1. Introduction
Family farms, as new agricultural business entities, partake in more commercially oriented agricultural production and management than their forerunners (“farming households”) in China, even though their main labour force still consists of family members, and the major source of their income remains agricultural work [
1]. Family farms are the main mode of management in the Inner Mongolian pastoral area of North China [
2]. Whole-plant corn silage, alfalfa hay, grass hay, crop straw, and natural grass are the major forages of family farms in the desert steppe of Inner Mongolia. In general, these farmers ensile whole-plant corn without any additives to save processing costs [
3]. Moreover, the microbial communities present in this form of whole-plant corn silage are still unclear, and understanding them is critical to explaining the fermentation quality and aerobic stability of such silage.
In the past decade, the development of next-generation sequencing technologies has helped to improve our understanding of the microbial communities present in silage [
4].
Lactobacillus dominates the bacterial succession and determines the fermentation quality of whole-plant corn silage during fermentation process [
5,
6]. The main bacterial species present in the silage at the end of the process are
Lactobacillus acetotolerans,
Lactobacillus silagei,
Lactobacillus parafarraginis,
Lactobacillus buchneri, and
Lactobacillus odoratitofui [
7]. The geographical location where at the corn was grown influences the bacterial succession process taking place in whole-plant corn silage and the bacterial community in the final silage [
8,
9]. Drouin et al. [
10] reported that ensiling whole-plant corn with lactic acid bacteria (LAB) increases the Shannon indexes of bacterial and fungal communities and modifies the aerobic stability, and
Saccharomyces,
Issatchenkia, and
Kazachstania cause aerobic deterioration during aerobic exposure. Nevertheless, the effect of the geographical location on the fungal community and the aerobic stability of whole-plant corn silage has not yet been reported on.
Recently, the silage metabolome has attracted new interest [
11]. Some studies have reported the effect of inoculating on the metabolite contents of whole-plant corn silage, alfalfa silage, and sainfoin silage with LAB [
6,
9,
12]; the dynamics of metabolites in whole-plant corn silages during fermentation process [
6]; and the correlation of the main metabolites with the main bacterial species and fermentation quality in whole-plant corn silages [
6]. Additionally, Wu et al. [
13] analysed the metabolic profiles of high-moisture sweetcorn kernel silages. Biogenic amines—a class of metabolites in silage—affect the palatability of silages, and the feed intake and performance of ruminants [
14]. However, the concentration and composition of the metabolites—especially the biogenic amines in silages on the desert steppe—remain unclear.
In the present study, whole-plant corn silages on family farms in the desert steppe of Inner Mongolia in North China were assessed after about 300 d of ensiling. The hypothesis of the present study was the differences in microbial communities, metabolites, and aerobic stability between whole-plant corn silages from different areas. The objective was to analyse the bacterial and fungal communities, the metabolites, the fermentation quality and the aerobic stability of whole-plant corn silages.
4. Discussion
In the present study, the materials displayed sufficient LAB counts (>5 log
10 CFU/g FW) and WSC concentrations (>97 g/kg DM) (
Table 1), resulting in a satisfactory fermentation quality for the whole-plant corn silages (
Table 5). Before ensiling, S2 and S3 contained greater LAB and WSC contents and a lower DM content than S1 and S4 pre-ensiling (
Table 1), indicating the conditions of the former two were more conducive to LAB fermentation during the ensiling process. This contributed to the lower pH and higher lactic acid contents in S2 and S3 (
Table 5).
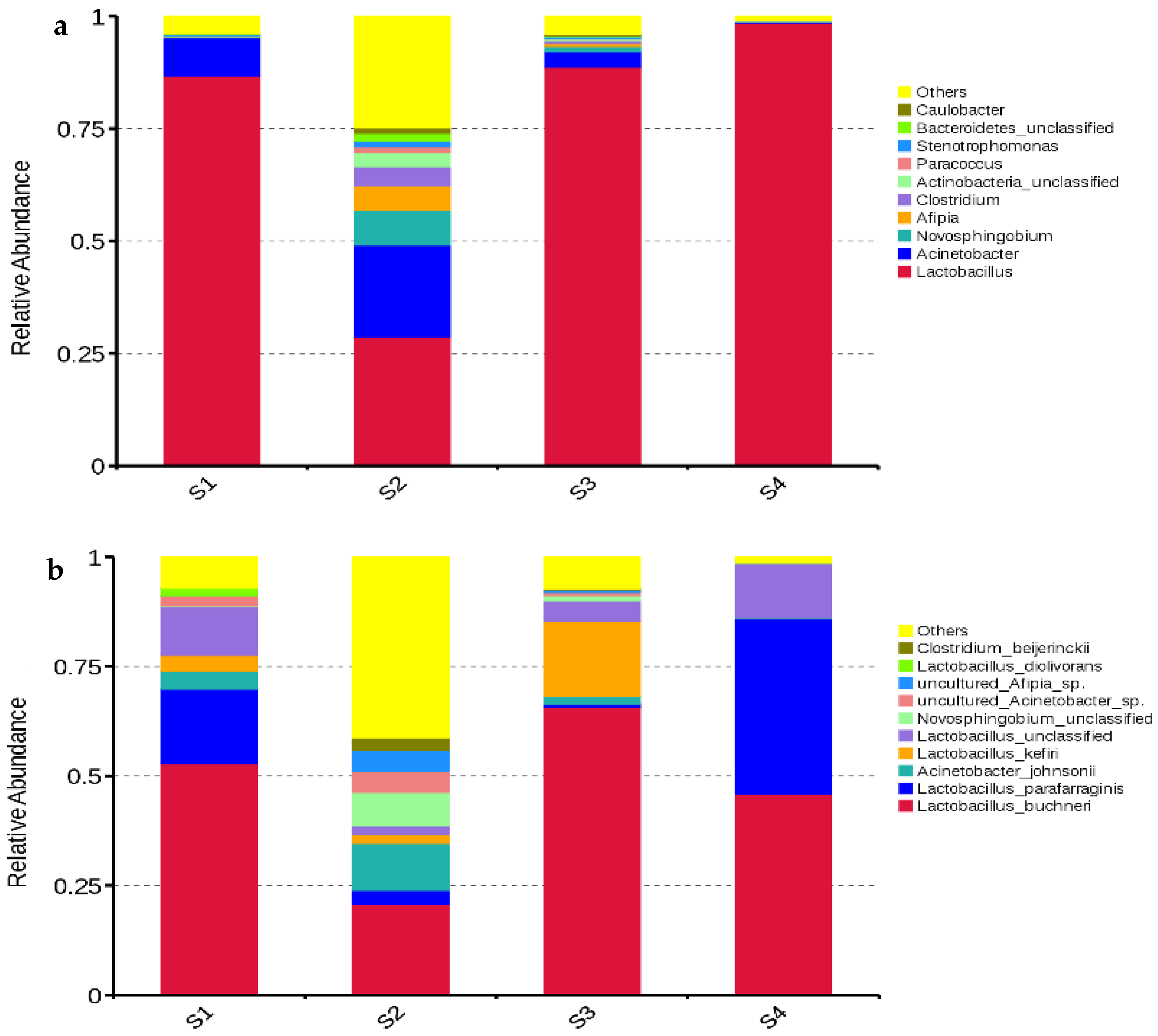
As the microbial community composition plays a crucial role in the fermentation quality and aerobic stability of silage, it is necessary to assess the bacterial and fungal communities in order to explain the different fermentation quality and aerobic stability among silages. In the present study,
Lactobacillus was the most dominant bacterial genus in all silages, followed by
Acinetobacter in S1 and S3, and by
Acinetobacter,
Novosphingobium,
Afipia,
Clostridium, and unclassified
Actinobacteria in S2 (
Figure 3a). This indicated that
Lactobacillus is generally the dominant bacterial genus in whole-plant corn silages after ensiling for about 300 d. Moreover, previous studies also reported
Lactobacillus at the greatest abundance in whole-plant corn silages collected from five regions in Southwest China [
9] and three sites in Iran [
8]. In silage with a good fermentation quality, the bacterial community is dominated by
Lactobacillus [
21], explaining the satisfactory fermentation quality of whole-plant corn silages in our study.
Acinetobacter was found in sugarcane top silage [
22], and
Novosphingobium was detected in whole-plant corn silage [
23] and red clover silage [
24]. However, the effect of these genera to fermentation quality and aerobic stability are not clear and require further research.
In the present study, the main bacterial species in S2 were
L. buchneri,
Acinetobacter johnsonii, and unclassified
Novosphingobium with abundances of 20.7%, 10.7% and 7.64%, respectively (
Figure 3b).
Lactobacillus buchneri,
L. parafarraginis,
L. kefiri, and unclassified
Lactobacillus dominated the bacterial communities in S1, S3, and S4 with total abundances of 84.5%, 88.2%, and 98.3%, respectively (
Figure 3b). The main LAB species in the silages after ensiling for about 300 d were
L. buchneri,
L. parafarraginis,
L. kefiri, and
L. diolivorans (
Figure 3b), which belong to
L. buchneri group as the sole heterofermentative LAB [
25]. However, in previous studies, the main LAB species were
L. acetotolerans,
L. silagei,
L. buchneri,
L. odoratitofui,
L. farciminis, and
L. parafarraginis in whole-plant corn silages (90 d) with low pH (from 3.68 to 3.74) [
7],
L. plantarum,
L. buchneri, and
L. brevisi in alfalfa silage (90 d) with high pH (from 4.9 to 5.2) [
26],
L. plantarum,
L. hammesi,
L. brevisi,
L. coryniformis, and
L. piscium in Italian ryegrass silages (42 d) with pH ranging from 4.40 to 5.52 [
10]. These studies suggested that homofermentative LAB co-existed with heterofermentative LAB in short silage (42–90 d). The differences in LAB population between our study and previous studies might result from the differences in silage age (300 d vs. 42–90 d), in the epiphytic population of bacteria on materials, and in the quality of the silages. The effects of short- and long-term storage on the microbial community and fermentation quality of silage requires further study. In the present study, the activities of heterofermentative LAB were weak in S2 and S3 under more acidic conditions (pH = 3.61 and 3.63, respectively) for a long period (about 300 d after ensiling), while they were stronger in S1 and S4 (pH 3.97). This is reflected in the higher acetic acid contents and LAB counts, and the lower lactic acid concentrations, in the latter at 0 and 2 d, along with greater aerobic stability (
Table 5). The bacteria count decreased in S2 in the first 2 d, while it increased in S3 and S4 (
Table 2). This might be because S2 still had a lower pH (3.69) and higher lactic acid content (121 g/kg DM) at 2 d than S1, S3, and S4 (
Table 5). More acidic conditions might have reduced the bacterial activity in the silage, although the silage had been exposed to air for 2 d.
Acinetobacter johnsonii, a known spoilage organism [
27], was one of the more dominant bacterial species in S1 and S2, and it was present in S3 and S4 at low levels (
Figure 3b). The effect of
A. johnsonii on the fermentation quality and safety of silage requires further study.
The main fungal genera in S1 and S4 were
Kazachstania,
Candida,
Cladosporium,
Rhizopus,
Rhizomucor,
Alternaria, and
Epicoccum, whereas
Kazachstania dominated the fungal communities in S2 and S3 (
Figure 4a). Other studies have reported dominant fungal genera of
Candida and
Monascus in whole-plant corn silage (90 d) [
28],
Pichia,
Sporisorium, Meyerozyma, and
Hannaella in sugarcane top silage (60 d) [
22],
Issatchenki in barley silage (60 d) [
29], and
Candida,
Kazachstania, and
Pichia in sugarcane top silage (90 d) [
15]. These results suggest that, in contrast to the bacterial community, the dominant fungal community in the silage was different between the silages at the genera level. In the present study,
Kazachstania dominated the fungal communities in S2 and S3 with poor aerobic stability (
Figure 5). S2 and S3 had lower pH and acetic acid, and higher lactic acid than the other silages (
Table 5). During aerobic exposure, the pH increased and the lactic acid decreased in S3, with acetic acid reducing at 2 d and increasing rapidly at 5 d. Between 2 and 5 d in S2, the pH increased quickly and the lactic acid decreased rapidly, with acetic acid disappearing at 5 d (
Table 5). These results agree with the findings of Wang et al. for sugarcane top silage [
15]. The results above indicate that
Kazachstania might have a strong tolerance to lactic acid and be a yeast species that is crucially involved in initiating the aerobic deterioration of silage with a relatively low pH and acetic acid content.
Kazachstania was also detected in barley silage [
29] and sugarcane top silage [
22] as minor taxa.
Candida was the predominant fungal genus in S1 and S4 (
Figure 4a), which had greater aerobic stability and higher acetic acid content (
Table 5), and lower temperatures, than other silages (
Figure 6) during aerobic exposure. However, Liu et al. [
29] and Romero et al. [
30] reported that a high abundance of
Candida sp. was associated with a lower aerobic stability in silage, owing to
Candida assimilating lactic acid after aerobic exposure [
28]. The difference between our results and those of previous studies might be due to the higher acetic acid concentration in our study (42.8 g/kg) than in the studies of Romero et al. [
4] and Liu et al. [
29] (7.4 g/kg and 18.3 g/kg, respectively)—acetic acid inhibits fungus growth in silage and can enhance aerobic stability [
4,
28,
30,
31].
In the present study, the dominant fungal species in S1 and S4 were unclassified
Kazachstania,
K. bulderi,
C. xylopsoci, unclassified
Cladosporium,
Rhizopus microspores, and
C. glabrata with total abundances of 78.5% and 84.9% in fungal communities, respectively (
Figure 4b). Unclassified
Kazachstania and
K. bulderi dominated the fungal communities in S2 and S3, respectively with abundances of 96.2% in S2 and 93.6% in S3 (
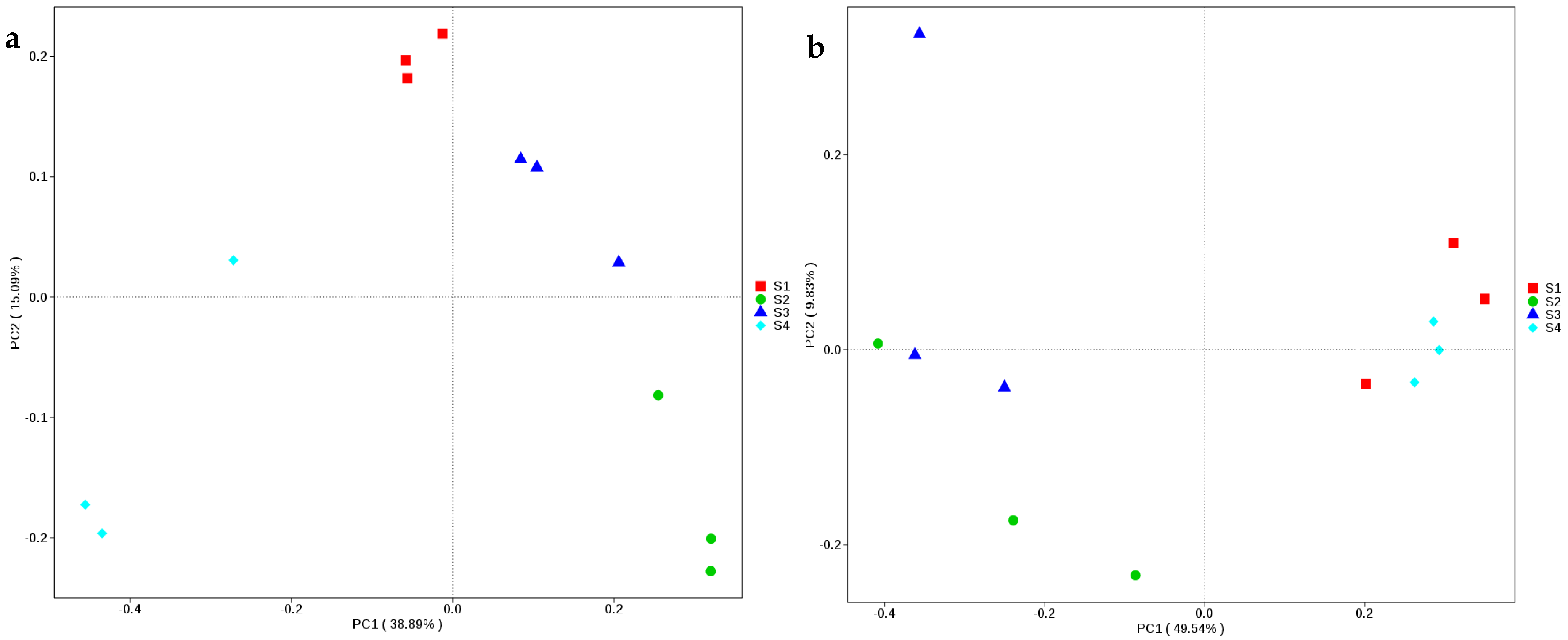
Figure 4b). According to PCoA, the fungal species in S1 and S4 were connected, and were clearly separated from those of S2 and S3 (
Figure 2). S1 and S4 thus had similar fungal species compositions, which were differentiated from those of S2 and S3. The differences in fungal species might be due to the differences in the fermentation processes of silage and in the epiphytic populations of fungi in the plant prior to ensiling [
28]. Moreover, unclassified
Kazachstania, and
K. bulderi belonging to
Kazachstania, were the dominant fungi in S2 and S3, respectively (
Figure 4b), and they had poor aerobic stability (
Figure 6). This indicates that the aerobic deterioration taking place in the two silages might result from
Kazachstania sp. growth after opening. Wang et al. [
15] found
K. humilis to be the key fungal yeast initiating aerobic deterioration in sugarcane top silage.
Kazachstania bulderi was the predominant fungus in S3 and S4 in the present study. This strain was formerly named
Saccharomyces bulderi [
32] and was first isolated from maize silage as a novel species [
33]. The present study first detected
C. xylopsoci,
R. microspores, and
Rhizomucor pusillus in the silage in the form of fungal pathogens.
Candida xylopsoci is usually isolated via fuel ethanol fermentation processes [
34],
R. microsporus has been detected in the traditional Chinese liquor
Daqu using 18S rRNA gene sequencing [
35], and
R. pusillus has been identified in immunocompromised patients using real-time PCR [
36]. The effects of these fungi on the fermentation quality and aerobic stability of silage should be investigated in future studies.
Moreover, Drouin et al. [
10] revealed that ensiling whole-plant corn with LAB inoculant improved the aerobic stability of the silage, by maintaining a higher microbial diversity (Shannon index of bacteria and fungi), avoiding the dominance of a few bacteria, and preventing fungi from damaging silage quality. However, in the present study, the fungal communities in S1 and S4 had higher Shannon, Simpson, and Chao1 indexes, and greater aerobic stability, than S2 and S3, but the bacterial communities in S1, S3, and S4 had lower Shannon, Simpson, and Chao1 indexes than S2 (
Table 3,
Figure 6). This suggests that high fungal diversity, rather than bacterial diversity, contributes most significantly to aerobic stability in whole-plant corn silage.
S2 displayed the highest bacterial Shannon and Simpson indexes among the silages (
Table 3), suggesting S2 had greater bacterial diversity in the present study. Additionally, the relative concentration of total metabolites in S2 was the highest among the silages (
Table 4), and containing 93, 103, and 81 more metabolites identified than S1, S3, and S4, respectively (
Supplementary Tables S2–S4). Previous studies showed that inoculating with LAB at ensiling increased the relative concentrations of some metabolites (e.g., organic acids, amino acids, and fatty acids) as well as the α-diversity (Shannon index) in whole-plant corn silages, sainfoin silages, and alfalfa silages [
6,
7,
9,
12]. This indicates that the high bacterial diversity might contribute to the accumulation of metabolites in whole-plant corn silages during fermentation process.
Biogenic amines in silages are produced by the decarboxylation of amino acids via the activities of plant enzymes and microbial enzymes, which accumulate during fermentation process and influence the palatability of silages, as well as the feed intake and performance of ruminants [
14,
37,
38]. The intake of adequate amounts of biogenic amines can promote normal physiological activities in man and animals, but excessive amounts may result in food poisoning [
38]. In the present study, tyramine, putrescine, phenylethylamine, spermidine, and noradrenaline were detected in whole-plant corn silages; the relative concentrations of tyramine and putrescine were considerable in S1, S2 and S3. Steidlová and Kalac also found that the concentrations of tyramine and putrescine were greater than those of histamine, cadaverine, tryptamine, spermidine and spermine in maize silages in 1999 (62 silages) and 2000 (51 silages) [
14]. Nishino et al. [
37] reported that inoculating with
Lactobacillus casei reduced the concentrations of biogenic amines (histamine, tyramine, putrescine and cadaverine) in grass silage, maize silage, and total mixed ration silage, and that adding
L. buchneri lowered the contents of biogenic amines in grass silages. Additionally, the activities of some undesirable bacterial genera (
Clostridia,
Bacillus,
Klebsiella,
Escherichia,
Pseudomonas,
Citrobacter,
Proteus,
Salmonella,
Shigella, and
Photobacterium) might result in the accumulation of biogenic amines in silages during ensiling [
38,
39]. In the present study, S2 contained higher
Bacillus,
Pseudomonas, and
Salmonella contents than other silages, and greater
Escherichia and
Klebsiella than S3 and S4 (
Supplementary Table S5), which might explain the higher relative concentrations of biogenic amines in S2 (
Table 4).


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}