The recent study by Wan et al. (2018) demonstrated the one-step recovery of intracellular ectoine from an H. salina DSM 5928T cell mixture using a poly(propylene) glycol (PPG)/sulphate ABS. Crude intracellular ectoine was extracted from the fermentation broth of H. salina DSM 5928T after the osmotic downshock. The ABS was later applied to recover the ectoine from the H. salina DSM5 928T cell mixture by optimizing the concentrations of the phase-forming components, amount of loaded crude, pH of the system, and the concentration of neutral salts added into the ABS. A successful extractive ABS purification of ectoine was established in which ectoine was recovered in the salt-rich bottom phase of the PPG/sulphate ABS by exclusive partition with a partition coefficient (KE) of 150.21 ± 6.04. A total ectoine yield of 94.7% with a final purity of 87.03% and enrichment factor of 1.7 were obtained in this study, demonstrating the feasibility of the ABS to separate and recover ectoine from the H. salina cell mixture in a one-step operation [45]. In a later study, ectoine was recovered using an ionic liquids-based ABS, which further enhanced the yields and purity of the ectoine recovered with green solvent [71].
The ABS can integrate the upstream and downstream processes of ectoine into a one-step unit operation and thereby reduces the product’s inhibitory effects and the cost of the overall processes [72]. Extractive ABS fermentation is exceptionally efficient for the industrial manufacturing of ectoine without any additional investment for sophisticated equipment setup. This strategy is well suited for the recovery of extracellular ectoine sourced from the microbial fermentation of ectoine-excreting bacterial strains.
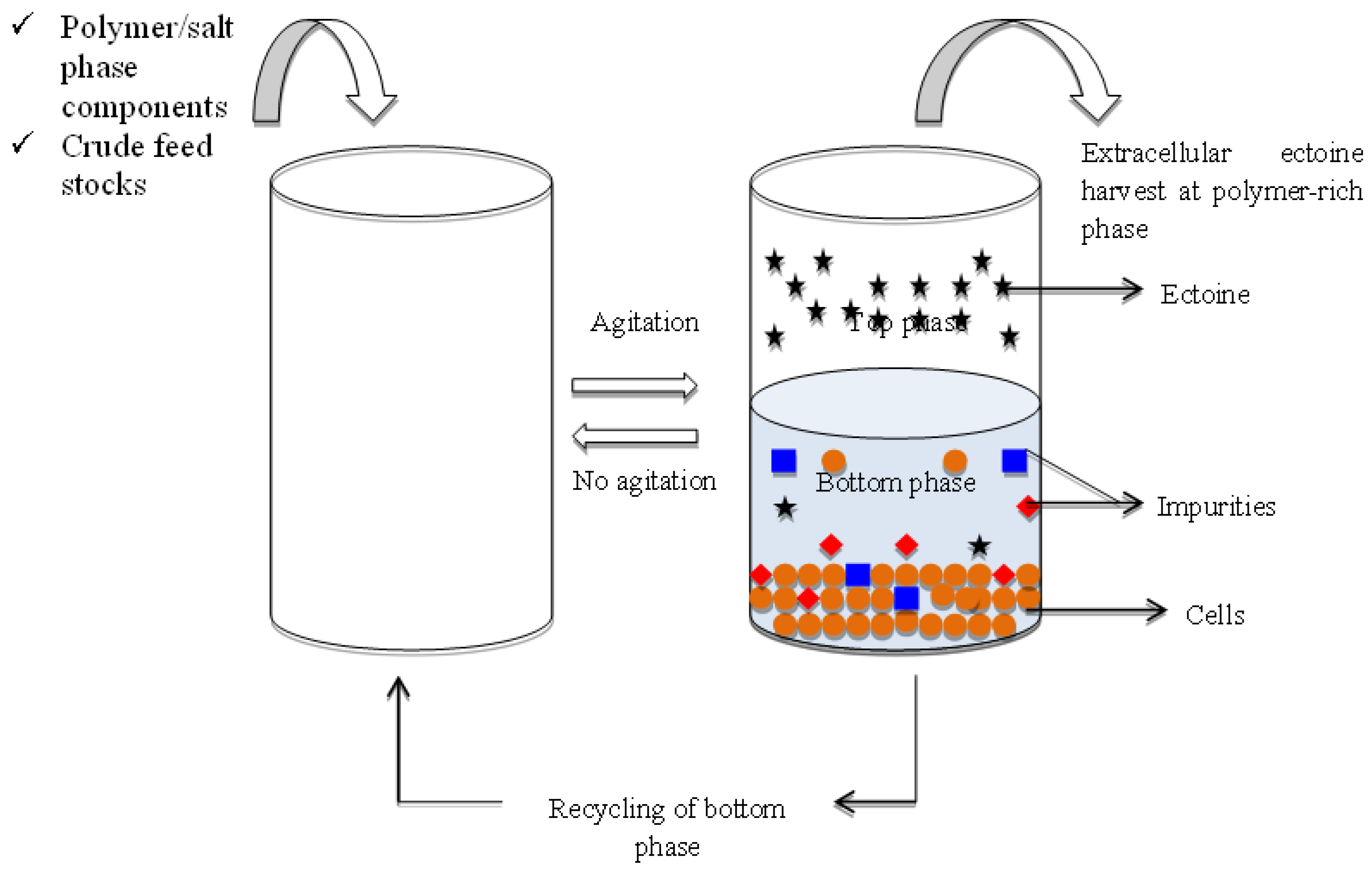
The proposed extractive ABS fermentation for ectoine is first inoculated with the desired microbial culture broth into the phase system at the optimum partition conditions. Extractive fermentation of extracellular ectoine using the ABS is achieved when the microbial cells and other impurities are partitioned to one phase, whereas ectoine was accumulated in another phase after the phase separation (Figure 1). Continuous cultivation of microbial cells and sequestration of extracellular ectoine can be attained with this integrated continuous operation by retaining the microbial cells in one phase and harvesting the ectoine from the cell-free phase. Preliminary studies on the stability of the microbial cell growth in the designed ABS are important for the development of the extractive ABS fermentation of the ectoine recovery, despite the ABS parameters for enhanced yield of ectoine.
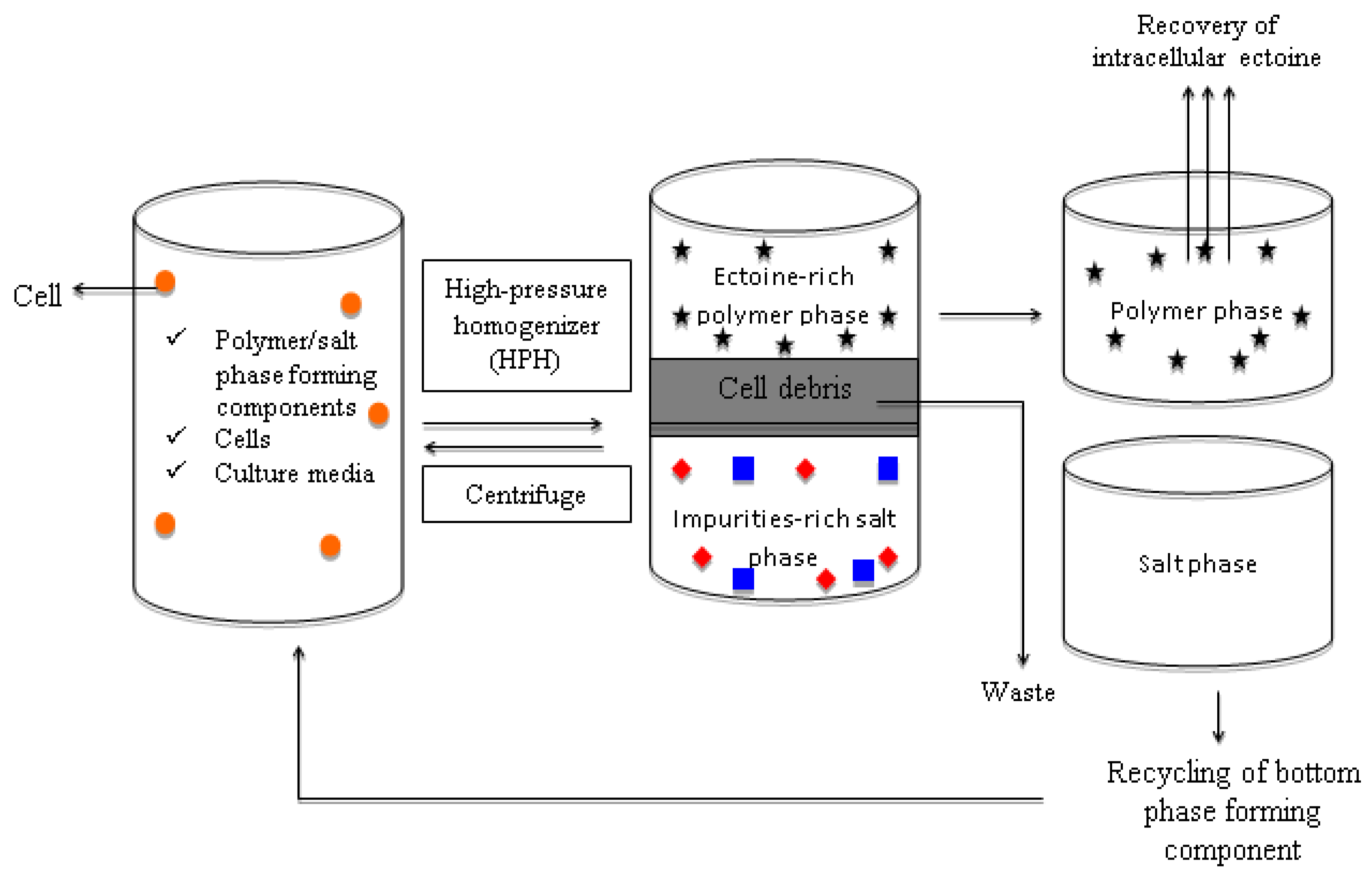
In addition, simultaneous microbial cell disruption and ectoine recovery can be achieved by integrating the ABS into the extractive disruption of intracellular ectoine. The extractive ABS disruption is feasible by incorporating the ultrasonication, homogenization, or chemical cell disruption into the ABS recovery of the intracellular ectoine. Since most of the microorganisms tend to synthesize ectoine intracellularly, extractive ABS disruption is regarded as a promising recovery strategy to integrate the cell disruption and ectoine recovery into a one-unit operation with enhanced yield and great cost efficiency. The schematic representation of the continuous, repetitive-batch extractive disruption of the intracellular ectoine-producing strain using the ABS is illustrated in Figure 2.
In the proposed ABS (Figure 2), the disrupted bacterial cells are retained in the salt-rich bottom phase of the polymer/salt ABS and the extracted ectoine was partitioned to the top phase of the ABS. However, further studies and validations are required in developing an ideal ABS for the direct recovery of ectoine via this extractive disruption approach.
4. Overall Perspective and Conclusions
The current state of the industrial production of ectoine was thoroughly discussed in this review. The major challenges of ectoine production are the use of high salt concentrations in the production process and the nature of the ectoine being synthesized intracellularly in most of the microorganisms. Therefore, the advances of genetic and metabolic engineering ease the overall production process of ectoine while enhancing the yields of ectoine by releasing the ectoine extracellularly to the fermentation broth of the microbial fermentation. Moreover, the applications of successful recovery strategies, such as the ABS, which can remove the ectoine promptly from the fermentation broth to prevent product inhibition, can enhance the overall production process while reducing the overall production cost. The research on genetically modified microbial strains and cost-efficient recovery strategies are among the main interests in the research of ectoine production.
Author Contributions
H.S.N.: Conceptualization, Funding acquisition, Supervision, Writing. P.-K.W.: Data curation, Formal Analysis, Writing. A.K.: Formal analysis, Validation writing—original draft. J.-S.C.: Conceptualization, Writing—reviewing and editing. J.C.-W.L.: Conceptualization, Supervision, Writing—Reviewing and Editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work is financially supported by National Science and Technology Council, Taiwan (Grant Number 111-2923-E-155-003-MY3 and 111-2221-E-155-007-MY3) and Research Excellence & Innovation Grant (REIG-FAS-2020-028), UCSI University, Malaysia.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors report there are no competing interest to declare.
References
- Shivanand, P.; Mugeraya, G. Halophilic bacteria and their compatible solutes–osmoregulation and potential applications. Curr. Sci. 2011, 100, 1516–1521. [Google Scholar]
- Roberts, M.F. Organic compatible solutes of halotolerant and halophilic microorganisms. Saline Syst. 2005, 1, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, C.; Jebbar, M.; Carrasco, R.; Blanco, C.; Calderón, M.I.; Iglesias-Guerra, F.; Nieto, J.J. Ectoines as compatible solutes and carbon and energy sources for the halophilic bacterium Chromohalobacter salexigens. J. Appl. Microbiol. 2006, 100, 98–107. [Google Scholar] [CrossRef]
- Galinski, E.A.; Pfeiffer, H.P.; Trüper, H.G. 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidinecarboxylic acid: A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. Eur. J. Biochem. 1985, 149, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A. Osmoadaptation in bacteria. Adv. Microb. Physiol. 1995, 37, 273–328. [Google Scholar]
- Louis, P.; Galinski, E.A. Characterization of genes for the biosynthesis of the compatible solute ectoine from Marinococcus halophilus and osmoregulated expression in Escherichia coli. Microbiology 1997, 143, 1141–1149. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Huang, X.; Gu, L.; Jiang, J.; Li, S. Cloning of the ectoine biosynthesis gene ectABC from Halomonas sp. BYS-1 and salt stressed expression in Escherichia coli. Wei Sheng Wu Xue Bao Acta Microbiol. Sin. 2006, 46, 28–32. [Google Scholar]
- Becker, J.; Schafer, R.; Kohlstedt, M.; Harder, B.J.; Borchert, N.S.; Stoveken, N.; Bremer, E.; Wittmann, C. Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine. Microb. Cell Factories 2013, 12, 110. [Google Scholar] [CrossRef] [Green Version]
- Bestvater, T.; Louis, P.; Galinski, E.A. Heterologous ectoine production in Escherichia coli: By-passing the metabolic bottle-neck. Saline Syst. 2008, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Bursy, J.; Pierik, A.J.; Pica, N.; Bremer, E. Osmotically induced synthesis of the compatible solute hydroxyectoine is mediated by an evolutionarily conserved ectoine hydroxylase. J. Biol. Chem. 2007, 282, 31147–31155. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. Bioenergetic aspects of halophilism. Microbiol. Mol. Biol. Rev. 1999, 63, 334–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor, J.M.; Salvador, M.; Argandoña, M.; Bernal, V.; Reina-Bueno, M.; Csonka, L.N.; Iborra, J.L.; Vargas, C.; Nieto, J.J.; Cánovas, M.J. Ectoines in cell stress protection: Uses and biotechnological production. Biotechnol. Adv. 2010, 28, 782–801. [Google Scholar] [CrossRef] [PubMed]
- Lippert, K.; Galinski, E.A. Enzyme stabilization be ectoine-type compatible solutes: Protection against heating, freezing and drying. Appl. Microbiol. Biotechnol. 1992, 37, 61–65. [Google Scholar] [CrossRef]
- Kolp, S.; Pietsch, M.; Galinski, E.A.; Gütschow, M.J. Compatible solutes as protectants for zymogens against proteolysis. Biochim. Biophys. Acta BBA Proteins Proteom. 2006, 1764, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Göller, K.; Galinski, E.A. Protection of a model enzyme (lactate dehydrogenase) against heat, urea and freeze-thaw treatment by compatible solute additives. J. Mol. Catal. B Enzym. 1999, 7, 37–45. [Google Scholar] [CrossRef]
- Khmelenina, V.N.; Mustakhimov, I.I.; Reshetnikov, A.S.; Kalyuzhnaya, M.G.; Trotsenko, Y.A. Genetic and biochemical aspects of ectoine biosynthesis in moderately halophilic and halotolerant methylotrophic bacteria. J. Agric. Biol. Sci. 2010, 5, 446–458. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Martínez, A.A.; Marcos-Rodrigo, E.; Bordel, S.; Marín, D.; Herrero-Lobo, R.; García-Encina, P.A.; Muñoz, R. Elucidating the key environmental parameters during the production of ectoines from biogas by mixed methanotrophic consortia. J. Environ. Manag. 2021, 298, 113462. [Google Scholar] [CrossRef] [PubMed]
- Cantera, S.; Phandanouvong-Lozano, V.; Pascual, C.; García-Encina, P.A.; Lebrero, R.; Hay, A.; Muñoz, R. A systematic comparison of ectoine production from upgraded biogas using Methylomicrobium alcaliphilum and a mixed haloalkaliphilic consortium. Waste Manag. 2020, 102, 773–781. [Google Scholar] [CrossRef]
- Widderich, N.; Czech, L.; Elling, F.J.; Konneke, M.; Stoveken, N.; Pittelkow, M.; Riclea, R.; Dickschat, J.S.; Heider, J.; Bremer, E. Strangers in the archaeal world: Osmostress-responsive biosynthesis of ectoine and hydroxyectoine by the marine thaumarchaeon Nitrosopumilus maritimus. Environ. Microbiol. 2016, 18, 1227–1248. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, L.; Li, X.; Liu, S.; Li, D. Poly-β-hydroxybutyrate/ectoine co-production by ectoine-excreting strain Halomonas salina. Process Biochem. 2014, 49, 33–37. [Google Scholar] [CrossRef]
- Van-Thuoc, D.; Guzmán, H.; Quillaguamán, J.; Hatti-Kaul, R. High productivity of ectoines by Halomonas boliviensis using a combined two-step fed-batch culture and milking process. J. Biotechnol. 2010, 147, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Schwibbert, K.; Marin-Sanguino, A.; Bagyan, I.; Heidrich, G.; Lentzen, G.; Seitz, H.; Rampp, M.; Schuster, S.C.; Klenk, H.P.; Pfeiffer, F. A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581T. Environ. Microbiol. 2011, 13, 1973–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widderich, N.; Hoppner, A.; Pittelkow, M.; Heider, J.; Smits, S.H.; Bremer, E. Biochemical properties of ectoine hydroxylases from extremophiles and their wider taxonomic distribution among microorganisms. PLoS ONE 2014, 9, e93809. [Google Scholar] [CrossRef]
- Fenizia, S.; Thume, K.; Wirgenings, M.; Pohnert, G. Ectoine from Bacterial and Algal Origin is a Compatible Solute in Microalgae. Mar. Drugs 2020, 18, 42. [Google Scholar] [CrossRef] [Green Version]
- Pérez, V.; Moltó, J.L.; Lebrero, R.; Muñoz, R. Ectoine production from biogas: A sensitivity analysis. Effect of local commodity prices, economy of scale, market trends and biotechnological limitations. J. Clean. Prod. 2022, 369, 133440. [Google Scholar] [CrossRef]
- Rodero, M.D.R.; Herrero-Lobo, R.; Pérez, V.; Muñoz, R. Influence of operational conditions on the performance of biogas bioconversion into ectoines in pilot bubble column bioreactors. Bioresour. Technol. 2022, 358, 127398. [Google Scholar] [CrossRef]
- Kang, J.Y.; Lee, B.; Kim, J.A.; Kim, M.-S.; Kim, C.H. Identification and characterization of an ectoine biosynthesis gene cluster from Aestuariispira ectoiniformans sp. nov., isolated from seawater. Microbiol. Res. 2022, 254, 126898. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L. Ectoine improves yield of biodiesel catalyzed by immobilized lipase. J. Mol. Catal. B Enzym. 2010, 62, 90–95. [Google Scholar] [CrossRef]
- Bethlehem, L.; van Echten-Deckert, G. Ectoines as novel anti-inflammatory and tissue protective lead compounds with special focus on inflammatory bowel disease and lung inflammation. Pharmacol. Res. 2021, 164, 105389. [Google Scholar] [CrossRef]
- Buenger, J.; Driller, H. Ectoin: An effective natural substance to prevent UVA-induced premature photoaging. Ski. Pharmacol. Physiol. 2004, 17, 232–237. [Google Scholar] [CrossRef]
- Graf, R.; Anzali, S.; Buenger, J.; Pfluecker, F.; Driller, H. The multifunctional role of ectoine as a natural cell protectant. Clin. Dermatol. 2008, 26, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Marini, A.; Reinelt, K.; Krutmann, J.; Bilstein, A. Ectoine-containing cream in the treatment of mild to moderate atopic dermatitis: A randomised, comparator-controlled, intra-individual double-blind, multi-center trial. Ski. Pharmacol. Physiol. 2014, 27, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Bow, J.R.; Sonoki, Y.; Uchiyama, M.; Dauskardt, R.H. Ectoine disperses keratin and alters hydration kinetics in stratum corneum. Biochem. Biophys. Rep. 2021, 28, 101134. [Google Scholar] [CrossRef] [PubMed]
- Sydlik, U.; Gallitz, I.; Albrecht, C.; Abel, J.; Krutmann, J.; Unfried, K. The compatible solute ectoine protects against nanoparticle-induced neutrophilic lung inflammation. Am. J. Respir. Crit. Care Med. 2009, 180, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wedeking, A.; Buttner, R.; Kalff, J.C.; Tolba, R.H.; van Echten-Deckert, G. A natural tetrahydropyrimidine protects small bowel from cold ischemia and subsequent warm in vitro reperfusion injury. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2009, 76, 212–220. [Google Scholar] [CrossRef]
- Kanapathipillai, M.; Lentzen, G.; Sierks, M.; Park, C.B. Ectoine and hydroxyectoine inhibit aggregation and neurotoxicity of Alzheimer’s beta-amyloid. FEBS Lett. 2005, 579, 4775–4780. [Google Scholar] [CrossRef] [Green Version]
- Harishchandra, R.K.; Sachan, A.K.; Kerth, A.; Lentzen, G.; Neuhaus, T.; Galla, H.J. Compatible solutes: Ectoine and hydroxyectoine improve functional nanostructures in artificial lung surfactants. Biochim. Biophys. Acta 2011, 1808, 2830–2840. [Google Scholar] [CrossRef] [Green Version]
- Bownik, A.; Stepniewska, Z. Protective effects of bacterial osmoprotectant ectoine on bovine erythrocytes subjected to staphylococcal alpha-haemolysin. Toxicon Off. J. Int. Soc. Toxinology 2015, 99, 130–135. [Google Scholar] [CrossRef]
- Unfried, K.; Kroker, M.; Autengruber, A.; Gotić, M.; Sydlik, U. The Compatible Solute Ectoine Reduces the Exacerbating Effect of Environmental Model Particles on the Immune Response of the Airways. J. Allergy 2014, 2014, 7. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.-L.; Lin, Y.-M.; Mohamed, M.S.; Chen, J.-H. Inhibitory effect of ectoine on melanogenesis in B16-F0 and A2058 melanoma cell lines. Biochem. Eng. J. 2013, 78, 163–169. [Google Scholar] [CrossRef]
- Anzali, S.; Von Heydebreck, A.; Herget, T. Elucidation of the anti-aging effects of ectoine using cdna microarray analysis and signaling pathway evaluation. Int. J. Cosmet. Sci. 2010, 32, 319. [Google Scholar] [CrossRef]
- Werkhäuser, N.; Bilstein, A.; Sonnemann, U. Treatment of Allergic Rhinitis with Ectoine Containing Nasal Spray and Eye Drops in Comparison with Azelastine Containing Nasal Spray and Eye Drops or with Cromoglycic Acid Containing Nasal Spray. J. Allergy 2014, 2014, 13. [Google Scholar] [CrossRef] [Green Version]
- Sonnemann, U.; Scherner, O.; Werkhäuser, N. Treatment of Rhinitis Sicca Anterior with Ectoine Containing Nasal Spray. J. Allergy 2014, 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Eichel, A.; Wittig, J.; Shah-Hosseini, K.; Mosges, R. A prospective, controlled study of SNS01 (ectoine nasal spray) compared to BNO-101 (phytotherapeutic dragees) in patients with acute rhinosinusitis. Curr. Med. Res. Opin. 2013, 29, 739–746. [Google Scholar] [CrossRef]
- Wan, P.K.; Lan, J.C.-W.; Chen, P.-W.; Tan, J.S.; Ng, H.S. Recovery of intracellular ectoine from Halomonas salina cells with poly(propylene) glycol/salt aqueous biphasic system. J. Taiwan Inst. Chem. Eng. 2018, 82, 28–32. [Google Scholar] [CrossRef]
- Zhang, L.H.; Lang, Y.J.; Nagata, S. Efficient production of ectoine using ectoine-excreting strain. Extrem. Life Under Extrem. Cond. 2009, 13, 717–724. [Google Scholar] [CrossRef]
- Chen, P.-W.; Cui, Z.-Y.; Ng, H.S.; Chi-Wei Lan, J. Exploring the additive bio-agent impacts upon ectoine production by Halomonas salina DSM5928T using corn steep liquor and soybean hydrolysate as nutrient supplement. J. Biosci. Bioeng. 2020, 130, 195–199. [Google Scholar] [CrossRef]
- Onraedt, A.E.; Walcarius, B.A.; Soetaert, W.K.; Vandamme, E.J. Optimization of ectoine synthesis through fed-batch fermentation of Brevibacterium epidermis. Biotechnol. Prog. 2005, 21, 1206–1212. [Google Scholar] [CrossRef]
- Fallet, C.; Rohe, P.; Franco-Lara, E. Process optimization of the integrated synthesis and secretion of ectoine and hydroxyectoine under hyper/hypo-osmotic stress. Biotechnol. Bioeng. 2010, 107, 124–133. [Google Scholar] [CrossRef]
- Sauer, T.; Galinski, E.A. Bacterial milking: A novel bioprocess for production of compatible solutes. Biotechnol. Bioeng. 1998, 57, 306–313. [Google Scholar] [CrossRef]
- Guzman, H.; Van-Thuoc, D.; Martin, J.; Hatti-Kaul, R.; Quillaguaman, J. A process for the production of ectoine and poly(3-hydroxybutyrate) by Halomonas boliviensis. Appl. Microbiol. Biotechnol. 2009, 84, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.J.; Bai, L.; Ren, Y.N.; Zhang, L.H.; Nagata, S. Production of ectoine through a combined process that uses both growing and resting cells of Halomonas salina DSM 5928T. Extrem. Life Under Extrem. Cond. 2011, 15, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.; Song, Y.; You, J.; Zhang, X.; Xu, M.; Rao, Z. High-yield ectoine production in engineered Corynebacterium glutamicum by fine metabolic regulation via plug-in repressor library. Bioresour. Technol. 2022, 362, 127802. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, J.; Wang, Y.; Du, G.; Kang, Z. Engineering Escherichia coli for high-yield production of ectoine. Green Chem. Eng. 2021, in press. [Google Scholar] [CrossRef]
- Anbu Rajan, L.; Joseph, T.C.; Thampuran, N.; James, R.; Ashok Kumar, K.; Viswanathan, C.; Bansal, K.C. Cloning and heterologous expression of ectoine biosynthesis genes from Bacillus halodurans in Escherichia coli. Biotechnol. Lett. 2008, 30, 1403–1407. [Google Scholar] [CrossRef]
- Schubert, T.; Maskow, T.; Benndorf, D.; Harms, H.; Breuer, U. Continuous synthesis and excretion of the compatible solute ectoine by a transgenic, nonhalophilic bacterium. Appl. Environ. Microbiol. 2007, 73, 3343–3347. [Google Scholar] [CrossRef] [Green Version]
- He, Y.-Z.; Gong, J.; Yu, H.-Y.; Tao, Y.; Zhang, S.; Dong, Z.-Y. High production of ectoine from aspartate and glycerol by use of whole-cell biocatalysis in recombinant Escherichia coli. Microb. Cell Factories 2015, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Tanimura, K.; Nakayama, H.; Tanaka, T.; Kondo, A. Ectoine production from lignocellulosic biomass-derived sugars by engineered Halomonas elongata. Bioresour. Technol. 2013, 142, 523–529. [Google Scholar] [CrossRef]
- Tanimura, K.; Matsumoto, T.; Nakayama, H.; Tanaka, T.; Kondo, A. Improvement of ectoine productivity by using sugar transporter-overexpressing Halomonas elongata. Enzym. Microb. Technol. 2016, 89, 63–68. [Google Scholar] [CrossRef]
- Ning, Y.; Wu, X.; Zhang, C.; Xu, Q.; Chen, N.; Xie, X. Pathway construction and metabolic engineering for fermentative production of ectoine in Escherichia coli. Metab. Eng. 2016, 36, 10–18. [Google Scholar] [CrossRef]
- Wei, Y.H.; Yuan, F.W.; Chen, W.C.; Chen, S.Y. Production and characterization of ectoine by Marinococcus sp. ECT1 isolated from a high-salinity environment. J. Biosci. Bioeng. 2011, 111, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Simental-Martínez, J.; Montalvo-Hernández, B.; Rito-Palomares, M.; Benavides, J. Application of Aqueous Two-Phase Systems for the Recovery of Bioactive Low-Molecular Weight Compounds. Sep. Sci. Technol. 2014, 49, 1872–1882. [Google Scholar] [CrossRef]
- Ng, H.S.; Wang, C.-C.; Tan, J.S.; Lan, J.C.-W. Primary recovery of recombinant human serum albumin from transgenic Oryza sativa with a single-step aqueous biphasic system. J. Taiwan Inst. Chem. Eng. 2018, 84, 60–66. [Google Scholar] [CrossRef]
- Gu, B.H.; Zheng, P.; Yan, Q.; Liu, W. Aqueous two-phase system: An alternative process for recovery of succinic acid from fermentation broth. Sep. Purif. Technol. 2014, 138, 47–54. [Google Scholar] [CrossRef]
- Campos-Pinto, I.; Espitia-Saloma, E.; Rosa, S.A.S.L.; Rito-Palomares, M.; Aguilar, O.; Arévalo-Rodríguez, M.; Aires-Barros, M.R.; Azevedo, A.M. Integration of cell harvest with affinity-enhanced purification of monoclonal antibodies using aqueous two-phase systems with a dual tag ligand. Sep. Purif. Technol. 2017, 173, 129–134. [Google Scholar] [CrossRef]
- Tan, Z.; Li, F.; Xu, X. Isolation and purification of aloe anthraquinones based on an ionic liquid/salt aqueous two-phase system. Sep. Purif. Technol. 2012, 98, 150–157. [Google Scholar] [CrossRef]
- Asenjo, J.A.; Andrews, B.A. Aqueous two-phase systems for protein separation: A perspective. J. Chromatogr. A 2011, 1218, 8826–8835. [Google Scholar] [CrossRef]
- Aziz, N.F.H.A.; Abbasiliasi, S.; Ng, H.S.; Phapugrangkul, P.; Bakar, M.H.A.; Tam, Y.J.; Tan, J.S. Purification of β-mannanase derived from Bacillus subtilis ATCC 11774 using ionic liquid as adjuvant in aqueous two-phase system. J. Chromatogr. B 2017, 1055, 104–112. [Google Scholar] [CrossRef]
- Ruiz, C.S.; van den Berg, C.; Wijffels, R.H.; Eppink, M.H.M. Rubisco separation using biocompatible aqueous two-phase systems. Sep. Purif. Technol. 2018, 196, 254–261. [Google Scholar] [CrossRef]
- Ng, H.S.; Ooi, C.W.; Mokhtar, M.N.; Show, P.L.; Ariff, A.; Tan, J.S.; Ng, E.-P.; Ling, T.C. Extractive bioconversion of cyclodextrins by Bacillus cereus cyclodextrin glycosyltransferase in aqueous two-phase system. Bioresour. Technol. 2013, 142, 723–726. [Google Scholar] [CrossRef] [Green Version]
- Ng, H.S.; Wan, P.K.; Ng, T.-C.; Lan, J.C.-W. Primary purification of intracellular Halomonas salina ectoine using ionic liquids-based aqueous biphasic system. J. Biosci. Bioeng. 2020, 130, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Ooi, C.W.; Hii, S.L.; Kamal, S.M.M.; Ariff, A.; Ling, T.C. Extractive fermentation using aqueous two-phase systems for integrated production and purification of extracellular lipase derived from Burkholderia pseudomallei. Process Biochem. 2011, 46, 68–73. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram on the proposed extractive fermentation of microbial ectoine with ABS. A process scheme was illustrated for continuous repetitive batch of extractive ABS fermentation for recovery of ectoine. The microbial cells is represented by “🟠”, the ectoine is denoted by “🟊” and the impurities are denoted with “▄” and “◆”.
Figure 1.
Schematic diagram on the proposed extractive fermentation of microbial ectoine with ABS. A process scheme was illustrated for continuous repetitive batch of extractive ABS fermentation for recovery of ectoine. The microbial cells is represented by “🟠”, the ectoine is denoted by “🟊” and the impurities are denoted with “▄” and “◆”.

Figure 2.
Schematic diagram on the proposed extractive disruption of microbial ectoine with ABS. A process scheme was illustrated for continuous repetitive batch of extractive ABS disruption for recovery of ectoine. The microbial cells is represented by “🟠”, the ectoine is denoted by “🟊” and the impurities are denoted with “▄” and “◆”.
Figure 2.
Schematic diagram on the proposed extractive disruption of microbial ectoine with ABS. A process scheme was illustrated for continuous repetitive batch of extractive ABS disruption for recovery of ectoine. The microbial cells is represented by “🟠”, the ectoine is denoted by “🟊” and the impurities are denoted with “▄” and “◆”.


{kind=link}
{kind=link}
Table 1.
Summary of several potential microbial ectoine and hydroxyectoine producers with ectoine biosynthetic gene cluster. “√” indicates whether the ectD is located within/outside of the ect gene cluster (adapted from [23]).
Table 1.
Summary of several potential microbial ectoine and hydroxyectoine producers with ectoine biosynthetic gene cluster. “√” indicates whether the ectD is located within/outside of the ect gene cluster (adapted from [23]).
| Phylum | Microorganisms | ectD is Outside of ect Gene Cluster | ectD is within the ect Gene Cluster |
|---|---|---|---|
| Actinobacteria | Brachybacterium faecium DSM 4810 Brevibacterium album DSM 18261 | √ √ | |
| Gordonia terrae NBRC 100016 | √ | ||
| Kytococcus sedentarius DSM 20547 | √ | ||
| Streptomyces coelicolor A3 (2) | √ | ||
| Firmicutes | Bacillus halodurans DSM 497T Bacillus pseudofirmus OF4 | √ √ | |
| Halobacillus halophilus DSMZ 2266T | √ | ||
| Marinococcus halotolerans DSM 16375 Marinococcus sp. M52 Virgibacillus salexigens DSM 11438 | √ √ | √ | |
| Proteobacteria | Achromobacter xylosidans A8 | √ | |
| Acidiphilum crytum JF-5 | √ | ||
| Cellvibrio japonicus Ueda107 Chromohalobacter salexigens DSM 3043 | √ | √ | |
| Halomonas boliviensis Halomonas elongata DSM 2581 | √ √ | ||
| Nitrosococcus oceani ATCC 19707 Pseudomonas stutzeri A1501 Roseobacter sp. MED193 Vibrio cholerae O395 | √ √ | √ √ | |
| Archaea | Nitrosopumilus sp. AR2 Candidatus | √ |
Table 2.
Several significant biomedical and skin-protective effects of ectoine.
| Protective Effects against/on: | Treatment Target | Ref. |
|---|---|---|
| Nanoparticle | Neutrophilic lung inflammation | [34] |
| Small bowel | Cold ischemia, reperfusion injury | [35] |
| Aggregation and neurotoxicity of Alzheimer’s β-amyloid | Alzheimer’s disease | [36] |
| Improvement on functional nanostructures | Artificial lung surfactants | [37] |
| Staphylococcal α-hemolysin | Bovine erythrocytes | [38] |
| Exacerbating effect of environmental model particles | Immune response of the airways | [39] |
| Melanogens | Melanogenesis | [40] |
| Ultraviolet A, anti-aging | Skin protection | [23,41] |
| Mild to moderate atopic dermatitis | Allergy | [25] |
| Transepidermal water loss | Skin dehydration | [24] |
| In vivo protection | Allergic rhinitis, Acute rhinosinusitis, rhinitis sicca anterior | [42,43] |
Table 3.
Large-scale production of ectoine by different microbial sources.
| Microorganisms | Fermentation Setting | Intracellular /Extracellular Ectoine | Carbon Source | Concentration of NaCl (M) | Maximum Production Yield (mg/gcdw) | Maximum Productivity (g/L/day) | Ref. |
|---|---|---|---|---|---|---|---|
| Brevibacterium epidermis DSM 20659 | Fed-batch | Intracellular | Sodium glutamate | 1.0 | 160 | 2 | [48] |
| Chromohalobacter salexigens | Continuous cell retention | Intracellular | Glucose | 1.8 | 540 | 32.5 | [49] |
| Halomonas elongata DSM 142T | Batch | Intracellular | Glucose, ammonium chloride | 2.6 | 155 | 5.3 | [50] |
| Halomonas boliviensis DSM 15516 | Two-step fed-batch | Intracellular | Glucose and sodium glutamate (c) | 0.17 (a) 2.13 (b) | 170 | 3.4 | [51] |
| Halomonas salina DSM 5928 | Batch | Both | Sodium glutamate | 0.5 | 245.9 | 7.9 | [46] |
| Halomonas salina DSM 5928T | Combined Batch | Both | Sodium glutamate | 0.5 | 140 | 7.75 | [52] |
| Halomonas boliviensis LC1T | Two-step fed-batch, batch | Intracellular | Glucose and sodium glutamate (c) | 0.77 (a) 2.55 (b) | 154 | 9.1 | [21] |
(a) Salt concentration for first fed-batch; (b) salt concentration for second fed-batch; (c) glutamate only fed for the first fed-batch.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ng, H.S.; Wan, P.-K.; Kondo, A.; Chang, J.-S.; Lan, J.C.-W. Production and Recovery of Ectoine: A Review of Current State and Future Prospects. Processes 2023, 11, 339. https://0-doi-org.brum.beds.ac.uk/10.3390/pr11020339
AMA Style
Ng HS, Wan P-K, Kondo A, Chang J-S, Lan JC-W. Production and Recovery of Ectoine: A Review of Current State and Future Prospects. Processes. 2023; 11(2):339. https://0-doi-org.brum.beds.ac.uk/10.3390/pr11020339
Chicago/Turabian StyleNg, Hui Suan, Pik-Ki Wan, Akihiko Kondo, Jo-Shu Chang, and John Chi-Wei Lan. 2023. "Production and Recovery of Ectoine: A Review of Current State and Future Prospects" Processes 11, no. 2: 339. https://0-doi-org.brum.beds.ac.uk/10.3390/pr11020339
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.