
The Potential of Micromammals for the Stratigraphy and the Timing of Human Occupations at La Roche-à-Pierrot (Saint-Césaire, France)

 , , , ,
, , , ,
Abstract
:1. Introduction
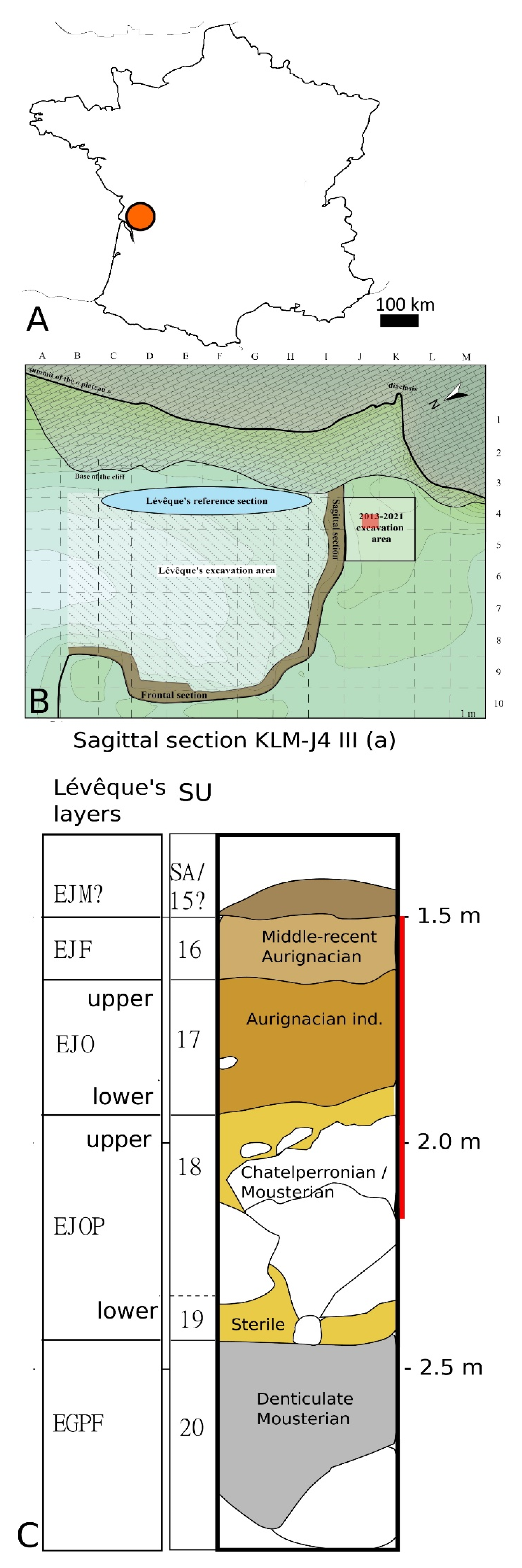
2. Site Presentation
3. Material and Methods
4. Results
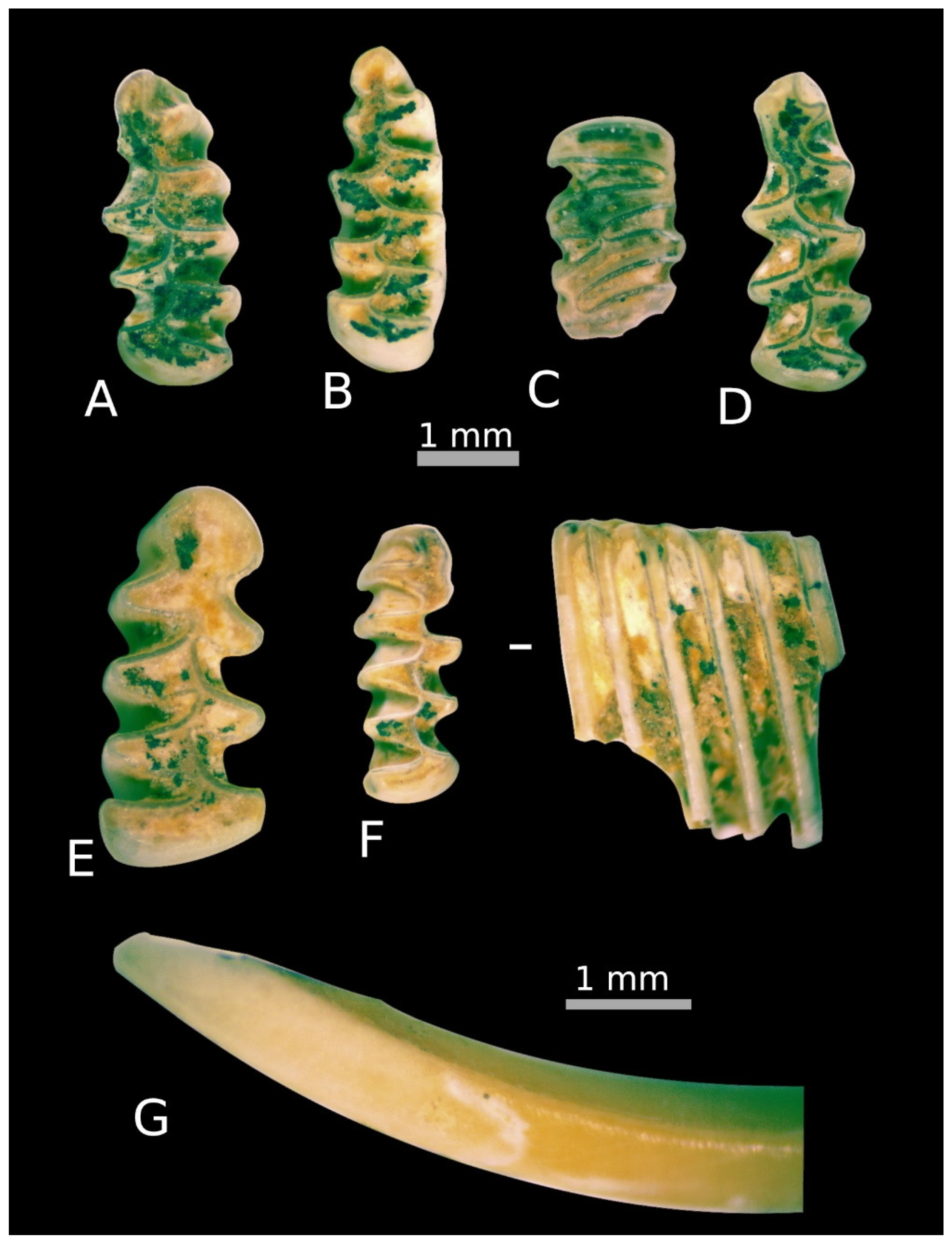
4.1. Taxonomic Composition
4.2. Origin of the Micromammal Accumulation
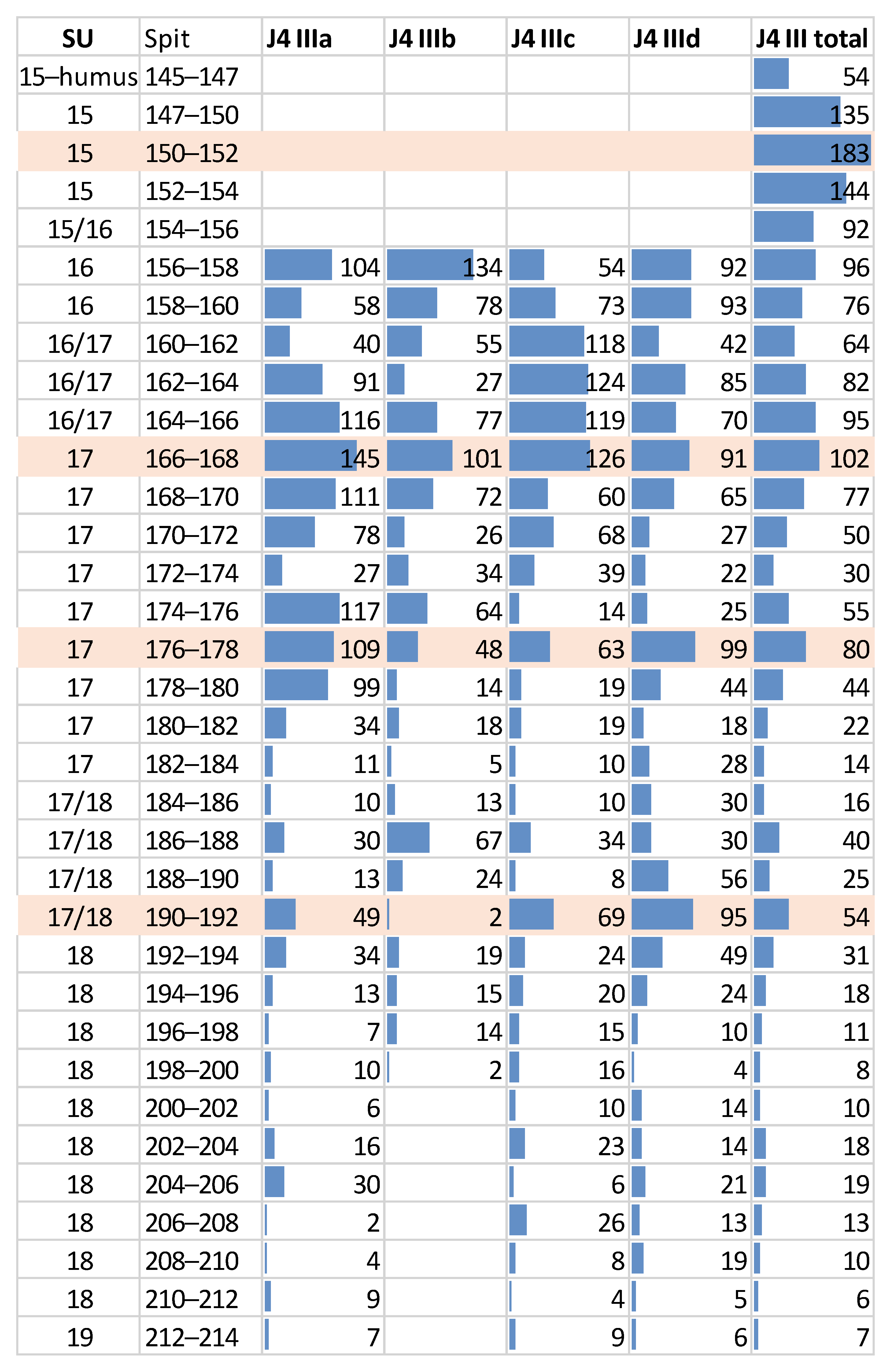
4.3. Density Variation
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Royer, A.; Lécuyer, C.; Montuire, S.; Escarguel, G.; Fourel, F.; Mann, A.; Maureille, B. Late Pleistocene (MIS 3-4) climate inferred from micromammal communities and δ18O of rodents from Les Pradelles, France. Quat. Res. 2013, 80, 113–124. [Google Scholar] [CrossRef]
- Lebreton, L.; Desclaux, E.; Hanquet, C.; Moigne, A.-M.; Perrenoud, C. Environmental context of the Caune de l’Arago Acheulean occupations (Tautavel, France), new insights from microvertebrates in Q-R levels. Quat. Int. 2016, 411, 182–192. [Google Scholar] [CrossRef]
- Stoetzel, E.; Montuire, S. Les rongeurs, indicateurs des paléoclimats: Application aux assemblages de trois sites du nord de la France. Quaternaire 2016, 27, 227–238. [Google Scholar] [CrossRef] [Green Version]
- López-García, J.M.; Blain, H.A.; Sánchez-Bandera, C.; Cohen, J.; Lebreton, L.; Montuire, S.; Stewart, J.R.; Desclaux, E. Multi-method approach using small vertebrate assemblages to reconstruct the Marine Isotope Stage 6 climate and environment of the Lazaret cave sequence (Maritime Alps, Nice, France). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 577, 110529. [Google Scholar] [CrossRef]
- Piñero, P.; Agustí, J.; Blain, H.A.; Laplana, C. Paleoenvironmental reconstruction of the Early Pleistocene site of Quibas (SE Spain) using a rodent assemblage. C. R. Palevol 2016, 15, 659–668. [Google Scholar] [CrossRef]
- Denys, C. Nouveaux critères de reconnaissance des concentrations de microvertébrés d’après l’étude des pelotes de chouettes du Botswana (Afrique australe). Bulletin du Muséum National d’Histoire Naturelle 1985, 7, 879–933. [Google Scholar]
- Andrews, P. Owls, Caves and Fossils: Predation, Preservation, and Accumulation of Small Mammal Bones in Caves, with an Analysis of the Pleistocene Cave Faunas from Westbury-Sub-Mendip; University of Chicago Press: Somerset, UK, 1990; 231p. [Google Scholar]
- Fernández-Jalvo, Y.; Andrews, P. Small Mammal Taphonomy of Gran Dolina, Atapuerca (Burgos), Spain. J. Archaeol. Sci. 1992, 19, 407–428. [Google Scholar] [CrossRef]
- Fernández-Jalvo, Y.; Denys, C.; Andrews, P.; Williams, T.W.; Dauphin, Y.; Humphrey, L. Taphonomy and palaeoecology of Olduvai Bed-I (Pleistocene, Tanzania). J. Hum. Evol. 1998, 34, 137–172. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Jalvo, Y.; Andrews, P.; Denys, C.; Sese, C.; Stoetzel, E.; Marin Monfort, D.; Pequero, D. Taphonomy for taxonomists, implications in small mammal studies. Quat. Sci. Rev. 2016, 139, 138–157. [Google Scholar] [CrossRef]
- Denys, C. Des référentiels en taphonomie des petits vertébrés: Bilan et perspectives. In Taphonomie des Petits Vertébrés: Référentiels et Transferts Aux Fossiles; Laroulandie, V., Mallye, J.-B., Denys, C., Eds.; British Archaeological Reports; International Series 2269; Archaeopress: Oxford, UK, 2011; pp. 7–22. [Google Scholar]
- Rofes, J.; Ordiales, A.; Iriarte, E.; Cuenca-Bescós, G.; Galindo-Pellicena, M.Á.; Pérez-Romero, A.; Carretero, J.M.; Arsuaga, J.L. Human Activities, Biostratigraphy and Past Environment Revealed by Small-Mammal Associations at the Chalcolithic Levels of El Portalón de Cueva Mayor (Atapuerca, Spain). Quaternary 2021, 4, 16. [Google Scholar] [CrossRef]
- Royer, A. Intérêts de l’utilisation des décapages lors des fouilles archéologiques pour l’étude des restes de petits vertébrés. PALEO 2014, 25, 277–286. [Google Scholar] [CrossRef]
- Laroulandie, V.; Royer, A.; Mallye, J.-B.; Bailon, S.; Danger, M. Les petits vertébrés de Peyrazet et les chasseurs-collecteurs du Magdalénien supérieur. In La Grotte-Abri de Peyrazet (Creysse, Lot, France) au Magdalénien. Originalité fonctionnelle d’un Habitat des Derniers Chasseurs de Rennes du Quercy; Langlais, M., Laroulandie, V., Eds.; Supplément à Gallia Préhistoire 43; CNRS: Paris, France, 2021; pp. 141–152. [Google Scholar]
- Rambaud, D.; Laroulandie, V.; Primault, J.; Béarez, P. Les poissons et les oiseaux du Taillis des Coteaux (Antigny, Vienne), niveaux magdaléniens: Origine naturelle ou culturelle? In Taphonomie des Petits Vertébrés: Référentiels et Transferts Aux Fossiles; Laroulandie, V., Mallye, J.-B., Denys, C., Eds.; British Archaeological Reports; International Series 2269; Archaeopress: Oxford, UK, 2011; pp. 167–179. [Google Scholar]
- Richardson, C.; Miller, C. Recommendations for Protecting Raptors from Human Disturbance: A Review. Wildl. Soc. Bull. (1973–2006) 1997, 25, 634–638. [Google Scholar]
- Martínez-Abraín, A.; Oro, D.; Jiménez, J.; Stewart, G.; Pullin, A. A systematic review of the effects of recreational activities on nesting birds of prey. Basic Appl. Ecol. 2010, 11, 312–319. [Google Scholar] [CrossRef]
- Lévêque, F.; Vandermeersch, B. Découverte de restes humains dans un niveau castelperronien à Saint-Césaire (Charente-Maritime). C. R. l’Acad. Sci. Paris Sér. D 1980, 291, 187–189. [Google Scholar]
- Gravina, B.; Bachellerie, F.; Caux, S.; Discamps, E.; Faivre, J.-P.; Galland, A.; Michel, A.; Teyssandier, N.; Bordes, J.-G. No reliable evidence for a Neanderthal-Châtelperronian association at La Roche-à-Pierrot, Saint-Césaire. Sci. Rep. 2018, 8, 15134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, E. Reassessing Paleolithic Subsistence: The Neandertal and Modern Human Foragers of Saint-Césaire; Cambridge University Press: New York, NY, USA, 2012; 358p. [Google Scholar]
- Lévêque, F.; Backer, A.M.; Guilbaud, M. (Eds.) Context of a Late Neandertal. Implications of Multidisciplinary Research for the Transition to Upper Paleolithic Adaptations at Saint-Césaire, Charente-Maritime, France; Monographs in World Archaeology 16; Prehistory Press: Madison, WI, USA, 1993; 131p. [Google Scholar]
- Marquet, J.-C. Paléoenvironnement et Chronologie des Sites du Domaine Atlantique Français d’âge Pléistocène Moyen et Supérieur D’après L’étude des Rongeurs; Les Cahiers de la Claise, Supplément 2: Tours, France, 1993; 330p. [Google Scholar]
- Crevecoeur, I. Reprise des fouilles à La Roche-à-Pierrot, Saint-Césaire. In Le troisième Homme. Préhistoire de l’Altaï; Cleyet-Merle, J.-J., Derevianko, A.P., Geneste, J.-M., Gravina, B., Krivoshapkin, A.I., Maureille, B., Turq, A., Shunkov, M.V., Slimak, L., Eds.; Réunion des Musées Nationaux-Grand Palais: Paris, France, 2017; p. 107. [Google Scholar]
- Dodson, P.; Wexlar, D. Taphonomic investigations of owl pellets. Paleobiology 1979, 5, 275–284. [Google Scholar] [CrossRef]
- Morin, E. Evidence for declines in human population densities during the early Upper Paleolithic in western Europe. Proc. Natl. Acad. Sci. USA 2008, 105, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.L.; Bowman, N. Diet and ecology of Short-eared Owls Asio flammeus breeding on heather moor. Bird Study 1986, 33, 12–17. [Google Scholar] [CrossRef]
- Martínez, J.A.; Zuberogoitia, I. Habitat preferences for Long-eared Owls Asio otus and Little Owls Athene noctua in semi-arid environments at three spatial scales. Bird Study 2004, 51, 163–169. [Google Scholar] [CrossRef]
- Slon, V.; Hopfe, C.; Weiß, C.L.; Mafessoni, F.; de la Rasilla, M.; Lalueza-Fox, C.; Rosas, A.; Soressi, M.; Knul, M.V.; Miller, R.; et al. Neandertal and Denisovan DNA from Pleistocene sediments. Science 2017, 356, 605–608. [Google Scholar] [CrossRef] [Green Version]
- Rohland, N.; Glocke, I.; Aximu-Petri, A.; Meyer, M. Extraction of highly degraded DNA from ancient bones, teeth and sediments for high-throughput sequencing. Nat. Protoc. 2018, 13, 2447–2461. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/SU | 15-Humus | 15 | 16 | 16/17 | 17 | 17/18 | 18 | 18/19 | 19 |
|---|---|---|---|---|---|---|---|---|---|
| Microtus arvalis/agrestis (Common/field vole) | 5 | 63 | 33 | 77 | 106 | 26 | 28 | 3 | 1 |
| Lasiopodomys gregalis (Narrow-headed vole) | 7 | 74 | 40 | 42 | 109 | 32 | 30 | 4 | 2 |
| Arvicola amphibius (European water vole) | 0 | 5 | 0 | 0 | 8 | 1 | 1 | 1 | 1 |
| Microtus oeconomus (Root vole) | 0 | 4 | 0 | 0 | 2 | 0 | 1 | 0 | 0 |
| Lemmus/Myopus sp. (Lemming) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Spermophilus sp. (Ground squirrel) | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 |
| Sorex sp. (Shrew) | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 |
| Talpa sp. (Mole) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| SU | Incisors | Molars | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | N Total | 0 | 1 | 2 | 3 | N Total | |||||||||
| N | % | N | % | N | % | N | % | N | % | N | % | N | % | N | % | |||
| 15 | 82 | 92 | 5 | 6 | 2 | 2 | 0 | 0 | 89 | 62 | 97 | 1 | 2 | 0 | 0 | 1 | 2 | 64 |
| 16 | 83 | 89 | 6 | 6 | 1 | 1 | 3 | 3 | 93 | 104 | 96 | 0 | 0 | 1 | 1 | 3 | 3 | 108 |
| 16/17 | 95 | 95 | 5 | 5 | 0 | 0 | 0 | 0 | 100 | 177 | 95 | 7 | 4 | 0 | 0 | 3 | 2 | 187 |
| 17 | 251 | 93 | 18 | 7 | 0 | 0 | 0 | 0 | 269 | 374 | 95 | 12 | 3 | 5 | 1 | 3 | 1 | 394 |
| 17/18 | 66 | 94 | 4 | 6 | 0 | 0 | 0 | 0 | 70 | 92 | 95 | 4 | 4 | 0 | 0 | 1 | 1 | 97 |
| 18 | 57 | 92 | 3 | 5 | 2 | 3 | 0 | 0 | 62 | 94 | 90 | 5 | 5 | 2 | 2 | 3 | 3 | 104 |
| 18/19 | 1 | _ | 0 | _ | 0 | _ | 0 | _ | 1 | 6 | _ | 1 | _ | 1 | _ | 0 | _ | 8 |
| 19 | 0 | _ | 0 | _ | 0 | _ | 0 | _ | 0 | 2 | _ | 1 | _ | 2 | _ | 0 | _ | 5 |
| Density | MNE Micro | N Fauna J4 III | N Fauna J4 III-IV | N Lithics J4 III | N Lithics J4 III-IV | W Fauna | W Lithics | |
|---|---|---|---|---|---|---|---|---|
| Density | 1.97 × 10−10 | 0.681 | 0.040 | 0.091 | 0.002 | 0.604 | 0.717 | |
| MNE Micro | 0.864 | 0.866 | 0.003 | 0.019 | 3.79 × 10−5 | 0.234 | 0.095 | |
| N Fauna J4 III | 0.076 | 0.031 | 8.55 × 10−5 | 0.016 | 0.789 | 0.039 | 0.328 | |
| N Fauna J4 III-IV | −0.364 | −0.502 | 0.638 | 0.001 | 3.62 × 10−5 | 0.0002 | 0.002 | |
| N Lithics J4 III | −0.303 | −0.412 | 0.423 | 0.552 | 0.001 | 0.017 | 0.088 | |
| N Lithics J4 III-IV | −0.521 | −0.661 | −0.049 | 0.662 | 0.556 | 0.012 | 0.002 | |
| W Fauna | 0.095 | −0.216 | 0.366 | 0.621 | 0.417 | 0.441 | 8.17 × 10−13 | |
| W Lithics | 0.067 | −0.300 | 0.179 | 0.531 | 0.306 | 0.530 | 0.907 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebreton, L.; Morin, E.; Gravina, B.; Michel, A.; Bachellerie, F.; Beauval, C.; Flas, D.; Laroulandie, V.; Marot, J.; Rougier, H.; et al. The Potential of Micromammals for the Stratigraphy and the Timing of Human Occupations at La Roche-à-Pierrot (Saint-Césaire, France). Quaternary 2021, 4, 33. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040033
Lebreton L, Morin E, Gravina B, Michel A, Bachellerie F, Beauval C, Flas D, Laroulandie V, Marot J, Rougier H, et al. The Potential of Micromammals for the Stratigraphy and the Timing of Human Occupations at La Roche-à-Pierrot (Saint-Césaire, France). Quaternary. 2021; 4(4):33. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040033
Chicago/Turabian StyleLebreton, Loïc, Eugène Morin, Brad Gravina, Alexandre Michel, François Bachellerie, Cédric Beauval, Damien Flas, Véronique Laroulandie, Josserand Marot, Hélène Rougier, and et al. 2021. "The Potential of Micromammals for the Stratigraphy and the Timing of Human Occupations at La Roche-à-Pierrot (Saint-Césaire, France)" Quaternary 4, no. 4: 33. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4040033