Production of Hydrogen Sulfide by Fermentation in Rumen and Its Impact on Health and Production of Animals

 ,
,  ,
, 
 and
and 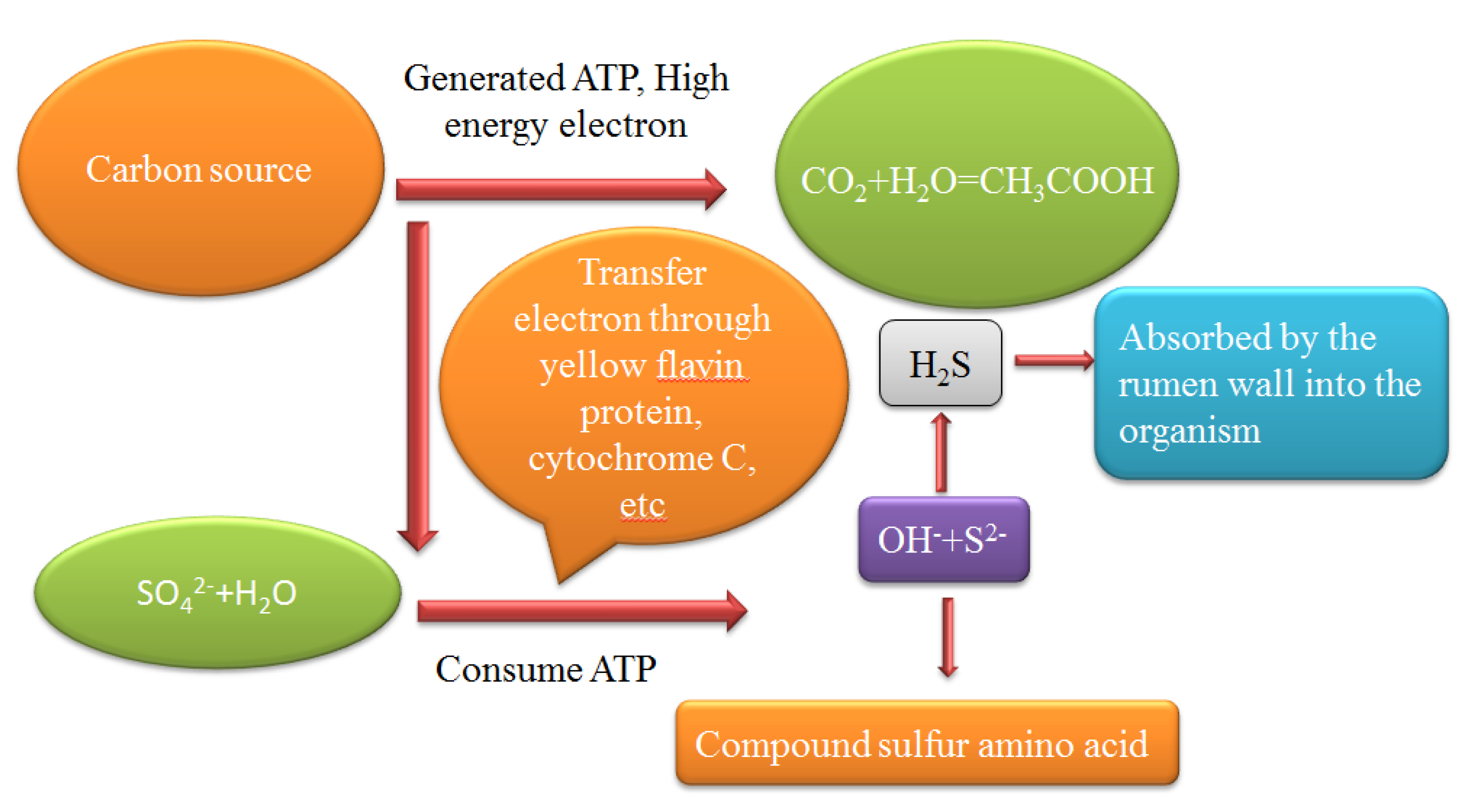{kind=link}
Abstract
:1. Introduction
2. Hydrogen Sulfide Toxicity in Rumen
3. Hydrogen Sulfide Production Pathway in Rumen
3.1. Hydrogen Sulfide Production in Rumen
3.2. Sulfide-Reducing Bacteria and Methanogens Relationship in the Rumen
4. Factors Affecting the Production of Rumen Hydrogen Sulfide
4.1. Dietary Sulfur Levels Influence the Production of Hydrogen Sulfide in the Rumen
4.2. Rumen pH Influences the Production of Hydrogen Sulfide
4.3. Dietary NDF Content Influences the Production of Hydrogen Sulfide in Rumen
4.4. Effect of Ruminal Hydrogen Sulfide on Ruminants Health/Beneficial Impact of Hydrogen Sulfide on Animals
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, T.W.; Wang, E.A.; Gould, S.; Stein, E.V.; Kaur, S.; Lim, L.; Amarnath, S.; Fowler, D.H.; Roberts, D.D. Hydrogen sulfide is an endogenous potentiator of T cell activation. J. Biol. Chem. 2012, 287, 4211–4221. [Google Scholar] [PubMed] [Green Version]
- Pal, V.K.; Bandyopadhyay, P.; Singh, A. Hydrogen sulfide in physiology and pathogenesis of bacteria and viruses. IUBMB Life 2018, 70, 393–410. [Google Scholar] [PubMed]
- Kimura, Y.; Dargusch, R.; Schubert, D.; Kimura, H. Hydrogen sulfide protects HT22 neuronal cells from oxidative stress. Antioxid. Redox Signal. 2006, 8, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Shatalin, K.; Shatalina, E.; Mironov, A.; Nudler, E. H2S: A universal defense against antibiotics in bacteria. Science 2011, 334, 986–990. [Google Scholar] [CrossRef]
- Aroca, A.; Gotor, C.; Bassham, D.C.; Romero, L.C. Hydrogen sulfNide: From a toxic molecule to a key molecule of cell life. Antioxidants 2020, 9, 621. [Google Scholar] [CrossRef]
- Wallace, J.L.; Ferraz, J.G.; Muscara, M.N. Hydrogen sulfide: An endogenous mediator of resolution of inflammation and injury. Antioxid. Redox Signal. 2012, 17, 58–67. [Google Scholar] [CrossRef]
- Pouokam, E.; Steidle, J.; Diener, M. Regulation of colonic ion transport by gasotransmitters. Biol. Pharm. Bull. 2011, 34, 789–793. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.L.; Dicay, M.; McKnight, W.; Martin, G.R. Hydrogen sulfide enhances ulcer healing in rats. FASEB J. 2007, 21, 4070–4076. [Google Scholar] [CrossRef]
- Magierowski, M.; Magierowska, K.; Hubalewska-Mazgaj, M.; Sliwowski, Z.; Pajdo, R.; Ginter, G.; Kwiecien, S.; Brzozowski, T. Exogenous and endogenous hydrogen sulfide protects gastric mucosa against the formation and time-dependent development of ischemia/reperfusion-induced acute lesions progressing into deeper ulcerations. Molecules 2017, 22, 295. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.L.; Wang, R. Hydrogen sulfide-based therapeutics: Exploiting a unique but ubiquitous gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [CrossRef]
- Binversie, E.Y.; Ruiz-Moreno, M.; Carpenter, A.; Heins, B.; Crawford, G.; DiCostanzo, A.; Stern, M. Effects of dietary roughage and sulfur in diets containing corn dried distillers grains with solubles on hydrogen sulfide production and fermentation by rumen microbes in vitro. J. Anim. Sci. 2016, 94, 3883–3893. [Google Scholar] [CrossRef] [PubMed]
- Gould, D.H.; Cummings, B.A.; Hamar, D.W. In vivo indicators of pathologic ruminal sulfide production in steers with diet-induced polioencephalomalacia. J. Vet. Diag. Investig. 1997, 9, 72–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchamp, R.; Bus, J.S.; Popp, J.A.; Boreiko, C.J.; Andjelkovich, D.A.; Leber, P. A critical review of the literature on hydrogen sulfide toxicity. CRC Crit. Rev. Toxicol. 1984, 13, 25–97. [Google Scholar] [CrossRef] [PubMed]
- Drewnoski, M.; Richter, E.; Hansen, S. Dietary sulfur concentration affects rumen hydrogen sulfide concentrations in feedlot steers during transition and finishing. J. Anim. Sci. 2012, 90, 4478–4486. [Google Scholar] [CrossRef]
- Gould, D.H. Polioencephalomalacia. J. Anim. Sci. 1998, 76, 309–314. [Google Scholar] [CrossRef]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D. The reduction of sulphate in the rumen of the sheep. Biochem. J. 1954, 56, 391. [Google Scholar] [CrossRef] [PubMed]
- Huisingh, J.; McNeill, J.; Matrone, G. Sulfate reduction by a Desulfovibrio species isolated from sheep rumen. Appl. Environ. Microbiol. 1974, 28, 489–497. [Google Scholar] [CrossRef] [Green Version]
- Howard, B.; Hungate, R.E. Desulfovibrio of the sheep rumen. Appl. Environ. Microbiol. 1976, 32, 598–602. [Google Scholar] [CrossRef] [Green Version]
- Cummings, B.; Gould, D.; Caldwell, D.; Hamar, D. Ruminal microbial alterations associated with sulfide generation in steers with dietary sulfate-induced polioencephalomalacia. Am. J. Vet. Res. 1995, 56, 1390–1395. [Google Scholar]
- Fron, M.J.; Boling, J.A.; Bush, L.P.; Dawson, K.A. Sulfur and nitrogen metabolism in the bovine fed different forms of supplemental sulfur. J. Anim. Sci. 1990, 68, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Sarturi, J.; Erickson, G.; Klopfenstein, T.; Rolfe, K.; Buckner, C.; Luebbe, M. Impact of source of sulfur on ruminal hydrogen sulfide and logic for the ruminal available sulfur for reduction concept. J. Anim. Sci. 2013, 91, 3352–3359. [Google Scholar] [CrossRef] [PubMed]
- Nava, G.M.; Carbonero, F.; Croix, J.A.; Greenberg, E.; Gaskins, H.R. Abundance and diversity of mucosa-associated hydrogenotrophic microbes in the healthy human colon. ISME J. 2012, 6, 57–70. [Google Scholar] [CrossRef]
- Jiang, G.; Gutierrez, O.; Sharma, K.R.; Yuan, Z. Effects of nitrite concentration and exposure time on sulfide and methane production in sewer systems. Water Res. 2010, 44, 4241–4251. [Google Scholar] [CrossRef]
- Singh, S.B.; Lin, H.C. Hydrogen sulfide in physiology and diseases of the digestive tract. Microorganisms 2015, 3, 866–889. [Google Scholar] [CrossRef] [Green Version]
- Bryant, M.P.; Campbell, L.L.; Reddy, C.A.; Crabill, M.R. Growth of Desulfovibrio in lactate or ethanol media low in sulfate in association with H2-utilizing methanogenic bacteria. Appl. Environ. Microbiol. 1977, 33, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- Zinder, S.H. Physiological ecology of methanogens. In Methanogenesis; Springer: Berlin/Heidelberg, Germany, 1993; pp. 128–206. [Google Scholar]
- Stefanie, J.O.E.; Visser, A.; Pol, L.W.H.; Stams, A.J. Sulfate reduction in methanogenic bioreactors. FEMS Microbiol. Rev. 1994, 15, 119–136. [Google Scholar]
- Colleran, E.; Finnegan, S.; Lens, P. Anaerobic treatment of sulphate-containing waste streams. Antonie van Leeuwenhoek 1995, 67, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Colleran, S.; Pender, S. Mesophilic and thermophilic anaerobic digestion of sulphate-containing wastewaters. Water Sci. Technol. 2002, 45, 231–235. [Google Scholar] [CrossRef]
- Richter, E.L. The Effect of Dietary Sulfur on Performance, Mineral Status, Rumen Hydrogen Sulfide, and Rumen Microbial Populations in Yearling Beef Steers. Master’s Thesis, Iowa State University, Ames, IA, USA, 2011. [Google Scholar]
- Crane, A.; Redden, R.; Swanson, K.; Howard, B.; Frick, T.; Maddock-Carlin, K.; Schauer, C. Effects of dried distiller’s grains and lasalocid inclusion on feedlot lamb growth, carcass traits, nutrient digestibility, ruminal fluid volatile fatty acid concentrations, and ruminal hydrogen sulfide concentration. J. Anim. Sci. 2017, 95, 3198–3205. [Google Scholar] [PubMed]
- Drewnoski, M.; Beitz, D.C.; Loy, D.D.; Hansen, S.L.; Ensley, S.M. Factors affecting ruminal hydrogen sulfide concentration of cattle. Anim.l Ind. Rep. 2011, 657, 11. [Google Scholar]
- Loneragan, G.; Wagner, J.; Gould, D.; Garry, F.; Thoren, M. Effects of water sulfate concentration on performance, water intake, and carcass characteristics of feedlot steers. J. Anim. Sci. 2001, 79, 2941–2948. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle; National Academies Press: Washington, DC, USA, 2000; Volume 22, pp. 351–369. [Google Scholar]
- National Research Council. Nutrient Requirements of Dairy Cattle; National Research Council: Washington, DC, USA, 2001. [Google Scholar]
- Kung, L., Jr.; Bracht, J.P. High sulfate induced polioencephalomalacia PEM in cattle—Burping can be dangerous if you are a ruminant. In Proceedings of the Chr. Hansen’s Technical Symposium, Pacific Northwest Nutrition Conference, Vancouver, BC, Canada, 13–15 October 1998. [Google Scholar]
- Smith, D.R.; Dilorenzo, N.; Leibovich, J.; May, M.L.; Quinn, M.J.; Homm, J.W.; Galyean, M.L. Effects of sulfur and monensin concentrations on in vitro dry matter disappearance, hydrogen sulfide production, and volatile fatty acid concentrations in batch culture ruminal fermentations. J. Anim. Sci. 2010, 88, 1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.; Liu, H. Effect of pH on hydrogen production from glucose by a mixed culture. Bioresour. Technol. 2002, 82, 87–93. [Google Scholar] [CrossRef]
- May, M.L.; Declerck, J.C.; Leibovich, J.; Quinn, M.J.; Galyean, M.L. Corn or sorghum wet distillers grains with solubles in combination with steam-flaked corn: In vitro fermentation and hydrogen sulfide production. J. Anim. Sci. 2010, 88, 2425–2432. [Google Scholar] [CrossRef]
- Price, J.; Will, A.; Paschaleris, G.; Chesters, J. Identification of thiomolybdates in digesta and plasma from sheep after administration of 99Mo-labelled compounds into the rumen. Br. J. Nutr. 1987, 58, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Huber, J.T.; Price, N.O.; Engel, R.W. Response of lactating dairy cows to high levels of dietary molybdenum. J. Anim. Sci. 1971, 32, 364–367. [Google Scholar] [CrossRef]
- Majak, W.; Steinke, D.; McGillivray, J.; Lysyk, T. Clinical signs in cattle grazing high molybdenum forage. J. Range Manag. 2004, 57, 269–274. [Google Scholar] [CrossRef]
- Kung, L.; Bracht, J.P.; Tavares, J.Y. Effects of various compounds on in vitro ruminal fermentation and production of sulfide. Anim. Feed Sci. Technol. 2000, 84, 69–81. [Google Scholar] [CrossRef]
- Taylor, B.F.; Oremland, R.S. Depletion of adenosine triphosphate in desulfovibrio by oxyanions of group VI elements. Curr. Microbiol. 1979, 3, 101–103. [Google Scholar] [CrossRef]
- Neville, B.; Lardy, G.; Karges, K.; Kirschten, L.; Schauer, C. Sulfur intake, excretion, and ruminal hydrogen sulfide concentrations in lambs fed increasing concentrations of distillers dried grains with solubles. Sheep Goat Res. J. 2011, 26, 13–19. [Google Scholar]
- Neville, B.; Schauer, C.; Karges, K.; Gibson, M.; Thompson, M.; Kirschten, L.; Dyer, N.; Berg, P.; Lardy, G. Effect of thiamine concentration on animal health, feedlot performance, carcass characteristics, and ruminal hydrogen sulfide concentrations in lambs fed diets based on 60% distillers dried grains plus solubles. J. Anim. Sci. 2010, 88, 2444–2455. [Google Scholar] [CrossRef] [Green Version]
- Coleman, G. A sulphate-reducing bacterium from the sheep rumen. Microbiology 1960, 22, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Rumsey, T. Effects of dietary sulfur addition and Synovex-S ear implants on feedlot steers fed an all-concentrate finishing diet. J. Anim. Sci. 1978, 46, 463–477. [Google Scholar] [CrossRef]
- Morine, S.; Drewnoski, M.; Hansen, S. Increasing dietary neutral detergent fiber concentration decreases ruminal hydrogen sulfide concentrations in steers fed high-sulfur diets based on ethanol coproducts. J. Anim. Sci. 2014, 92, 3035–3041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morine, S.; Drewnoski, M.; Johnson, A.; Hansen, S. Determining the influence of dietary roughage concentration and source on ruminal parameters related to sulfur toxicity. J. Anim. Sci. 2014, 92, 4068–4076. [Google Scholar] [CrossRef]
- Yan, L.; Ye, J.; Zhang, P.; Xu, D.; Wu, Y.; Liu, J.; Zhang, H.; Fang, W.; Wang, B.; Zeng, G. Hydrogen sulfide formation control and microbial competition in batch anaerobic digestion of slaughterhouse wastewater sludge: Effect of initial sludge pH. Bioresour. Technol. 2018, 259, 67–74. [Google Scholar] [CrossRef]
- Uwituze, S.; Parsons, G.; Karges, K.; Gibson, M.; Hollis, L.; Higgins, J.; Drouillard, J. Effects of distillers grains with high sulfur concentration on ruminal fermentation and digestibility of finishing diets. J. Anim. Sci. 2011, 89, 2817–2828. [Google Scholar] [CrossRef]
- National Research Council. Mineral Tolerance of Animals; National Academies Press: Washington, DC, USA, 2005. [Google Scholar]
- Gooneratne, S.R.; Olkowski, A.A.; Christensen, D.A. Sulfur-induced polioencephalomalacia in sheep: Some biochemical changes. Can. J. Vet. Res. 1989, 53, 462–467. [Google Scholar]
- Delfiol, D.J.Z.; Cunha, P.H.J.D.; Borges, A.S. Determination of ruminal hydrogen sulfide in sheep. Vet. Zootec. 2011, 18, 3625–3628. [Google Scholar]
- Kessler, K.L.; Olson, K.C.; Wright, C.L.; Austin, K.J.; Johnson, P.S.; Cammack, K.M. Effects of supplemental molybdenum on animal performance, liver copper concentrations, ruminal hydrogen sulfide concentrations, and the appearance of sulfur and molybdenum toxicity in steers receiving fiber-based diets. J. Anim. Sci. 2012, 90, 5005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brent, B.E.; Bartley, E.E. Thiamin and niacin in the rumen. J. Anim. Sci. 1984, 59, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Olkowski, A.A.; Gooneratne, S.R.; Rousseaux, C.G.; Christensen, D.A. Role of thiamine status in sulphur induced polioencephalomalacia in sheep. Res. Vet. Sci. 1992, 52, 78. [Google Scholar] [CrossRef]
- Mcdowell, L.R. Vitamins in Animal and Human Nutrition, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2008. [Google Scholar]
- Galyean, M.L.; Rivera, J.D. Nutritionally related disorders affecting feedlot cattle. Can. J. Anim. Sci. 2003, 83, 13–20. [Google Scholar] [CrossRef]
- Felix, T.L.; Zerby, H.N.; Moeller, S.J.; Loerch, S.C. Effects of increasing dried distillers grains with solubles on performance, carcass characteristics, and digestibility of feedlot lambs. J. Anim. Sci. 2012, 90, 1356–1363. [Google Scholar] [CrossRef] [Green Version]
- Magierowski, M.; Magierowska, K.; Hubalewska-Mazgaj, M.; Adamski, J.; Bakalarz, D.; Sliwowski, Z.; Pajdo, R.; Kwiecien, S.; Brzozowski, T. Interaction between endogenous carbon monoxide and hydrogen sulfide in the mechanism of gastroprotection against acute aspirin-induced gastric damage. Pharmacol. Res. 2016, 114, 235–250. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, A.M.; Ma, J.; Wang, Z.; Hu, R.; Wang, X.; Peng, Q.; Amevor, F.K.; Goswami, N. Production of Hydrogen Sulfide by Fermentation in Rumen and Its Impact on Health and Production of Animals. Processes 2020, 8, 1169. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091169
Shah AM, Ma J, Wang Z, Hu R, Wang X, Peng Q, Amevor FK, Goswami N. Production of Hydrogen Sulfide by Fermentation in Rumen and Its Impact on Health and Production of Animals. Processes. 2020; 8(9):1169. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091169
Chicago/Turabian StyleShah, Ali Mujtaba, Jian Ma, Zhisheng Wang, Rui Hu, Xueying Wang, Quanhui Peng, Felix Kwame Amevor, and Naqash Goswami. 2020. "Production of Hydrogen Sulfide by Fermentation in Rumen and Its Impact on Health and Production of Animals" Processes 8, no. 9: 1169. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091169