
Long-Term Cultivation of a Native Arthrospira platensis (Spirulina) Strain in Pozo Izquierdo (Gran Canaria, Spain): Technical Evidence for a Viable Production of Food-Grade Biomass

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Isolation and Maintenance
2.2. Outdoor Cultivation and Biomass Processing
2.2.1. Description of the Cultivation Site
2.2.2. Scaling-Up of the A. platensis BEA 1257B Strain
2.2.3. Experimental Setup, Culture Operation and Monitoring
2.2.4. Biomass Harvesting and Processing
Harvesting Parameters
2.3. Assessment and Identification of Biotic Contaminants
2.4. Biomass Analysis
2.5. Statistical Analysis
3. Results and Discussion
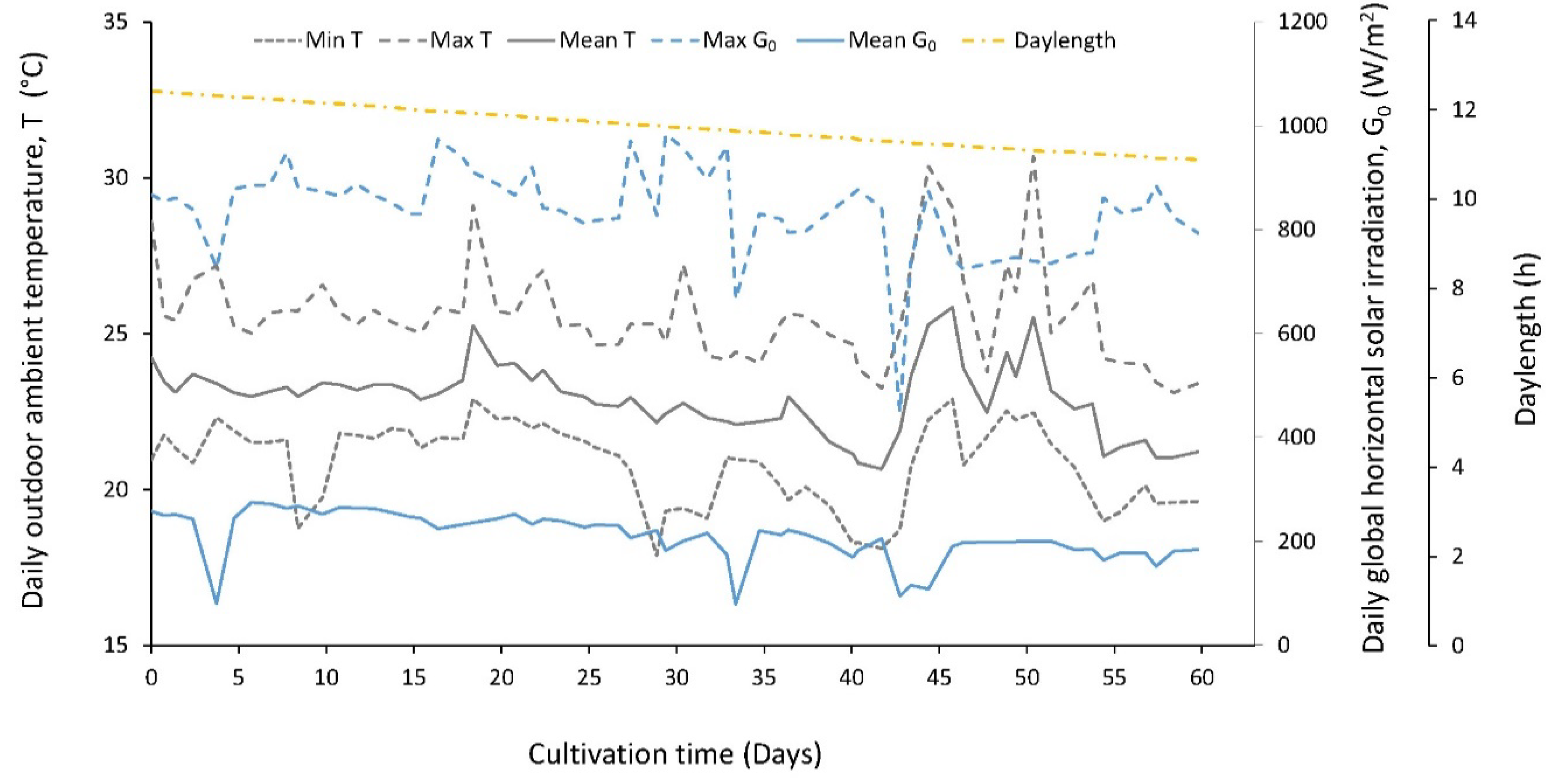
3.1. Environmental Parameters
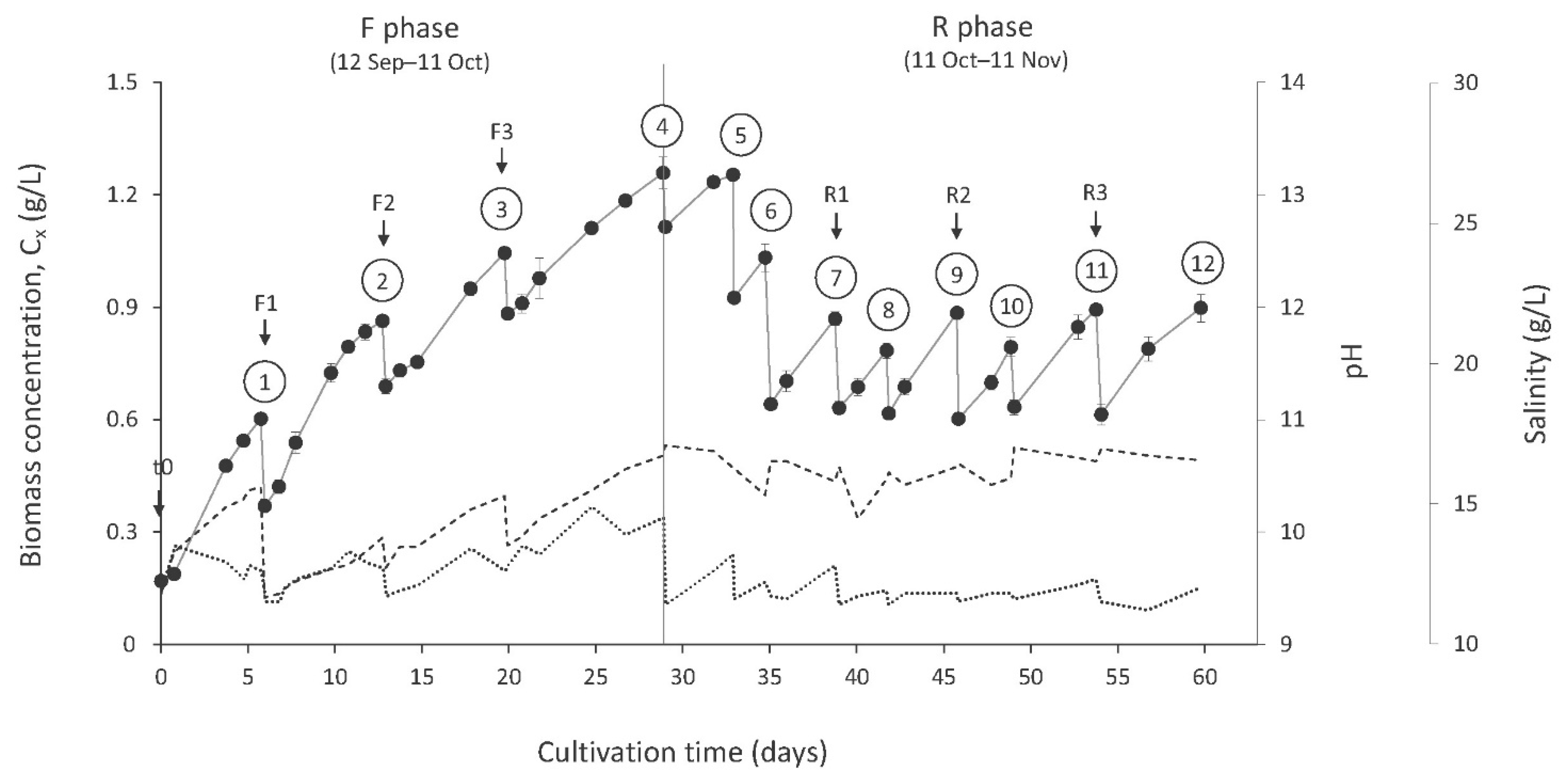
3.2. Culture Parameters
3.3. Culture Productivity and Harvesting Efficiency
3.4. Microzooplankton and Phototrophs Dynamics, and Culture Treatment
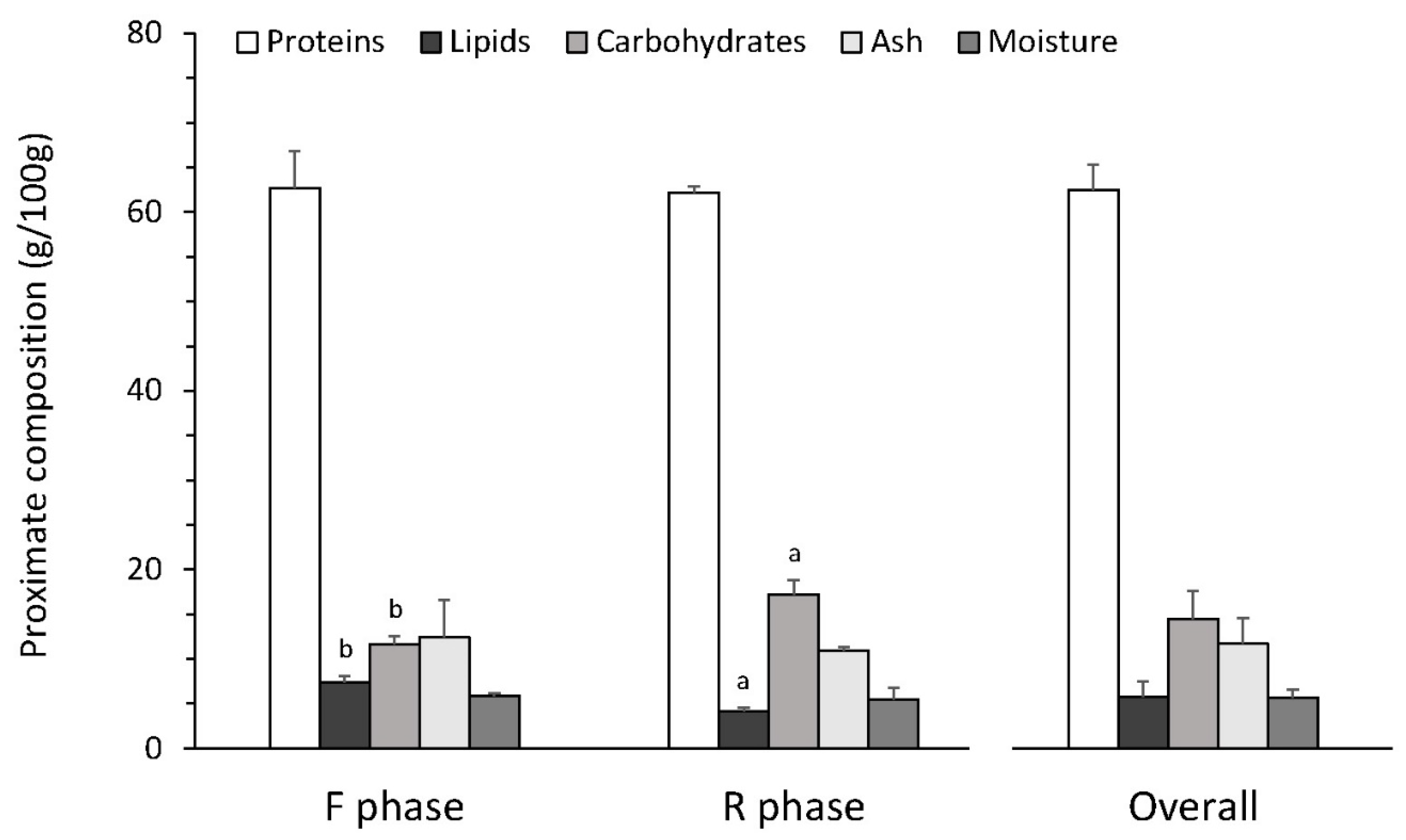
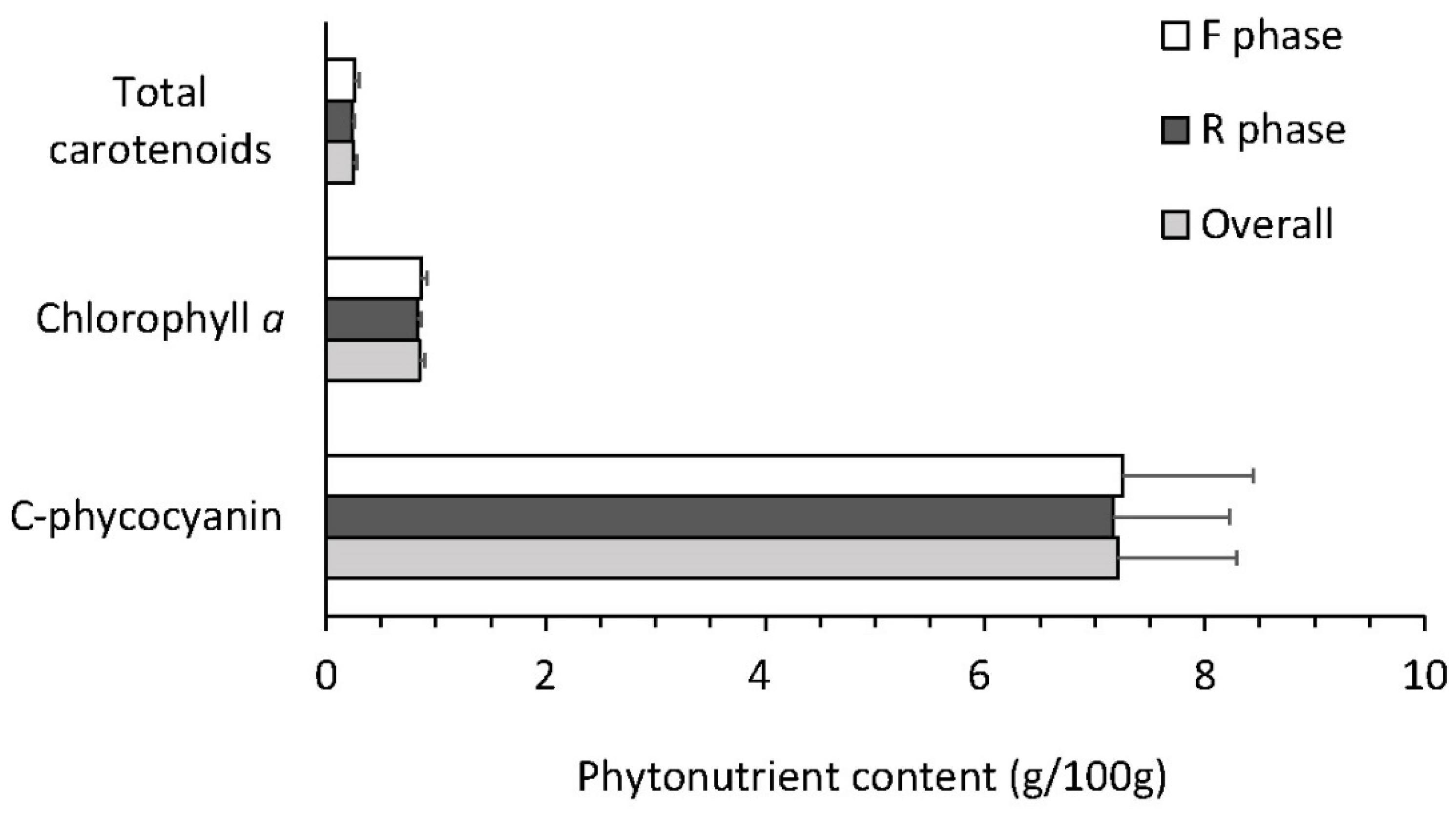
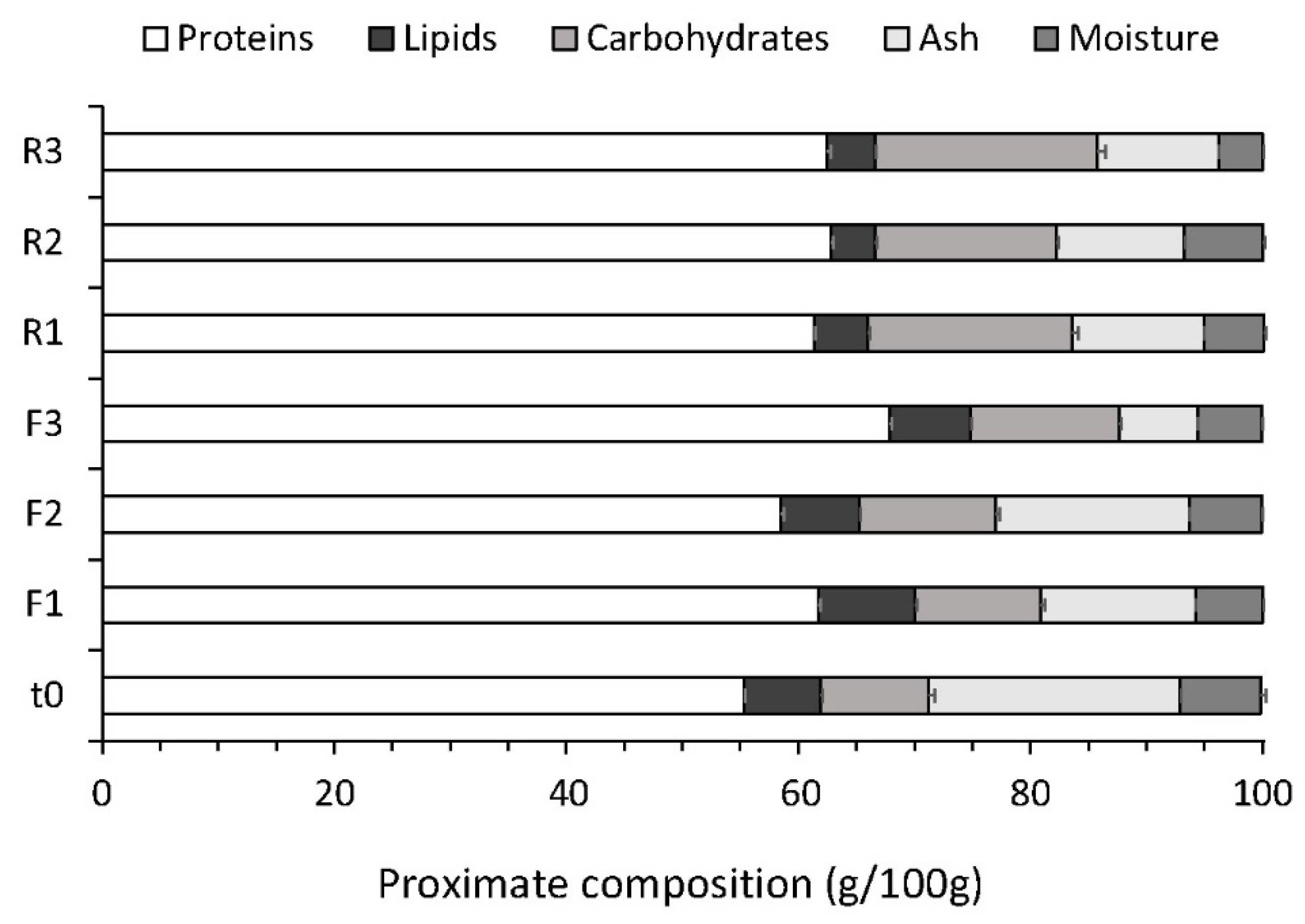
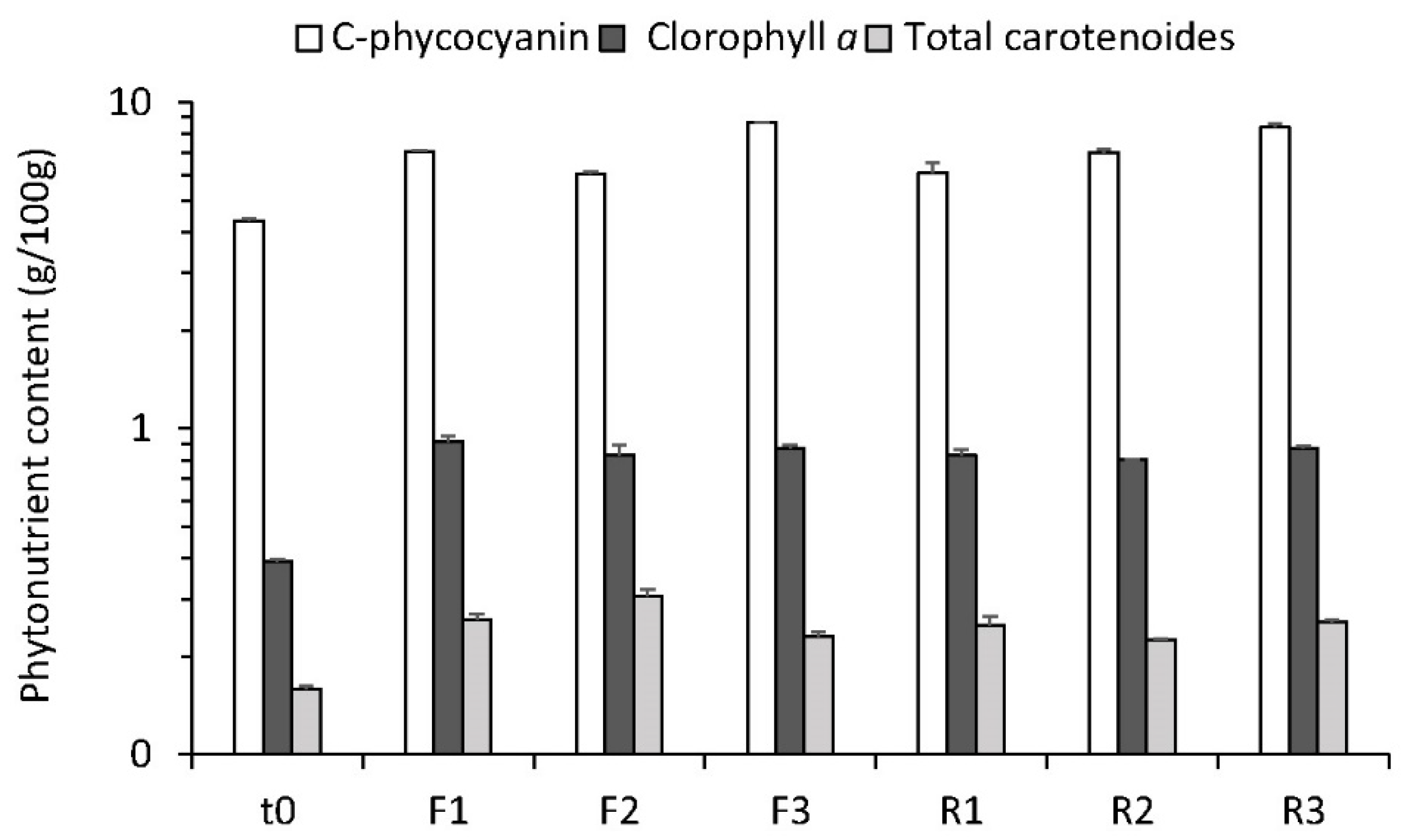
3.5. Biomass Profile and Quality
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Parameters | F Phase | R Phase | Overall | |||
|---|---|---|---|---|---|---|
| Range | Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | |
| Max daily G0 (W/m2) | 726.4–974.1 | 868.9 ± 50.8 * | 449.6–983.8 | 802.8 ± 104.2 * | 449.6–983.8 | 837.0 ± 87.0 |
| Mean daily G0 (W/m2) | 80.3–274.8 | 241.3 ± 35.6 * | 79.2–222.2 | 179.0 ± 36.7 * | 79.2–274.8 | 209.6 ± 47.6 |
| Daylength (h) | 11.7–12.4 | 12.0 ± 0.2 * | 10.9–11.6 | 11.2 ± 0.2 * | 10.9–12.4 | 11.6 ± 0.5 |
| Min daily T (°C) | 17.9–22.9 | 21.4 ± 1.0 * | 18.1–22.9 | 20.3 ± 1.4 * | 17.9–22.9 | 20.8 ± 1.3 |
| Max daily T (°C) | 24.6–29.1 | 25.9 ± 1.0 | 23.1–30.7 | 25.4 ± 2.0 | 23.1–30.7 | 25.6 ± 1.6 |
| Mean daily T (°C) | 22.1–25.3 | 23.3 ± 0.6 * | 20.7–25.8 | 22.5 ± 1.4 * | 20.7–25.8 | 22.9 ± 1.1 |

Appendix B
| Experimental Phase | Cycle No | Harvest (Day) | Cx (g/L DW) | HE (%) | Slurry Density (% DW) | CF | CFloss |
|---|---|---|---|---|---|---|---|
| F phase | 1 | 6 | 0.60 ± 0.010 | 74.4 ± 4.7 | 10.0 ± 0.15 | 166.1 ± 2.4 | 123.6 ± 1.8 |
| 2 | 13 | 0.86 ± 0.017 | 78.0 ± 3.8 | 10.3 ± 0.21 | 120.5 ± 2.3 | 94.0 ± 1.8 | |
| 3 | 20 | 1.04 ± 0.012 | 80.4 ± 3.8 | 9.5 ± 0.09 | 90.9 ± 0.8 | 73.1 ± 0.6 | |
| 4 | 29 | 1.26 ± 0.043 | 82.1 ± 2.8 | 9.0 ± 0.12 | 71.6 ± 1.1 | 58.7 ± 0.7 | |
| Cumulative F | 1–4 | 0.94 ± 0.28 | 78.7 ± 3.3 | 9.7 ± 0.6 | 87.4 ± 28.1 | 112.3 ± 41.1 |
| Schmidingerothrix sp. | C. sorokiniana | ||
|---|---|---|---|
| Time | Day 8 | Day 12 | |
| First detection | T (°C) | 23.4 | 25.3 |
| pH | 9.56 | 9.82 | |
| Salinity | 12.3 | 12.9 | |
| Cx (g/L) | 0.54 ± 0.029 | 0.83 ± 0.021 | |
| Chemical treatment | CO2 stop | Day 12 | |
| Add. NaHCO3 (4 g/L) | Day 13 | ||
| Add. (NH4)2SO4 (1mM) | Day 14, 20 | ||
| Last detection | Time | Day 15 | Day 20 * |
| T (°C) | 25.3 | 25.0 | |
| pH | 9.87 | 9.88 | |
| Salinity | 12.1 | 12.8 | |
| Cx (g/L) | 0.75 ± 0.010 | 0.88 ± 0.007 | |


References
- Alexandratos, N.; Bruinsma, J.; Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; FAO: Rome, Italy, 2012. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Elliott, J.; Deryng, D.; Ruane, A.C.; Müller, C.; Arneth, A.; Boote, K.J.; Folberth, C.; Glotter, M.; Khabarov, N.; et al. Assessing Agricultural Risks of Climate Change in the 21st Century in a Global Gridded Crop Model Intercomparison. Proc. Natl. Acad. Sci. USA 2014, 111, 3268–3273. [Google Scholar] [CrossRef] [Green Version]
- Muller, A.; Schader, C.; El-Hage Scialabba, N.; Brüggemann, J.; Isensee, A.; Erb, K.-H.; Smith, P.; Klocke, P.; Leiber, F.; Stolze, M.; et al. Strategies for Feeding the World More Sustainably with Organic Agriculture. Nat. Commun. 2017, 8, 1290. [Google Scholar] [CrossRef] [Green Version]
- Muo, I.; Azeez, A.A. Green entrepreneurship: Literature review and agenda for future research. Int. J. Entrep. Knowl. 2019, 7, 17–29. [Google Scholar] [CrossRef]
- Lage, S.; Gojkovic, Z.; Funk, C.; Gentili, F. Algal Biomass from Wastewater and Flue Gases as a Source of Bioenergy. Energies 2018, 11, 664. [Google Scholar] [CrossRef] [Green Version]
- Maroušek, J.; Maroušková, A. Economic Considerations on Nutrient Utilization in Wastewater Management. Energies 2021, 14, 3468. [Google Scholar] [CrossRef]
- Belay, A. Biology and Industrial Production of Arthrospira (Spirulina). In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 339–358. ISBN 978-1-118-56716-6. [Google Scholar]
- Delrue, F.; Alaux, E.; Moudjaoui, L.; Gaignard, C.; Fleury, G.; Perilhou, A.; Richaud, P.; Petitjean, M.; Sassi, J.-F. Optimization of Arthrospira Platensis (Spirulina) Growth: From Laboratory Scale to Pilot Scale. Fermentation 2017, 3, 59. [Google Scholar] [CrossRef] [Green Version]
- Yuan, D.; Zhan, X.; Wang, M.; Wang, X.; Feng, W.; Gong, Y.; Hu, Q. Biodiversity and Distribution of Microzooplankton in Spirulina (Arthrospira) Platensis Mass Cultures throughout China. Algal Res. 2018, 30, 38–49. [Google Scholar] [CrossRef]
- Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on Novel Foods, Amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and Repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001 (Text with EEA Relevance). Available online: https://eur-lex.europa.eu/legal-content/en/TXT/?uri=CELEX%3A32015R2283 (accessed on 29 July 2021).
- Marzorati, S.; Schievano, A.; Idà, A.; Verotta, L. Carotenoids, Chlorophylls and Phycocyanin from Spirulina: Supercritical CO2 and Water Extraction Methods for Added Value Products Cascade. Green Chem. 2020, 22, 187–196. [Google Scholar] [CrossRef]
- Becker, E.W. Microalgae for Human and Animal Nutrition. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 461–503. ISBN 978-1-118-56716-6. [Google Scholar]
- De la Jara, A.; Ruano-Rodriguez, C.; Polifrone, M.; Assunçao, P.; Brito-Casillas, Y.; Wägner, A.M.; Serra-Majem, L. Impact of Dietary Arthrospira (Spirulina) Biomass Consumption on Human Health: Main Health Targets and Systematic Review. J. Appl. Phycol. 2018, 30, 2403–2423. [Google Scholar] [CrossRef]
- Jourdan, J.P. Manuel de Culture Artisanale de Spiruline; Edition 2 Feb 2018; Antenna Technologies: Geneva, Switzerland, 2018. [Google Scholar]
- Sili, C.; Torzillo, G.; Vonshak, A. Arthrospira (Spirulina). In Ecology of Cyanobacteria II; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 677–705. ISBN 978-94-007-3854-6. [Google Scholar]
- Borowitzka, M.A. Culturing Microalgae in Outdoor Ponds. In Algal Culturing Techniques; Elsevier: Amsterdam, The Netherlands, 2005; pp. 205–218. ISBN 978-0-12-088426-1. [Google Scholar]
- Soni, R.A.; Sudhakar, K.; Rana, R.S. Spirulina—From Growth to Nutritional Product: A Review. Trends Food Sci. Technol. 2017, 69, 157–171. [Google Scholar] [CrossRef] [Green Version]
- Belay, A. Spirulina (Arthrospira): Production and Quality Assurance. In Spirulina in Human Nutrition and Health; Belay, A., Gershwin, M., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 1–25. ISBN 978-1-4200-5256-5. [Google Scholar]
- Richmond, A. Biological Principles of Mass Cultivation of Photoautotrophic Microalgae. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 169–204. ISBN 978-1-118-56716-6. [Google Scholar]
- Grima, E.M.; Acién Fernández, F.G.; Robles Medina, A. Downstream Processing of Cell Mass and Products. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 267–309. ISBN 978-1-118-56716-6. [Google Scholar]
- Guidi, F.; Guerrini, F.; Algeri, M.; Marchiselli, S.; Pinna, A.; Mazzotti, M.; Emiliani, G.; Pistocchi, R. Year-Long Growth of High Quality Spirulina in a Pilot Plant. In Proceedings of the Riunione Annuale Gruppo Algologia, Società Botanica Italiana, Venezia, Italy, 5 November 2015. [Google Scholar]
- Lu, Y.-M.; Xiang, W.-Z.; Wen, Y.-H. Spirulina (Arthrospira) Industry in Inner Mongolia of China: Current Status and Prospects. J. Appl. Phycol. 2011, 23, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Seghiri, R.; Kharbach, M.; Essamri, A. Functional Composition, Nutritional Properties, and Biological Activities of Moroccan Spirulina Microalga. J. Food Qual. 2019, 2019, 3707219. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, Y.; Benemann, J.R.; Zhang, X.; Hu, H.; Qin, S. Microalgal Industry in China: Challenges and Prospects. J. Appl. Phycol. 2016, 28, 715–725. [Google Scholar] [CrossRef]
- Jiménez, C.; Cossío, B.R.; Labella, D.; Xavier Niell, F. The Feasibility of Industrial Production of Spirulina (Arthrospira) in Southern Spain. Aquaculture 2003, 217, 179–190. [Google Scholar] [CrossRef]
- Belay, A. Mass Culture of Spirulina Outdoors—The Earthrise Farms Experience. In Spirulina Platensis (Arthrospira): Physiology, Cell-Biology and Biotechnology; CRC Press: Boca Raton, FL, USA, 1997; p. 252. ISBN 978-0-429-07994-8. [Google Scholar]
- Consejo Insular de Aguas de Gran Canaria. Cartografía temática, Precipitationes. Available online: http://www.aguasgrancanaria.com/cartografia/tematica/precipitaciones.php (accessed on 29 July 2021).
- Sadhwani, J.J.; Veza, J.M. Desalination and Energy Consumption in Canary Islands. Desalination 2008, 221, 143–150. [Google Scholar] [CrossRef]
- Tredici, M.R. Photobiology of Microalgae Mass Cultures: Understanding the Tools for the next Green Revolution. Biofuels 2010, 1, 143–162. [Google Scholar] [CrossRef]
- Dadheech, P.K.; Ballot, A.; Casper, P.; Kotut, K.; Novelo, E.; Lemma, B.; Pröschold, T.; Krienitz, L. Phylogenetic Relationship and Divergence among Planktonic Strains of Arthrospira (Oscillatoriales, Cyanobacteria) of African, Asian and American Origin Deduced by 16S–23S ITS and Phycocyanin Operon Sequences. Phycologia 2010, 49, 361–372. [Google Scholar] [CrossRef]
- Aiba, S.; Ogawa, T. Assessment of Growth Yield of a Blue-Green Alga, Spirulina Platensis, in Axenic and Continuous Culture. J. Gen. Microbiol. 1977, 102, 179–182. [Google Scholar] [CrossRef] [Green Version]
- Schlösser, U.G. SAG—Sammlung von Algenkulturen at the University of Göttingen Catalogue of Strains 1994. Bot. Acta 1994, 107, 113–186. [Google Scholar] [CrossRef]
- Richmond, A.; Grobbelaar, J.U. Factors Affecting the Output Rate of Spirulina Platensis with Reference to Mass Cultivation. Biomass 1986, 10, 253–264. [Google Scholar] [CrossRef]
- Béchet, Q.; Plouviez, M.; Chambonnière, P.; Guieysse, B. Environmental impacts of full-scale algae cultivation. In Microalgae-Based Biofuels and Bioproducts; Elsevier: Amsterdam, The Netherlands, 2017; pp. 505–525. ISBN 978-0-08-101023-5. [Google Scholar]
- Aouir, A.; Amiali, M.; Bitam, A.; Benchabane, A.; Raghavan, V.G. Comparison of the Biochemical Composition of Different Arthrospira Platensis Strains from Algeria, Chad and the USA. J. Food Meas. Charact. 2017, 11, 913–923. [Google Scholar] [CrossRef]
- Gojkovic, Ž.; Vílchez, C.; Torronteras, R.; Vigara, J.; Gómez-Jacinto, V.; Janzer, N.; Gómez-Ariza, J.-L.; Márová, I.; Garbayo, I. Effect of Selenate on Viability and Selenomethionine Accumulation of Chlorella Sorokiniana Grown in Batch Culture. Sci. World J. 2014, 2014, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cuellar-Bermudez, S.P.; Kilimtzidi, E.; Devaere, J.; Goiris, K.; Gonzalez-Fernandez, C.; Wattiez, R.; Muylaert, K. Harvesting of Arthrospira Platensis with Helicoidal and Straight Trichomes Using Filtration and Centrifugation. Sep. Sci. Technol. 2020, 55, 2381–2390. [Google Scholar] [CrossRef]
- Bischoff, H.W.; Bold, H.C. Phycological Studies IV. Some Soil Algae from Enchanted Rock and Related Algal Species; University of Texas Publication No. 6318; University of Texas: Austin, TX, USA, 1963; p. 95. [Google Scholar]
- Giraffa, G.; Rossetti, L.; Neviani, E. An Evaluation of Chelex-Based DNA Purification Protocols for the Typing of Lactic Acid Bacteria. J. Microbiol. Methods 2000, 42, 175–184. [Google Scholar] [CrossRef]
- Diez, B.; Pedros-Alio, C.; Marsh, T.L.; Massana, R. Application of Denaturing Gradient Gel Electrophoresis (DGGE) To Study the Diversity of Marine Picoeukaryotic Assemblages and Comparison of DGGE with Other Molecular Techniques. Appl. Environ. Microbiol. 2001, 67, 2942–2951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goff, L.J.; Moon, D.A.; Coleman, A.W. Molecular delineation of species and species relationships in the red algal Agarophytes Gracilariopsis and Gracilaria (gracilariales). J. Phycol. 1994, 30, 521–537. [Google Scholar] [CrossRef]
- Horwitz, W. Official Methods of Analysis of AOAC International, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- James, C.S. Analytical Chemistry of Foods; Springer: Boston, MA, USA, 1995; ISBN 978-1-4613-5905-0. [Google Scholar]
- Michael, A.; Kyewalyanga, M.S.; Lugomela, C.V. Biomass and Nutritive Value of Spirulina (Arthrospira Fusiformis) Cultivated in a Cost-Effective Medium. Ann. Microbiol. 2019, 69, 1387–1395. [Google Scholar] [CrossRef]
- Boussiba, S.; Richmond, A.E. Isolation and Characterization of Phycocyanins from the Blue-Green Alga Spirulina Platensis. Arch. Microbiol. 1979, 120, 155–159. [Google Scholar] [CrossRef]
- Cunniff, P. Official Methods of Analysis of AOAC International, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1995. [Google Scholar]
- Assaye, H.; Belay, A.; Desse, G.; Gray, D. Seasonal Variation in the Nutrient Profile of Arthrospira Fusiformis Biomass Harvested from an Ethiopian Soda Lake, Lake Chitu. J. Appl. Phycol. 2018, 30, 1597–1606. [Google Scholar] [CrossRef] [Green Version]
- Saleh, A.M.; Dhar, D.W.; Singh, P.K. Comparative Pigment Profiles of Different Spirulina Strains. Res. Biotechnol. 2011, 2, 8. [Google Scholar]
- Commission Regulation (EC) No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs (Text with EEA Relevance). Available online: https://eur-lex.europa.eu/eli/reg/2005/2073 (accessed on 29 July 2021).
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Vonshak, A. (Ed.) Outdoor Mass Production of Spirulina: The Basic Concept. In Spirulina Platensis Arthrospira; CRC Press: Boca Raton, FL, USA, 1997; pp. 97–118. ISBN 978-0-429-07994-8. [Google Scholar]
- Göksan, T.; Zeker, A. The Growth of Spirulina Platensis in Different Culture Systems Under Greenhouse Condition. Turk. J. Biol. 2007, 31, 47–52. [Google Scholar]
- Hidasi, N.; Belay, A. Diurnal Variation of Various Culture and Biochemical Parameters of Arthrospira Platensis in Large-Scale Outdoor Raceway Ponds. Algal Res. 2018, 29, 121–129. [Google Scholar] [CrossRef]
- Ogato, T.; Kifle, D.; Fetahi, T.; Sitotaw, B. Evaluation of Growth and Biomass Production of Arthrospira (Spirulina) Fusiformis in Laboratory Cultures Using Waters from the Ethiopian Soda Lakes Chitu and Shala. J. Appl. Phycol. 2014, 26, 2273–2282. [Google Scholar] [CrossRef]
- Guieysse, B.; Béchet, Q.; Shilton, A. Variability and Uncertainty in Water Demand and Water Footprint Assessments of Fresh Algae Cultivation Based on Case Studies from Five Climatic Regions. Bioresour. Technol. 2013, 128, 317–323. [Google Scholar] [CrossRef]
- Ruiz, J.; Olivieri, G.; de Vree, J.; Bosma, R.; Willems, P.; Reith, J.H.; Eppink, M.H.M.; Kleinegris, D.M.M.; Wijffels, R.H.; Barbosa, M.J. Towards Industrial Products from Microalgae. Energy Environ. Sci. 2016, 9, 3036–3043. [Google Scholar] [CrossRef] [Green Version]
- Duran Quintero, C.; Ventura, A.; Lépine, O.; Pruvost, J. Eco-Design of Spirulina Solar Cultivation: Key Aspects to Reduce Environmental Impacts Using Life Cycle Assessment. J. Clean. Prod. 2021, 299, 126741. [Google Scholar] [CrossRef]
- De Jesus, C.S.; da Silva Uebel, L.; Costa, S.S.; Miranda, A.L.; de Morais, E.G.; de Morais, M.G.; Costa, J.A.V.; Nunes, I.L.; de Souza Ferreira, E.; Druzian, J.I. Outdoor Pilot-Scale Cultivation of Spirulina Sp. LEB-18 in Different Geographic Locations for Evaluating Its Growth and Chemical Composition. Bioresour. Technol. 2018, 256, 86–94. [Google Scholar] [CrossRef]
- Valaskova, K.; Ward, P.; Svabova, L. Deep Learning-Assisted Smart Process Planning, Cognitive Automation, and Industrial Big Data Analytics in Sustainable Cyber-Physical Production Systems. J. Self Gov. Manag. Econ. 2021, 9, 9. [Google Scholar] [CrossRef]
- Kliestik, T.; Nica, E.; Musa, H.; Poliak, M.; Mihai, E.-A. Networked, Smart, and Responsive Devices in Industry 4.0 Manufacturing Systems. Econ. Manag. Financ. Mark. 2020, 15, 23. [Google Scholar] [CrossRef]
- Foissner, W. Schmidingerothrix Extraordinaria Nov. Gen., Nov. Spec., a Secondarily Oligomerized Hypotrich (Ciliophora, Hypotricha, Schmidingerotrichidae Nov. Fam.) from Hypersaline Soils of Africa. Eur. J. Protistol. 2012, 48, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Foissner, W.; Filker, S.; Stoeck, T. Schmidingerothrix Salinarum Nov. Spec. Is the Molecular Sister of the Large Oxytrichid Clade (Ciliophora, Hypotricha). J. Eukaryot. Microbiol. 2014, 61, 61–74. [Google Scholar] [CrossRef]
- Kim, C.-J.; Jung, Y.-H.; Ko, S.-R.; Kim, H.-I.; Park, Y.-H.; Oh, H.-M. Raceway Cultivation of Spirulina Platensis Using Underground Water. J. Microbiol. Biotechnol. 2007, 17, 853–857. [Google Scholar] [PubMed]
- Mendez, C.; Uribe, E. Control of Branchionus Sp. and Amoeba Sp. in Cultures of Arthrospira Sp. Lat. Am. J. Aquat. Res. 2012, 40, 553–561. [Google Scholar] [CrossRef]
- Salbitani, G.; Carfagna, S. Ammonium Utilization in Microalgae: A Sustainable Method for Wastewater Treatment. Sustainability 2021, 13, 956. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, W.; Chen, H.; Zhan, J.; He, C.; Wang, Q. Ammonium Nitrogen Tolerant Chlorella Strain Screening and Its Damaging Effects on Photosynthesis. Front. Microbiol. 2019, 9, 3250. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, D.; Foubert, I.; Fraeye, I.; Meesschaert, B.; Muylaert, K. Flocculation of Chlorella Vulgaris Induced by High PH: Role of Magnesium and Calcium and Practical Implications. Bioresour. Technol. 2012, 105, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Śliwińska-Wilczewska, S.; Wiśniewska, K.; Konarzewska, Z.; Cieszyńska, A.; Barreiro Felpeto, A.; Lewandowska, A.U.; Latała, A. The Current State of Knowledge on Taxonomy, Modulating Factors, Ecological Roles, and Mode of Action of Phytoplankton Allelochemicals. Sci. Total Environ. 2021, 773, 145681. [Google Scholar] [CrossRef] [PubMed]
- El-Sheekh, M.M.; Daboor, S.M.; Swelim, M.A.; Mohamed, S. Production and Characterization of Antimicrobial Active Substance from Spirulina platensis. Iran. J. Microbiol. 2014, 6, 112–119. [Google Scholar]
- Hernandez-Corona, A.; Nieves, I.; Meckes, M.; Chamorro, G.; Barron, B. Antiviral Activity of Spirulina Maxima against Herpes Simplex Virus Type 2. Antivir. Res. 2002, 56, 279–285. [Google Scholar] [CrossRef]
- Templeton, D.W.; Laurens, L.M.L. Nitrogen-to-Protein Conversion Factors Revisited for Applications of Microalgal Biomass Conversion to Food, Feed and Fuel. Algal Res. 2015, 11, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Darwish, R.; Gedi, M.A.; Akepach, P.; Assaye, H.; Zaky, A.S.; Gray, D.A. Chlamydomonas Reinhardtii Is a Potential Food Supplement with the Capacity to Outperform Chlorella and Spirulina. Appl. Sci. 2020, 10, 6736. [Google Scholar] [CrossRef]
- Zaky, A.S.; Pensupa, N.; Andrade-Eiroa, Á.; Tucker, G.A.; Du, C. A New HPLC Method for Simultaneously Measuring Chloride, Sugars, Organic Acids and Alcohols in Food Samples. J. Food Compos. Anal. 2017, 56, 25–33. [Google Scholar] [CrossRef]
- Koru, E. Earth Food Spirulina (Arthrospira): Production and Quality Standarts. In Food Additive; El-Samragy, Y., Ed.; IntechOpen: London, UK, 2012; ISBN 978-953-51-0067-6. [Google Scholar]
- Tatsuzawa, H.; Takizawa, E.; Wada, M.; Yamamoto, Y. Fatty Acid and Lipid Composition of the Acidophilic Green Alga Chlamydomonas Sp. J. Phycol. 1996, 32, 598–601. [Google Scholar] [CrossRef]
- Norici, A.; Bazzoni, A.M.; Pugnetti, A.; Raven, J.A.; Giordano, M. Impact of Irradiance on the C Allocation in the Coastal Marine Diatom Skeletonema marinoi Sarno and Zingone. Plant Cell Environ. 2011, 34, 1666–1677. [Google Scholar] [CrossRef] [PubMed]
- Johnson, X.; Alric, J. Central Carbon Metabolism and Electron Transport in Chlamydomonas Reinhardtii: Metabolic Constraints for Carbon Partitioning between Oil and Starch. Eukaryot. Cell 2013, 12, 776–793. [Google Scholar] [CrossRef] [Green Version]
- Markou, G.; Chatzipavlidis, I.; Georgakakis, D. Effects of Phosphorus Concentration and Light Intensity on the Biomass Composition of Arthrospira (Spirulina) Platensis. World J. Microbiol. Biotechnol. 2012, 28, 2661–2670. [Google Scholar] [CrossRef]
- Markou, G.; Chatzipavlidis, I.; Georgakakis, D. Carbohydrates Production and Bio-Flocculation Characteristics in Cultures of Arthrospira (Spirulina) Platensis: Improvements Through Phosphorus Limitation Process. BioEnergy Res. 2012, 5, 915–925. [Google Scholar] [CrossRef]
- Markou, G.; Angelidaki, I.; Georgakakis, D. Microalgal Carbohydrates: An Overview of the Factors Influencing Carbohydrates Production, and of Main Bioconversion Technologies for Production of Biofuels. Appl. Microbiol. Biotechnol. 2012, 96, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Yaakob, M.A.; Mohamed, R.M.S.R.; Al-Gheethi, A.; Aswathnarayana Gokare, R.; Ambati, R.R. Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview. Cells 2021, 10, 393. [Google Scholar] [CrossRef]
- Ballicora, M.A.; Iglesias, A.A.; Preiss, J. ADP-Glucose Pyrophosphorylase: A Regulatory Enzyme for Plant Starch Synthesis. Photosynth. Res. 2004, 79, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Batista, A.P.; Gouveia, L.; Bandarra, N.M.; Franco, J.M.; Raymundo, A. Comparison of Microalgal Biomass Profiles as Novel Functional Ingredient for Food Products. Algal Res. 2013, 2, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Calvo, J.J.; Mazuelos, C.; Hermosin, B.; Saiz-Jimenez, C. Chemical Composition of Spirulina and Eukaryotic Algae Food Products Marketed in Spain. J. Appl. Phycol. 1993, 5, 425–435. [Google Scholar] [CrossRef]
- Gojkovic, Ž.; Guidi, F.; Bustamante, B.; Venuleo, M.; Assuncao, P.; Portillo, E. Scaling-up and Semi-Continuous Cultivation of Locally Isolated Marine Microalgae Tetraselmis striata in the Tropical Region of Gran Canary Island. Processes 2021, 9, 1326. [Google Scholar] [CrossRef]
- Campanella, L.; Crescentini, G.; Avino, P. Chemical Composition and Nutritional Evaluation of Some Natural and Commercial Food Productsbased on Spirulina. Analusis 1999, 27, 533–540. [Google Scholar] [CrossRef]
- Pignolet, O.; Jubeau, S.; Vaca-Garcia, C.; Michaud, P. Highly Valuable Microalgae: Biochemical and Topological Aspects. J. Ind. Microbiol. Biotechnol. 2013, 40, 781–796. [Google Scholar] [CrossRef] [Green Version]
- Lauceri, R.; Bresciani, M.; Lami, A.; Morabito, G. Chlorophyll a Interference in Phycocyanin and Allophycocyanin Spectrophotometric Quantification. J. Limnol. 2018, 77, 169–177. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Guideline: Potassium Intake for Adults and Children; WHO: Geneva, Switzerland, 2012; ISBN 978-92-4-150482-9. [Google Scholar]
- World Health Organization. Guideline: Sodium Intake for Adults and Children; World Health Organization: Geneva, Switzerland, 2012; ISBN 978-92-4-150483-6. [Google Scholar]
- Commission Regulation (EC) No 629/2008 of 2 July 2008 Amending Regulation (EC) No 1881/2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs (Text with EEA Relevance). Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32008R0629 (accessed on 29 July 2021).
- Lopes, B.C.; Machado, E.C.; Rodrigues, H.F.; Leal, C.D.; de Araújo, J.C.; Teixeira de Matos, A. Effect of Alkaline Treatment on Pathogens, Bacterial Community and Antibiotic Resistance Genes in Different Sewage Sludges for Potential Agriculture Use. Environ. Technol. 2020, 41, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Akbarnezhad, M.; Shamsaie Mehrgan, M.; Kamali, A.; Javaheri Baboli, M. Effects of Microelements (Fe, Cu, Zn) on Growth and Pigment Contents of Arthrospira (Spirulina) Platensis. Iran. J. Fish. Sci. 2019, 19, 653–668. [Google Scholar] [CrossRef]
- Balaji, S.; Kalaivani, T.; Rajasekaran, C. Biosorption of Zinc and Nickel and Its Effect on Growth of Different Spirulina Strains: Biosorption Potentials of Spirulina Strains. CLEAN Soil Air Water 2014, 42, 507–512. [Google Scholar] [CrossRef]
- Nalimova, A.A.; Popova, V.V.; Tsoglin, L.N.; Pronina, N.A. The Effects of Copper and Zinc on Spirulina Platensis Growth and Heavy Metal Accumulation in Its Cells. Russ. J. Plant Physiol. 2005, 52, 229–234. [Google Scholar] [CrossRef]



| Chemical | Spirulina Medium 1 (g/L) | Medium OUT 2 (g/L) | Nutrient Replenishment 3 (g/kg of Algal DW) |
|---|---|---|---|
| NaHCO3 | 13.61 | 8 | - |
| Na2CO3 | 4.03 | - | - |
| NaNO3 | 2.5 | - | - |
| KNO3 | - | 2 | 1000 |
| K2HPO4 | 0.5 | - | - |
| NH4H2PO4 | - | 0.06 | 50 |
| NaCl | 1 | 5 | - |
| K2SO4 | 1 | - | - |
| CaCl2·2H2O | 0.04 | - | - |
| MgSO4·7H2O | 0.2 | 0.16 | 30 |
| FeSO4·7H2O | 0.01 | 0.005 | 2.5 |
| EDTA | 0.084 | - | - |
| CO(NH2)2 | - | 0.015 | 15 |
| H3BO3 | 0.00286 | - | - |
| MnCl2·4H2O | 0.00181 | - | - |
| ZnSO4·7H2O | 0.0022 | - | - |
| Na2MoO4·2H2O | 0.00039 | - | - |
| CuSO4·5H2O | 0.00008 | - | - |
| CO(NO3)2·6H2O | 0.00005 | - | - |
| Cycle | Time (Days) | T (°C) | pH | Salinity (g/L) | ER (mm/Day) | Cx Start (g/L DW) | Cx End (g/L DW) | Pvol (g/L/Day−1) | |
|---|---|---|---|---|---|---|---|---|---|
| F phase | 1 | 0–6 | 24.9 ± 0.3 | 10.09 ± 0.38 | 12.7 ± 0.5 | 3.5 ± 1.3 | 0.17 ± 0.008 | 0.60 ± 0.010 | 0.08 ± 0.002 |
| 2 | 6–13 | 24.8 ± 0.8 | 9.61 ± 0.19 | 12.3 ± 0.7 | 4.3 ± 0.6 | 0.37 ± 0.002 | 0.86 ± 0.017 | 0.07 ± 0.002 | |
| 3 | 13–20 | 25.2 ± 0.2 | 9.99 ± 0.26 | 12.3 ± 0.7 | 3.3 ± 0.9 | 0.69 ± 0.019 | 1.04 ± 0.012 | 0.05 ± 0.002 | |
| 4 | 20–29 | 24.8 ± 1.5 | 10.26 ± 0.33 | 13.8 ± 0.8 | 4.5 ± 1.0 | 0.88 ± 0.007 | 1.26 ± 0.043 | 0.04 ± 0.003 | |
| R phase | 5 | 29–33 | 23.5 ± 1.1 | 10.75 ± 0.04 | 12.4 ± 0.9 | 4.8 ± 0.9 | 1.11 ± 0.011 | 1.25 ± 0.011 | 0.04 ± 0.002 |
| 6 | 33–35 | 23.0 ± 0.9 | 10.33 ± 0.13 | 11.9 ± 0.4 | 4.0 ± 1.1 | 0.93 ± 0.008 | 1.03 ± 0.036 | 0.06 ± 0.004 | |
| 7 | 35–39 | 24.6 ± 1.6 | 10.57 ± 0.10 | 12.0 ± 0.7 | 4.3 ± 0.7 | 0.64 ± 0.013 | 0.87 ± 0.018 | 0.06 ± 0.001 | |
| 8 | 39–42 | 23.1 ± 1.3 | 10.39 ± 0.24 | 11.7 ± 0.3 | 3.1 ± 1.0 | 0.63 ± 0.017 | 0.78 ± 0.021 | 0.06 ± 0.002 | |
| 9 | 42–46 | 24.9 ± 2.2 | 10.51 ± 0.08 | 11.7 ± 0.2 | 2.8 ± 0.6 | 0.62 ± 0.009 | 0.88 ± 0.011 | 0.07 ± 0.002 | |
| 10 | 46–49 | 25.0 ± 2.6 | 10.50 ± 0.10 | 11.7 ± 0.2 | 2.4 ± 1.5 | 0.60 ± 0.016 | 0.79 ± 0.026 | 0.06 ± 0.005 | |
| 11 | 49–54 | 23.9 ± 0.5 | 10.68 ± 0.06 | 12.0 ± 0.4 | 2.0 ± 1.2 | 0.63 ± 0.020 | 0.89 ± 0.008 | 0.06 ± 0.002 | |
| 12 | 54–60 | 23.7 ± 0.4 | 10.69 ± 0.05 | 11.6 ± 0.4 | 2.5 ± 0.7 | 0.61 ± 0.028 | 0.90 ± 0.037 | 0.05 ± 0.002 | |
| Cumul. F | 1–4 | 0–29 | 24.9 ± 0.9 | 9.97 ± 0.38 * | 12.8 ± 0.9 * | 3.9 ± 1.0 * | 0.53 ± 0.32 | 0.94 ± 0.28 | 0.06 ± 0.016 |
| Cumul. R | 5–12 | 29–60 | 24.2 ± 1.6 | 10.56 ± 0.16 * | 11.9 ± 0.5 * | 3.0 ± 1.1 * | 0.72 ± 0.19 | 0.93 ± 0.15 | 0.06 ± 0.010 |
| Overall | 1–12 | 0–60 | 24.6 ± 1.3 | 10.25 ± 0.42 | 12.3 ± 0.8 | 3.5 ± 1.2 | 0.66 ± 0.25 | 0.93 ± 0.19 | 0.06 ± 0.012 |
| Elements | F Phase | R Phase | Overall | |
|---|---|---|---|---|
| Minerals (mg/100 g) | K | 1704.4 ± 636.0 | 1839.2 ± 894.9 | 1771.8 ± 698.3 |
| Na | 1580.8 ± 865.0 | 1490.7 ± 347.8 | 1535.7 ± 591.7 | |
| P | 542.1 ± 11.6 | 472.3 ± 56.0 | 507.2 ± 52.6 | |
| Mg | 163.0 ± 38.3 | 146.0 ± 5.3 | 154.5 ± 26.2 | |
| Ca | 43.7 ± 11.3 | 46.7 ± 11.5 | 45.2 ± 10.3 | |
| Trace elements (mg/100 g) | Fe | 47.7 ± 8.3 | 81.0 ± 25.6 | 64.4 ± 24.9 |
| Mn | 0.7 ± 0.2 | 0.8 ± 0.2 | 0.8 ± 0.2 | |
| Cu | 0.9 ± 0.1 | 0.8 ± 0.1 | 0.8 ± 0.1 | |
| Zn | 2.4 ± 0.6 | 2.3 ± 0.5 | 2.4 ± 0.5 | |
| B | 2.4 ± 1.6 | 2.5 ± 0.4 | 2.5 ± 1.1 | |
| Heavy metals (mg/kg) | Se | 0.007 ± 0.010 | 0.007 ± 0.008 | 0.007 ± 0.008 |
| Cr | 0.012 ± 0.008 | 0.010 ± 0.005 | 0.011 ± 0.006 | |
| Pb | 0.023 ± 0.016 | 0.026 ± 0.024 | 0.024 ± 0.018 | |
| Cd | 0.014 ± 0.025 | 0.007 ± 0.012 | 0.011 ± 0.018 | |
| Mo | 0.024 ± 0.032 | nd | 0.012 ± 0.027 | |
| Ni, Co, As, Hg | nd | nd | nd | |
| Microbiological Parameters | t0 | F1 | F2 | F3 | R1 | R2 | R3 |
|---|---|---|---|---|---|---|---|
| Total aerobic mesophilic flora (cfu/g) | 2.6 × 103 | 1.1 × 103 | 1.6 × 103 | 7.4 × 103 | 2.0 × 103 | 2.3 × 103 | 8.9 × 103 |
| Yeasts and molds (cfu/g) | <20 | <20 | <20 | ||||
| Enterobacteriaceae (cfu/g) | <100 | <100 | <100 | ||||
| Total coliforms (cfu/g) | <10 | <10 | <10 | ||||
| Escherichia coli (cfu/g) | <10 | <10 | <10 | ||||
| Staphylococcus spp. (cfu/g) | <10 | <10 | <10 | ||||
| Clostridium perfrigens (cfu/g) | <10 | <10 | <10 | ||||
| Salmonella spp. (Abs/25g) | nd | nd | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guidi, F.; Gojkovic, Z.; Venuleo, M.; Assunçao, P.A.C.J.; Portillo, E. Long-Term Cultivation of a Native Arthrospira platensis (Spirulina) Strain in Pozo Izquierdo (Gran Canaria, Spain): Technical Evidence for a Viable Production of Food-Grade Biomass. Processes 2021, 9, 1333. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9081333
Guidi F, Gojkovic Z, Venuleo M, Assunçao PACJ, Portillo E. Long-Term Cultivation of a Native Arthrospira platensis (Spirulina) Strain in Pozo Izquierdo (Gran Canaria, Spain): Technical Evidence for a Viable Production of Food-Grade Biomass. Processes. 2021; 9(8):1333. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9081333
Chicago/Turabian StyleGuidi, Flavio, Zivan Gojkovic, Marianna Venuleo, Patrícia Alexandra Clemente Janeiro Assunçao, and Eduardo Portillo. 2021. "Long-Term Cultivation of a Native Arthrospira platensis (Spirulina) Strain in Pozo Izquierdo (Gran Canaria, Spain): Technical Evidence for a Viable Production of Food-Grade Biomass" Processes 9, no. 8: 1333. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9081333