Effects of Mercury II on Cupriavidus metallidurans Strain MSR33 during Mercury Bioremediation under Aerobic and Anaerobic Conditions





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
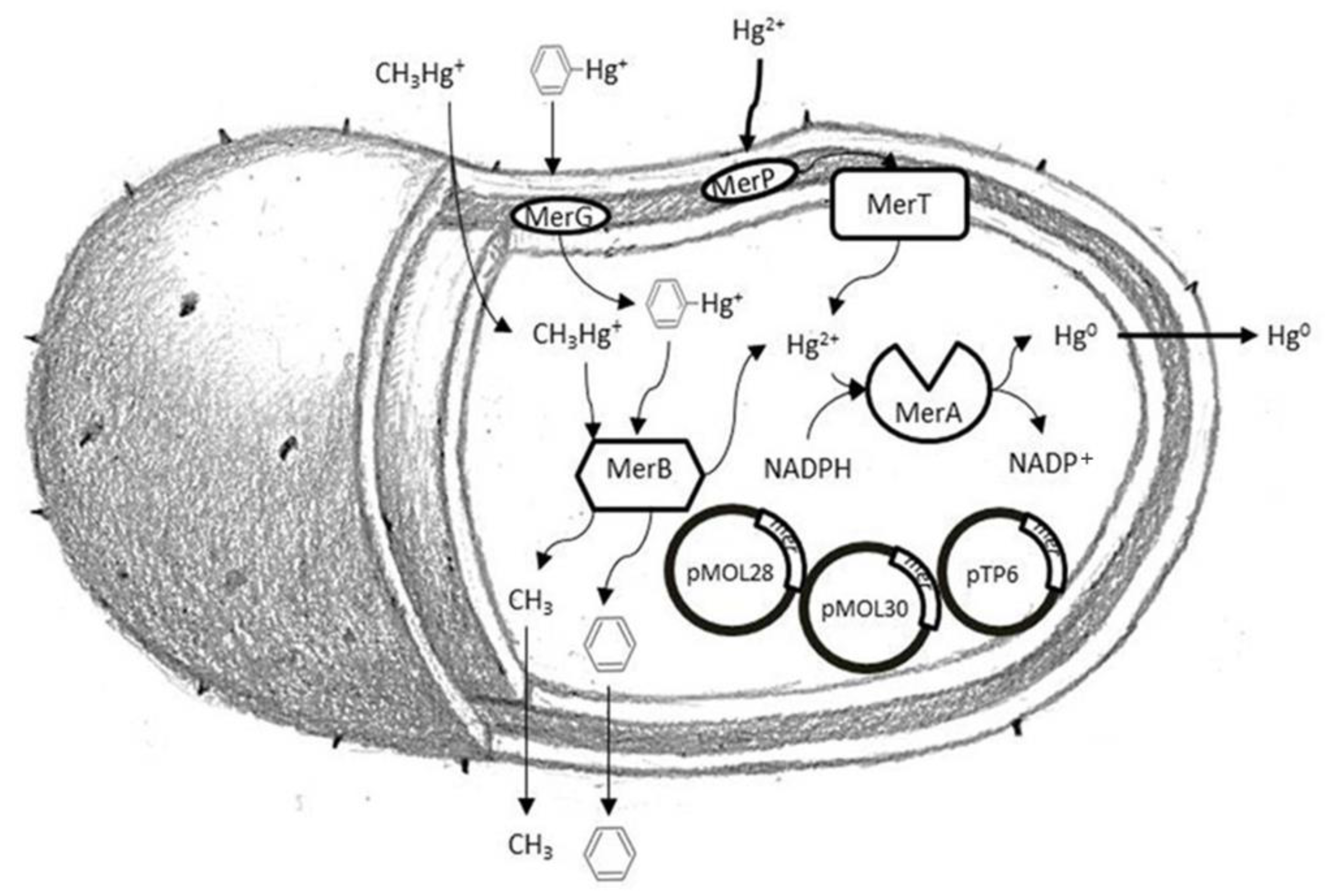
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Culture Medium
2.3. Biomass Determination
2.4. Succinate Quantification
2.5. Determination of Dissolved Oxygen Concentration
2.6. Mercury Determination
2.7. Kinetics of C. metallidurans MSR33 Growth
2.8. Effects of Mercury on Growth and Oxygen Uptake Rate of C. metallidurans Strain MSR33
2.9. Effects of Oxygen Availability on Mercury Reduction
3. Results
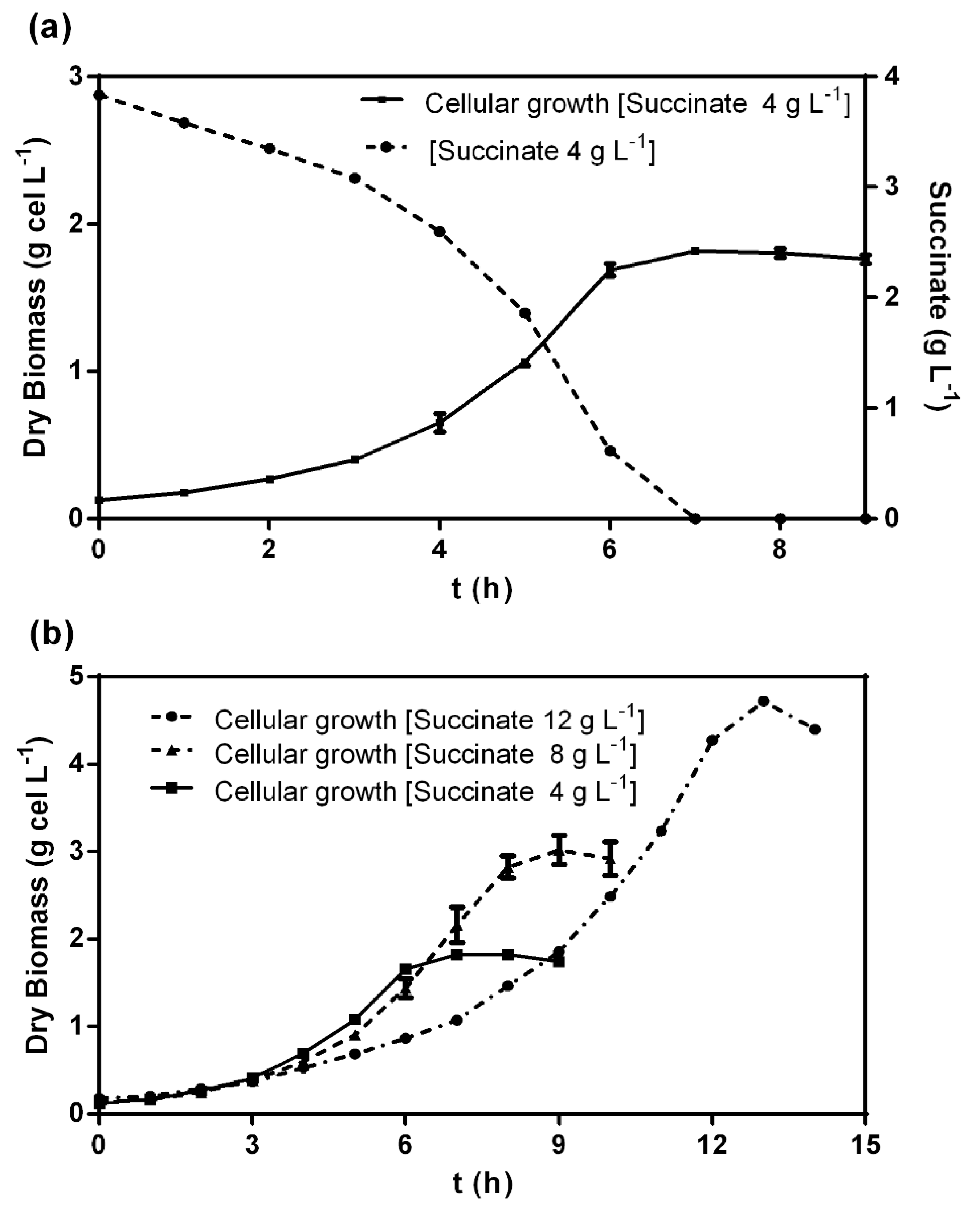
3.1. Culture Medium Design, Kinetics, and Operational Characterization of C. metallidurans MSR33 Growth
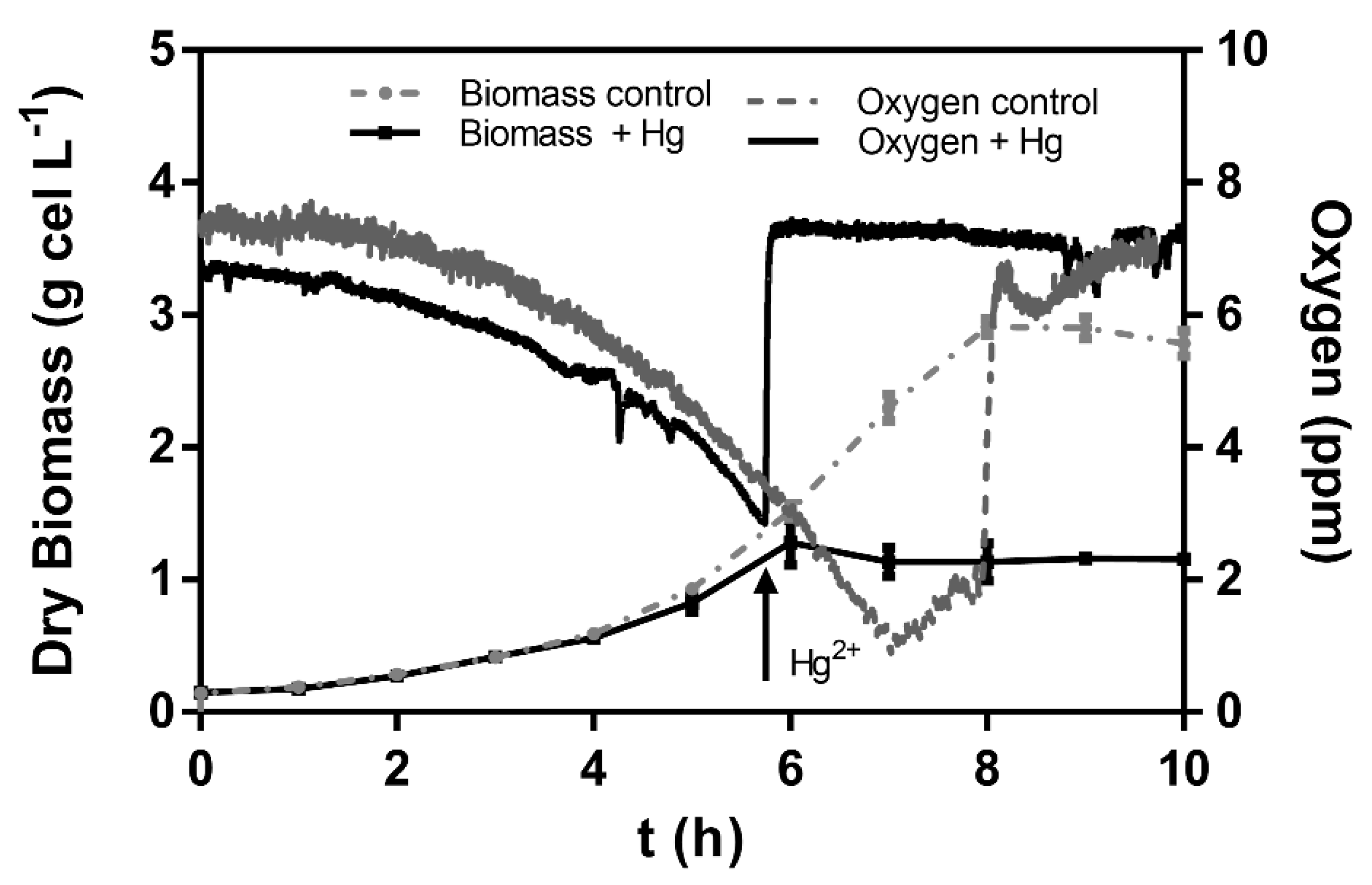
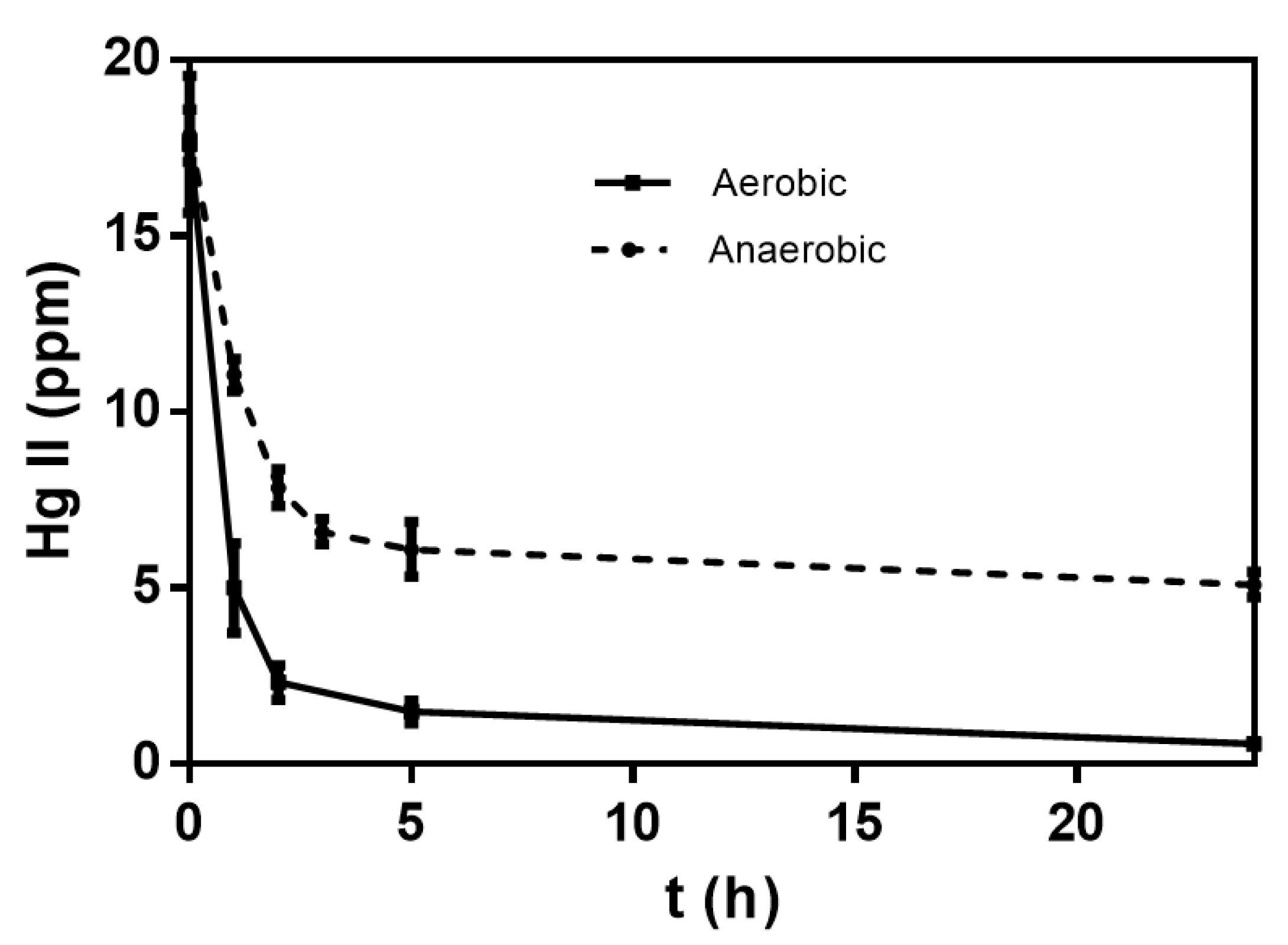
3.2. Effect of Mercury (II) on Bioremediation in Liquid Medium by C. metallidurans Strain MSR33
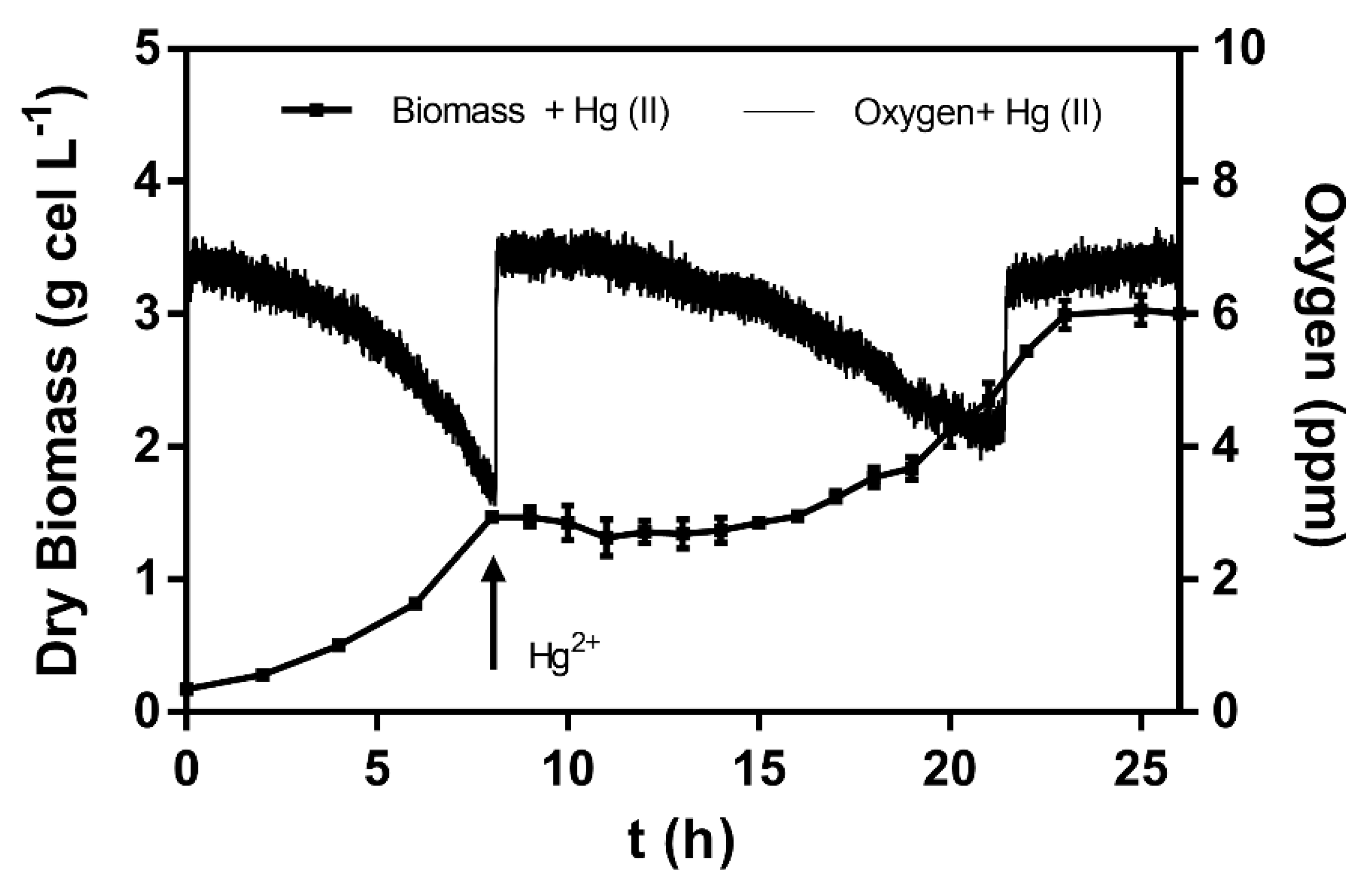
3.3. Effect of Oxygen Availability on Mercury (II) Reduction
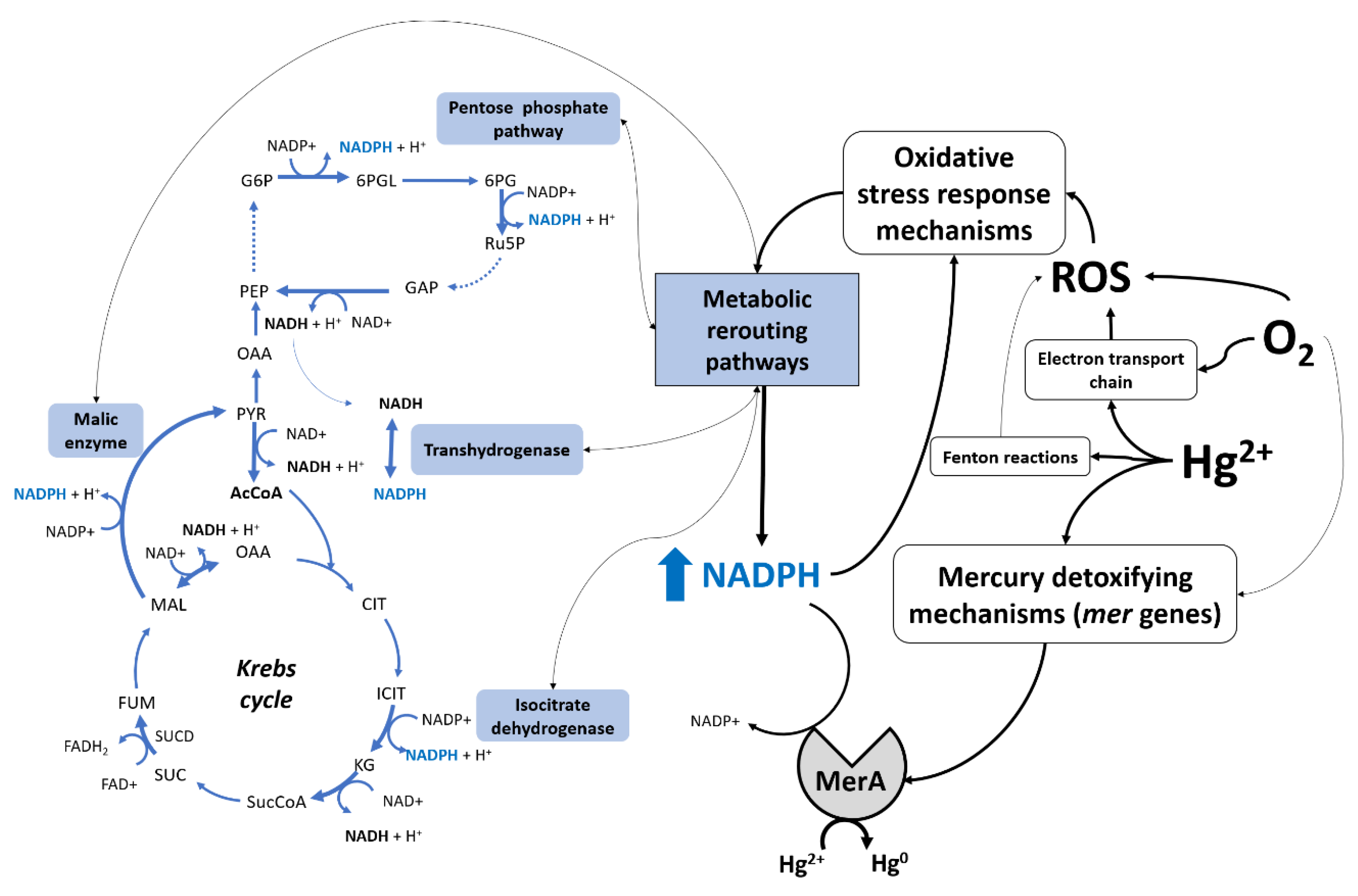
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nieboer, E.; Richardson, D.H. The replacement of the nondescript term ‘heavy metals’ by a biologically and chemically significant classification of metal ions. Environ. Pollut. Ser. B Chem. Phys. 1980, 1, 3–26. [Google Scholar] [CrossRef]
- Tagliafierro, L.; Officioso, A.; Sorbo, S.; Basile, A.; Manna, C. The protective role of olive oil hydroxytyrosol against oxidative alterations induced by mercury in human erythrocytes. Food Chem. Toxicol. 2015, 82, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Y.; Wang, F.; Luo, Z.; Guo, S.; Strähle, U. Toxicity of mercury: Molecular evidence. Chemosphere 2020, 245, 125586. [Google Scholar] [CrossRef] [PubMed]
- Patra, M.; Sharma, A. Mercury toxicity in plants. Bot. Rev. 2000, 66, 379–422. [Google Scholar] [CrossRef]
- Streets, D.G.; Horowitz, H.M.; Jacob, D.J.; Lu, Z.; Levin, L.; Ter Schure, A.F.H.; Sunderland, E.M. Total mercury released to the environment by human activities. Environ. Sci. Technol. 2017, 51, 5969–5977. [Google Scholar] [CrossRef] [PubMed]
- Spiller, H.A. Rethinking mercury: The role of selenium in the pathophysiology of mercury toxicity. Clin. Toxicol. 2017, 56, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Montero-Silva, F.; Durán, N.; Seeger, M. Synthesis of extracellular gold nanoparticles using Cupriavidus metallidurans CH34 cells. IET Nanobiotechnol. 2018, 12, 40–46. [Google Scholar] [CrossRef]
- Xu, F.F.; Imlay, J.A. Silver(I), mercury (II), cadmium (II), and zinc (II) target exposed enzymic iron-sulfur clusters when they toxify Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 3614–3621. [Google Scholar] [CrossRef] [Green Version]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Rojas, L.A.; Yañez, C.; González, M.; Lobos, S.; Smalla, K.; Seeger, M. Characterization of the metabolically modified heavy metal-resistant Cupriavidus metallidurans strain MSR33 generated for mercury bioremediation. PLoS ONE 2011, 6, e17555. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.; Edwards, G.C.; Gustin, M.S.; Care, A.; Miller, M.B.; Sunna, A. An innovative approach to bioremediation of mercury contaminated soils from industrial mining operations. Chemosphere 2017, 184, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.; Morgante, V.; Ávila, M.; Villalobos, P.; Millares, P.; González, M.; Seeger, M. Novel s-triazine-degrading bacteria isolated from agricultural soils of central Chile for herbicide bioremediation. Electron. J. Biotechnol. 2008, 11, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Seeger, M.; Hernández, M.; Mendez, V.; Ponce, B.; Córdova, M.; González, M. Bacterial degradation and bioremediation of chlorinated herbicides and biphenyls. J. Soil Sci. Plant Nutr. 2010, 10, 320–332. [Google Scholar] [CrossRef]
- Altimira, F.; Yáñez, C.; Bravo, G.; González, M.; Rojas, L.A.; Seeger, M. Characterization of copper-resistant bacteria and bacterial communities from copper-polluted agricultural soils of central Chile. BMC Microbiol. 2012, 12, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Silva, M.J.; Méndez, V.; Agulló, L.; Seeger, M. Genomic and functional analyses of the gentisate and protocatechuate ring-cleavage pathways and related 3-hydroxybenzoate and 4-hydroxybenzoate peripheral pathways in Burkholderia xenovorans LB400. PLoS ONE 2013, 8, e56038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentes, S.; Barra, B.; Caporaso, J.G.; Seeger, M. From rare to dominant: A fine-tuned soil bacterial bloom during petroleum hydrocarbon bioremediation. Appl. Environ. Microbiol. 2015, 82, 888–896. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, A.; Bernal, P.; Maureira, D.; Ramos, N.; Vásquez, J.; Urrutia, H.; Gentina, J.C.; Aroca, G. Biofiltration of trimethylamine in biotrickling filter inoculated with Aminobacter aminovorans. Electron. J. Biotechnol. 2018, 33, 63–67. [Google Scholar] [CrossRef]
- Orellana, R.; Macaya, C.; Bravo, G.; Dorochesi, F.; Cumsille, A.; Valencia, R.; Rojas, C.; Seeger, M. Living at the frontiers of life: Extremophiles in Chile and their potential for bioremediation. Front. Microbiol. 2018, 9, 2309. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, J.; Liu, D. Co3O4/g-C3N4 hybrids for gas-phase Hg0 removal at low temperature. Processes 2019, 7, 279. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, J.; Letowski, J.; Barkay, T. Mer-mediated resistance and volatilization of Hg (II) under anaerobic conditions. Geomicrobiol. J. 2002, 19, 87–102. [Google Scholar] [CrossRef]
- Deckwer, W.-D.; Becker, F.U.; Ledakowicz, S.; Wagner-Döbler, I. Microbial removal of ionic mercury in a three-phase fluidized bed reactor. Environ. Sci. Technol. 2004, 38, 1858–1865. [Google Scholar] [CrossRef] [PubMed]
- Mahbub, K.; Krishnan, K.; Megharaj, M.; Naidu, R. Bioremediation potential of a highly mercury resistant bacterial strain Sphingobium SA2 isolated from contaminated soil. Chemosphere 2016, 144, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Dash, H.R.; Basu, S.; Das, S. Evidence of mercury trapping in biofilm-EPS and mer operon-based volatilization of inorganic mercury in a marine bacterium Bacillus cereus BW-201B. Arch. Microbiol. 2016, 199, 445–455. [Google Scholar] [CrossRef]
- Wang, X.; He, Z.; Luo, H.; Zhang, M.; Zhang, D.; Pan, X.; Gadd, G.M. Multiple-pathway remediation of mercury contamination by a versatile selenite-reducing bacterium. Sci. Total Environ. 2018, 615, 615–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Lin, H.; Zheng, W.; Tomanicek, S.J.; Johs, A.; Feng, X.; Elias, D.A.; Liang, L.; Gu, B. Oxidation and methylation of dissolved elemental mercury by anaerobic bacteria. Nat. Geosci. 2013, 6, 751–754. [Google Scholar] [CrossRef]
- Lu, X.; Liu, Y.; Johs, A.; Zhao, L.; Wang, T.; Yang, Z.; Lin, H.; Elias, D.A.; Pierce, E.M.; Liang, L.; et al. Anaerobic mercury methylation and demethylation by Geobacter bemidjiensis Bem. Environ. Sci. Technol. 2016, 50, 4366–4373. [Google Scholar] [CrossRef] [PubMed]
- Mergeay, M.; Nies, D.; Schlegel, H.G.; Gerits, J.; Charles, P.; Van Gijsegem, F. Alcaligenes eutrophus CH34 is a facultative chemolithotroph with plasmid-bound resistance to heavy metals. J. Bacteriol. 1985, 162, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Alviz-Gazitua, P.; Fuentes-Alburquenque, S.; Rojas, L.A.; Turner, R.; Guiliani, N.; Seeger, M. The response of Cupriavidus metallidurans CH34 to cadmium involves inhibition of the initiation of biofilm formation, decrease in intracellular c-di-GMP levels, and a novel metal regulated phosphodiesterase. Front. Microbiol. 2019, 10, 1499. [Google Scholar] [CrossRef]
- Millacura, F.A.; Janssen, P.J.; Monsieurs, P.; Janssen, A.; Provoost, A.; Van Houdt, R.; Rojas, L.A. Unintentional genomic changes endow Cupriavidus metallidurans with an augmented heavy-metal resistance. Genes 2018, 9, 551. [Google Scholar] [CrossRef] [Green Version]
- Espinoza-Tofalos, A.; Daghio, M.; González, M.; Papacchini, M.; Franzetti, A.; Seeger, M. Toluene degradation by Cupriavidus metallidurans CH34 in nitrate-reducing conditions and in bioelectrochemical systems. FEMS Microbiol. Lett. 2018, 365, fny119. [Google Scholar] [CrossRef]
- Zhang, J.; Greasham, R. Chemically defined media for commercial fermentations. Appl. Microbiol. Biotechnol. 1999, 51, 407–421. [Google Scholar] [CrossRef]
- Acevedo, A.; Conejeros, R.; Aroca, G. Ethanol production improvement driven by genome-scale metabolic modeling and sensitivity analysis in Scheffersomyces stipites. PLoS ONE 2017, 12, e0180074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casas López, J.L.; Rodríguez Porcel, E.M.; Oller Alberola, I.; Ballesteros Martín, M.M.; Sánchez Pérez, J.; Fernández Sevilla, J.M.; Chisti, Y. Simultaneous determination of oxygen consumption rate and volumetric oxygen transfer coefficient in pneumatically agitated bioreactors. Ind. Eng. Chem. Res. 2006, 45, 1167–1171. [Google Scholar] [CrossRef]
- AOAC Official Method 977.15 Mercury in Fish Alternative Flameless Atomic Absorption Spectrophotometric Method First Action 1977 Final Action 1978. Available online: http://www.aoacofficialmethod.org/index.php?main_page=product_info&cPath=1&products_id=2383 (accessed on 24 July 2020).
- Attalah, S.; Waller, P.; Steichen, S.; Gao, S.; Brown, C.; Ogden, K.; Brown, J. Application of deoxygenation-aeration cycling to control the predatory bacterium Vampirovibrio chlorellavorus in Chlorella sorokiniana cultures. Algal Res. 2019, 39, 101427. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, R.; Sun, C.; Cheng, T.; Liu, Y.; Xian, M. Fermentative succinate production: An emerging technology to replace the traditional petrochemical processes. BioMed Res. Int. 2013, 2013, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.B.; Von Meyenburg, K. Are growth rates of Escherichia coli in batch cultures limited by respiration? J. Bacteriol. 1980, 144, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ochoa, F.; Gomez, E.; Santos, V.E.; Merchuk, J.C. Oxygen uptake rate in microbial processes: An overview. Biochem. Eng. J. 2010, 49, 289–307. [Google Scholar] [CrossRef]
- Belyaeva, E.A.; Sokolova, T.V.; Emelyanova, L.V.; Zakharova, I.O. Mitochondrial electron transport chain in heavy metal-induced neurotoxicity: Effects of cadmium, mercury, and copper. Sci. World J. 2012, 2012, 136063. [Google Scholar] [CrossRef] [Green Version]
- Norambuena, J.; Wang, Y.; Hanson, T.; Boyd, J.M.; Barkay, T. Low-molecular-weight thiols and thioredoxins are important players in Hg (II) resistance in Thermus thermophilus HB27. Appl. Environ. Microbiol. 2018, 84, e01931-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Robison, T.; Wiatrowski, H. The impact of ionic mercury on antioxidant defenses in two mercury-sensitive anaerobic bacteria. BioMetals 2013, 26, 1023–1031. [Google Scholar] [CrossRef]
- Asztalos, E.; Sipka, G.; Kis, M.; Trotta, M.; Maróti, P. The reaction center is the sensitive target of the mercury (II) ion in intact cells of photosynthetic bacteria. Photosynth. Res. 2012, 112, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Mieiro, C.; Pardal, M.; Duarte, A.C.; Pereira, E.; Palmeira, C. Impairment of mitochondrial energy metabolism of two marine fish by in vitro mercuric chloride exposure. Mar. Pollut. Bull. 2015, 97, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Dash, H.R.; Das, S. Bioremediation of mercury and the importance of bacterial mer genes. Int. Biodeterior. Biodegrad. 2012, 75, 207–213. [Google Scholar] [CrossRef]
- Shimizu, K.; Matsuoka, Y. Redox rebalance against genetic perturbations and modulation of central carbon metabolism by the oxidative stress regulation. Biotechnol. Adv. 2019, 37, 107441. [Google Scholar] [CrossRef] [PubMed]
- Bergkessel, M.; Basta, D.; Newman, D.K. The physiology of growth arrest: Uniting molecular and environmental microbiology. Nat. Rev. Microbiol. 2016, 14, 549–562. [Google Scholar] [CrossRef]
- Christodoulou, D.; Kuehne, A.; Estermann, A.; Fuhrer, T.; Lang, P.F.; Sauer, U. Reserve flux capacity in the pentose phosphate pathway by NADPH binding is conserved across kingdoms. iScience 2019, 19, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- Okino, S.; Iwasaki, K.; Yagi, O.; Tanaka, H. Development of a biological mercury removal-recovery system. Biotechnol. Lett. 2000, 22, 783–788. [Google Scholar] [CrossRef]
- Colombo, M.J.; Ha, J.; Reinfelder, J.R.; Barkay, T.; Yee, N. Oxidation of Hg (0) to Hg (II) by diverse anaerobic bacteria. Chem. Geol. 2014, 363, 334–340. [Google Scholar] [CrossRef]
- Ma, M.; Du, H.; Wang, D. Mercury methylation by anaerobic microorganisms: A review. Crit. Rev. Environ. Sci. Technol. 2019, 49, 1893–1936. [Google Scholar] [CrossRef]
- Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic metals and oxidative stress Part I: Mechanisms involved in metal-induced oxidative damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef]
- Maklashina, E.; Berthold, D.A.; Cecchini, G. Anaerobic expression of Escherichia coli succinate dehydrogenase: Functional replacement of fumarate reductase in the respiratory chain during anaerobic growth. J. Bacteriol. 1998, 180, 5989–5996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, P.J.; Van Houdt, R.; Moors, H.; Monsieurs, P.; Morin, N.; Michaux, A.; Benotmane, M.A.; Leys, N.; Vallaeys, T.; Lapidus, A.; et al. The complete genome sequence of Cupriavidus metallidurans strain CH34, a master survivalist in harsh and anthropogenic environments. PLoS ONE 2010, 5, e10433. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo, G.; Vega-Celedón, P.; Gentina, J.C.; Seeger, M. Effects of Mercury II on Cupriavidus metallidurans Strain MSR33 during Mercury Bioremediation under Aerobic and Anaerobic Conditions. Processes 2020, 8, 893. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8080893
Bravo G, Vega-Celedón P, Gentina JC, Seeger M. Effects of Mercury II on Cupriavidus metallidurans Strain MSR33 during Mercury Bioremediation under Aerobic and Anaerobic Conditions. Processes. 2020; 8(8):893. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8080893
Chicago/Turabian StyleBravo, Guillermo, Paulina Vega-Celedón, Juan Carlos Gentina, and Michael Seeger. 2020. "Effects of Mercury II on Cupriavidus metallidurans Strain MSR33 during Mercury Bioremediation under Aerobic and Anaerobic Conditions" Processes 8, no. 8: 893. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8080893