Drought-Induced Reduction in Net Primary Productivity across Mainland China from 1982 to 2015

 ,
,  , ,
, ,
Abstract
:
1. Introduction
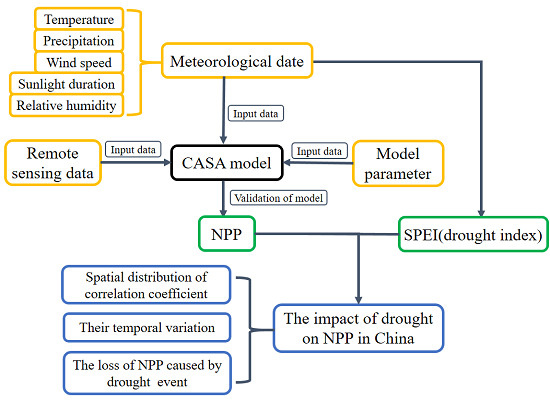
2. Data and Method
2.1. Method
2.1.1. Carnegie Ames Stanford Application (CASA) Model
2.1.2. SPEI
2.1.3. Mann–Kendall Analysis
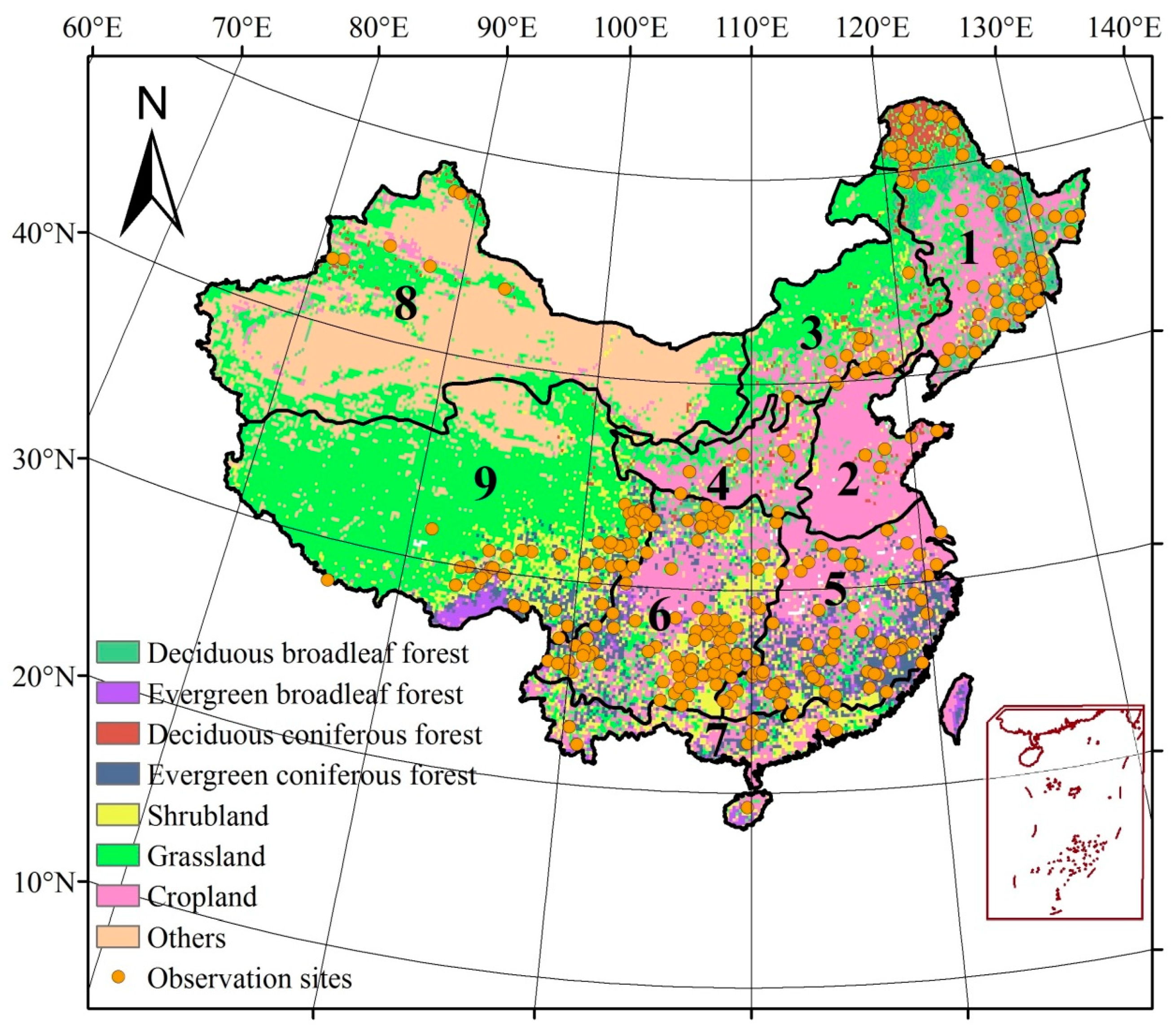
2.2. Data
3. Results
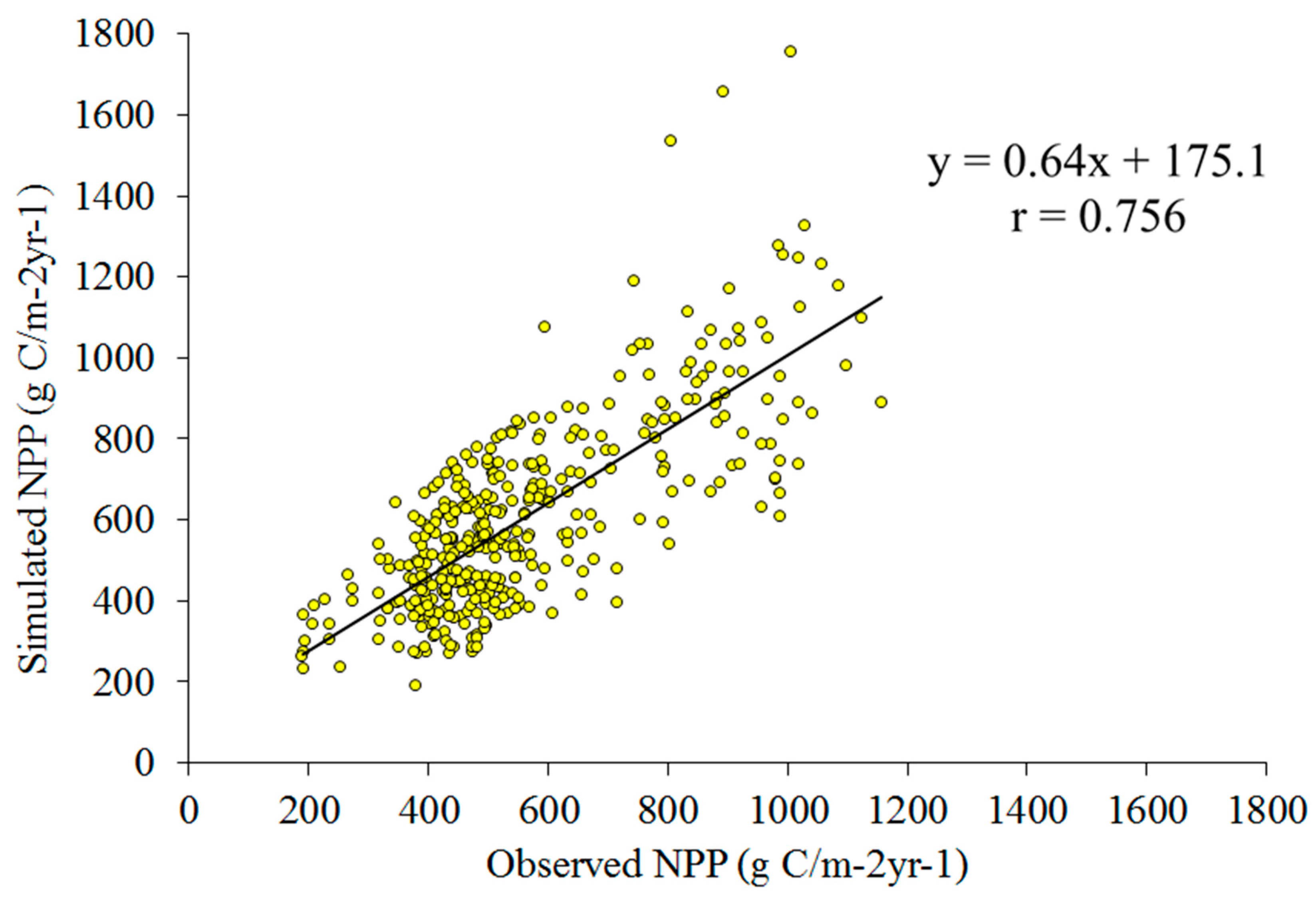
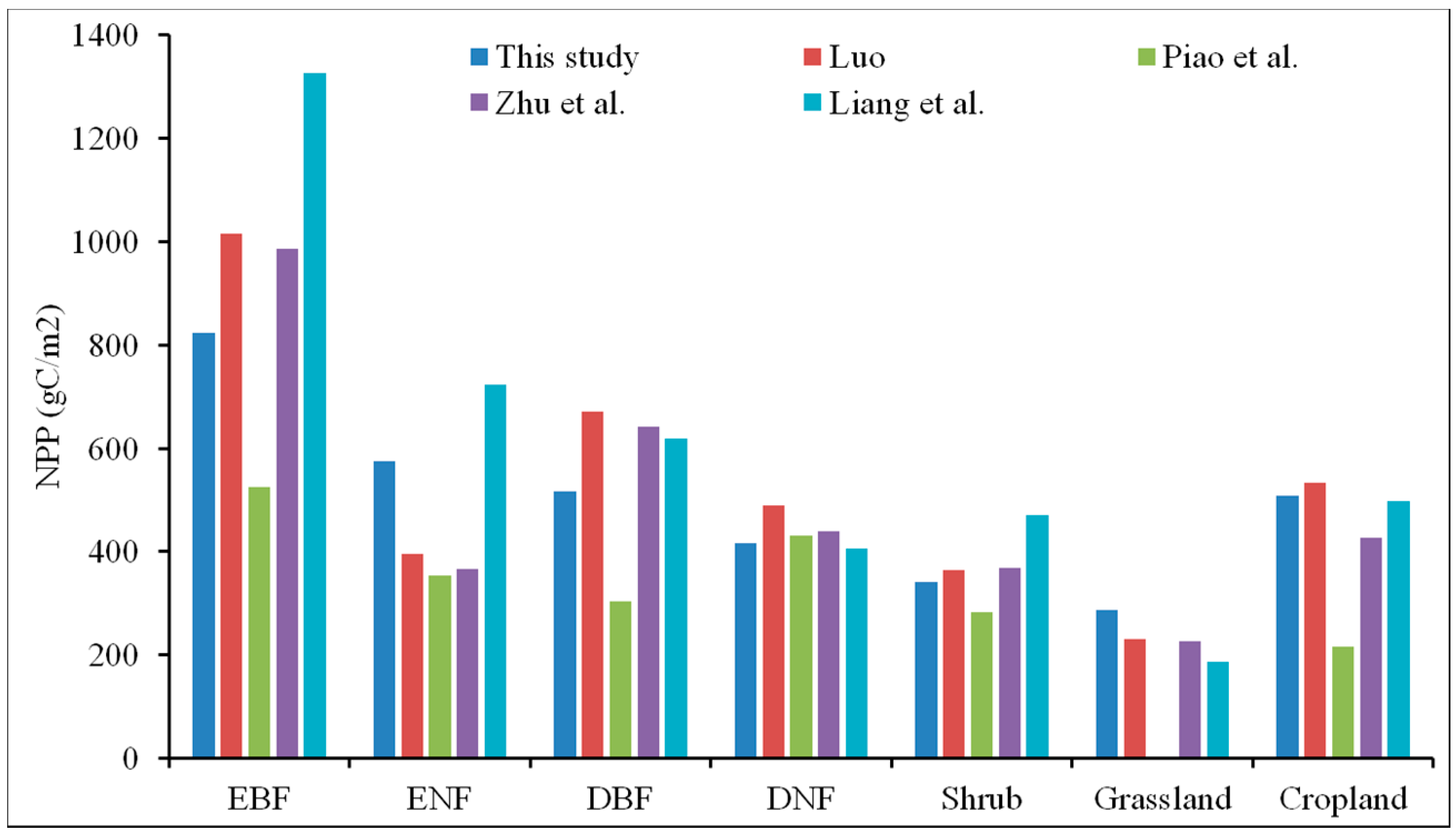
3.1. Validation of the NPP Calculation
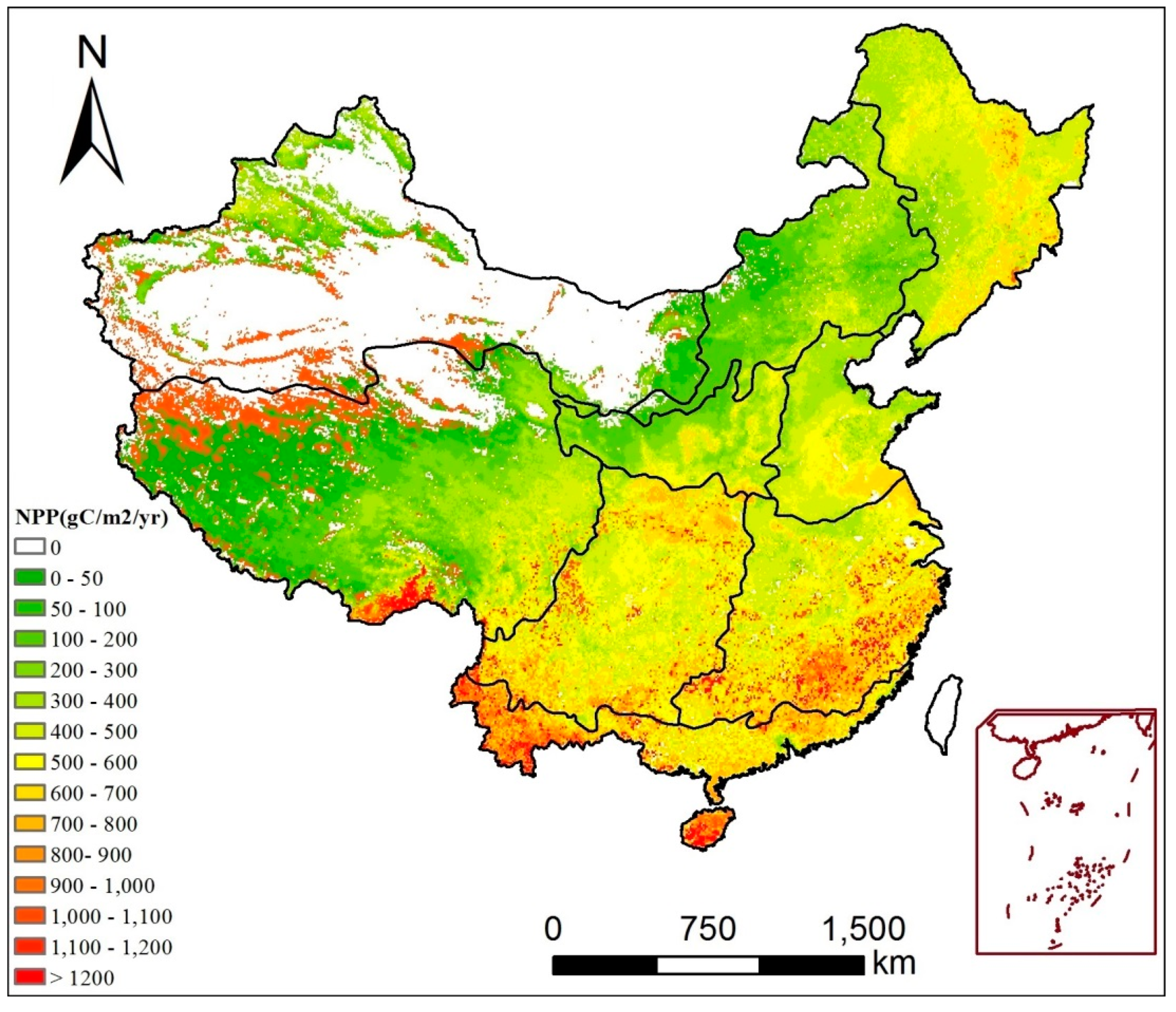
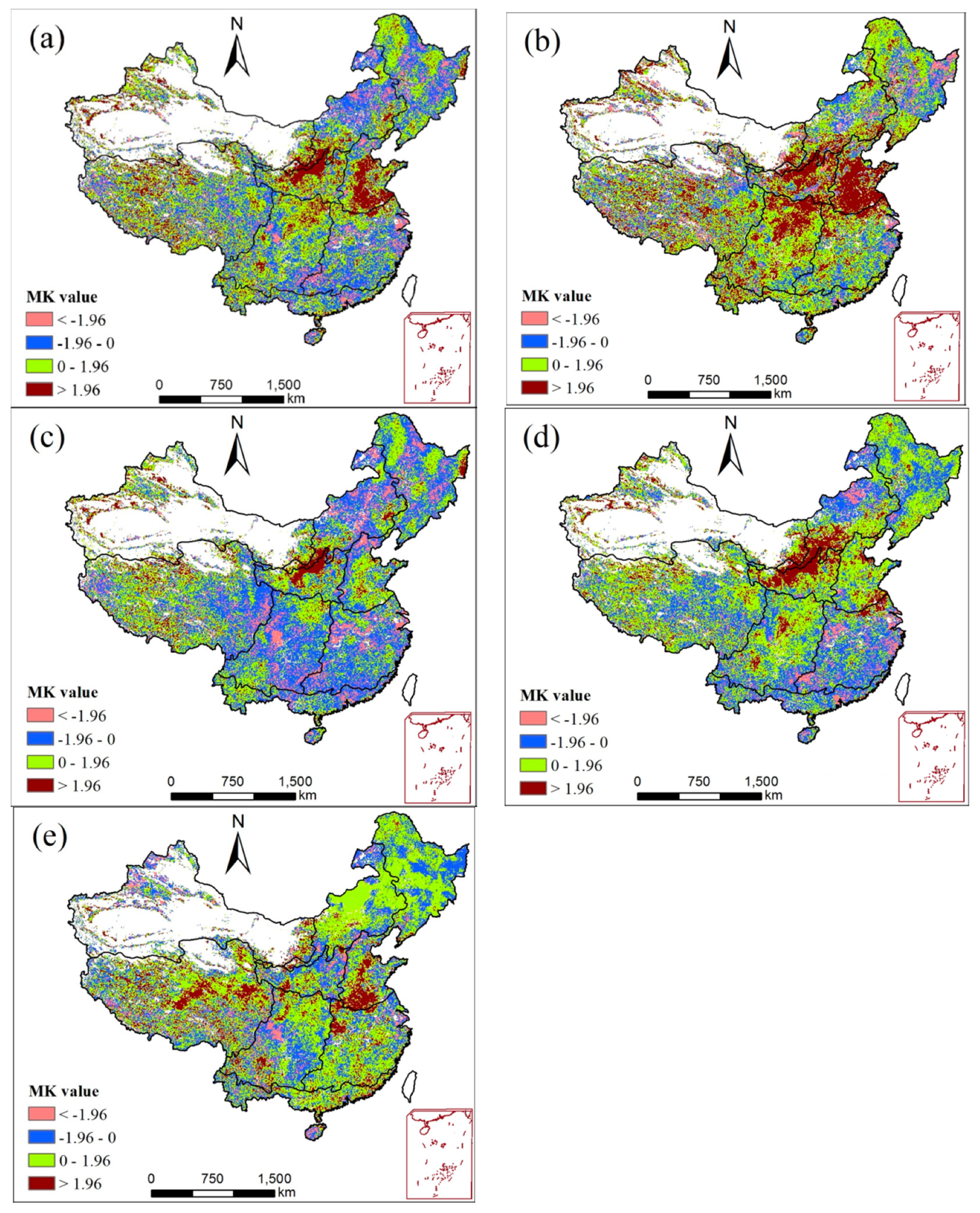
3.2. Spatiotemporal Trends of NPP
3.3. Drought Impact on NPP
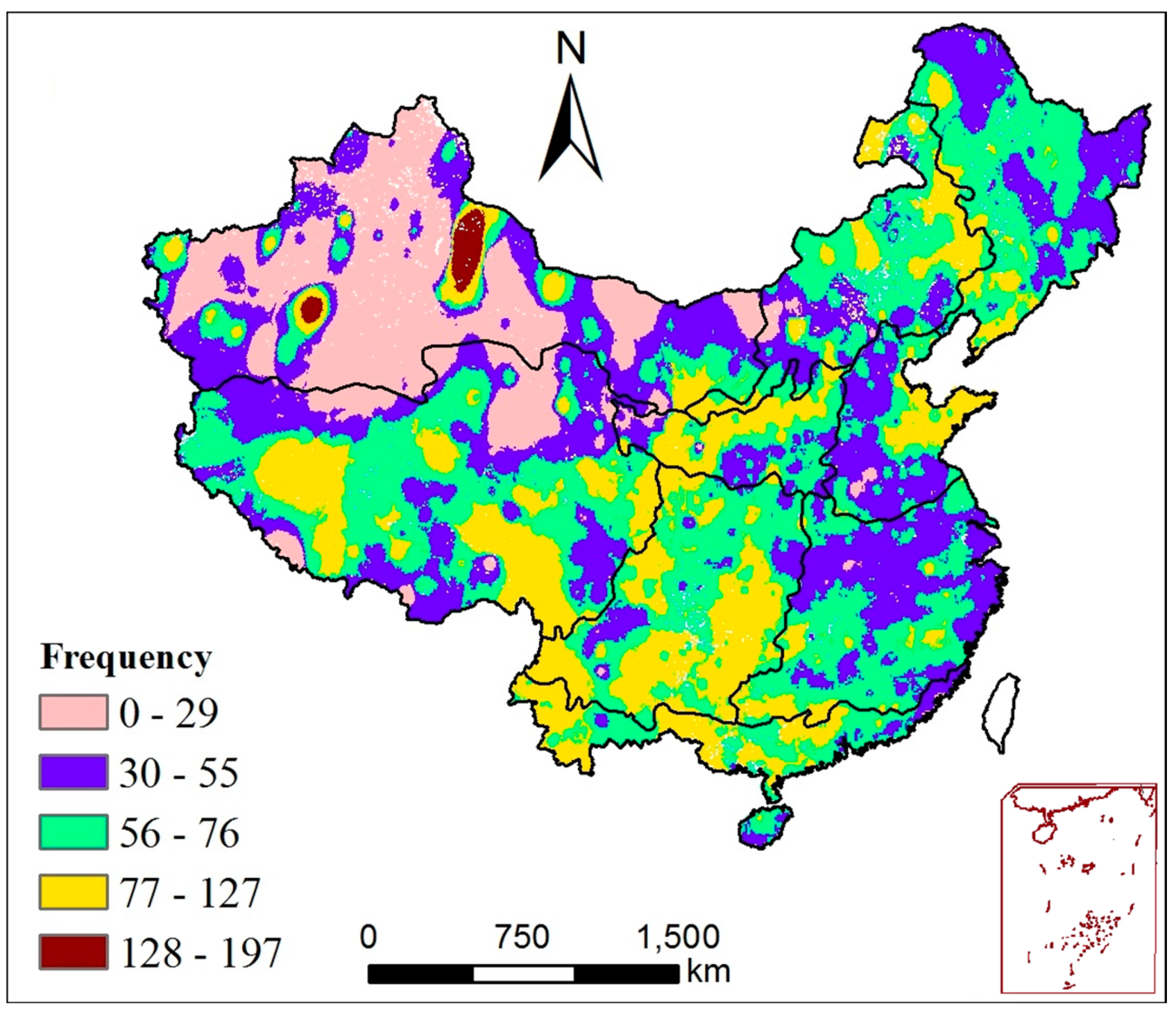
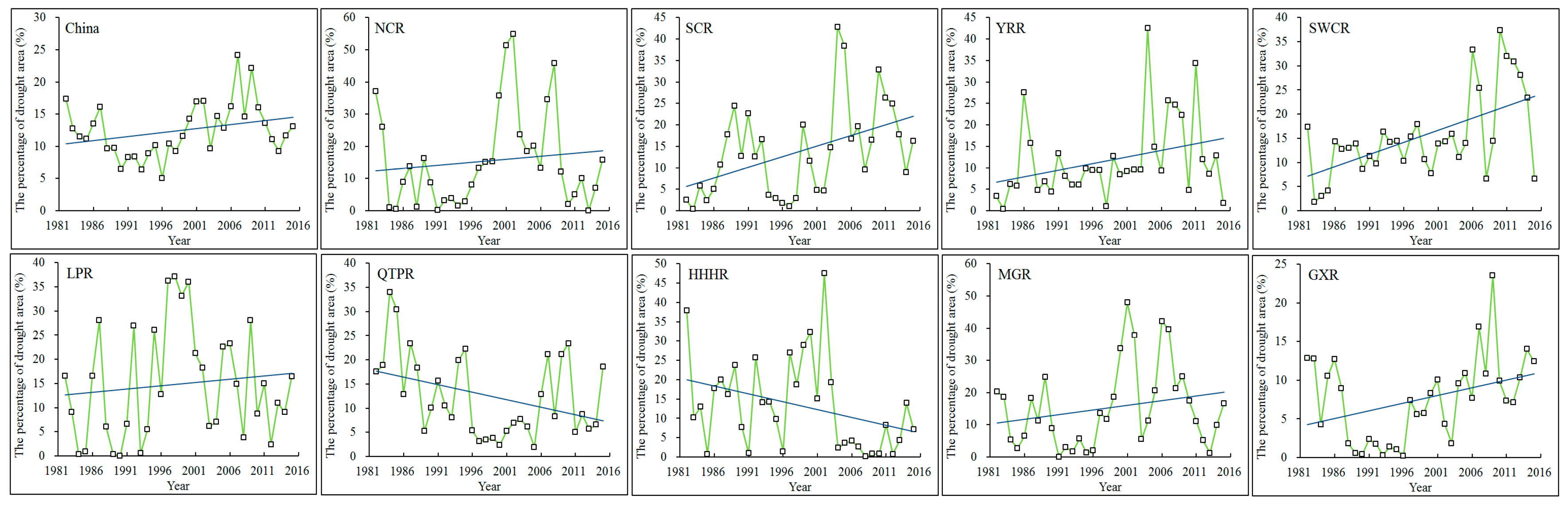
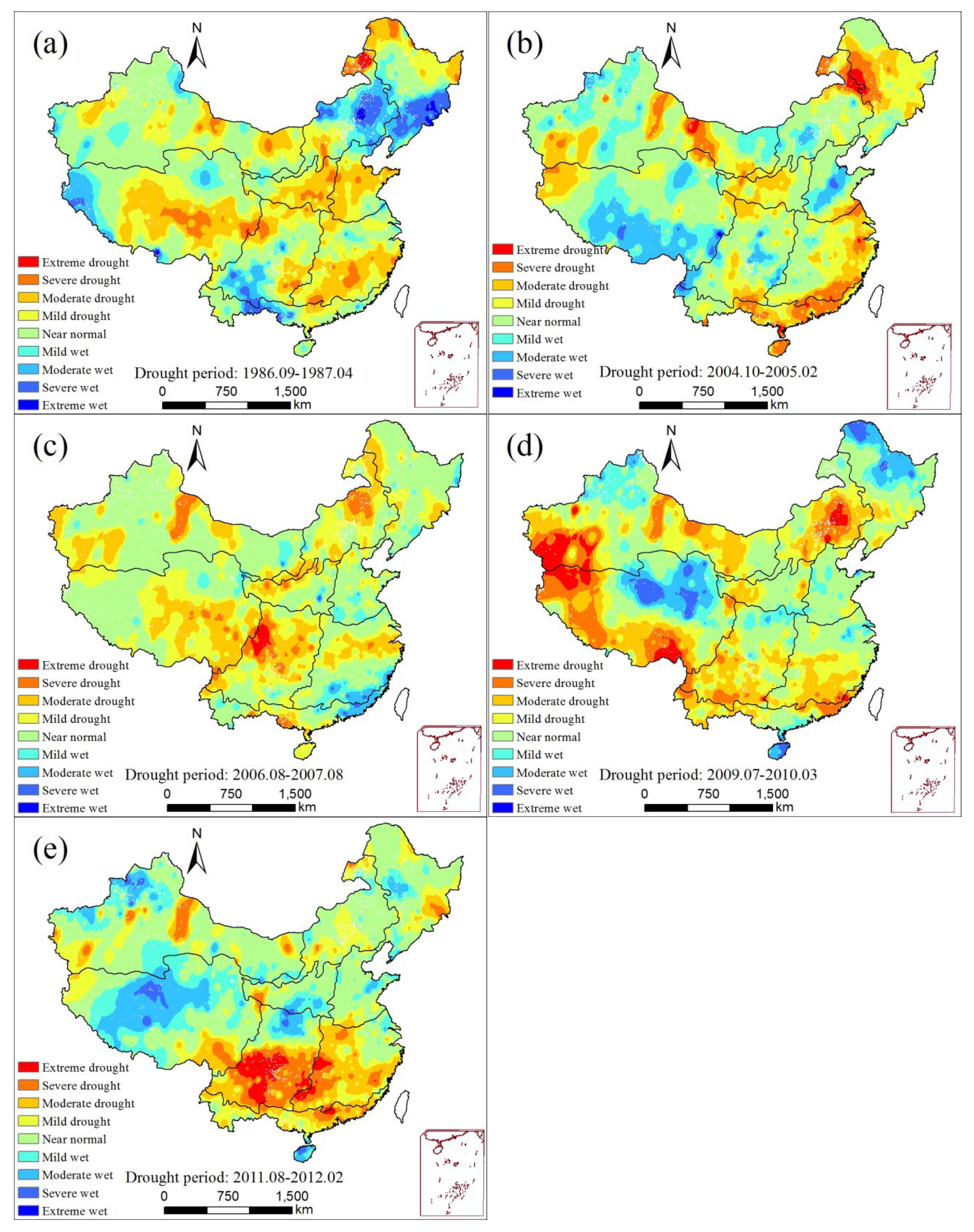
3.3.1. Characteristics of Drought
3.3.2. NPP Variation during Drought Events
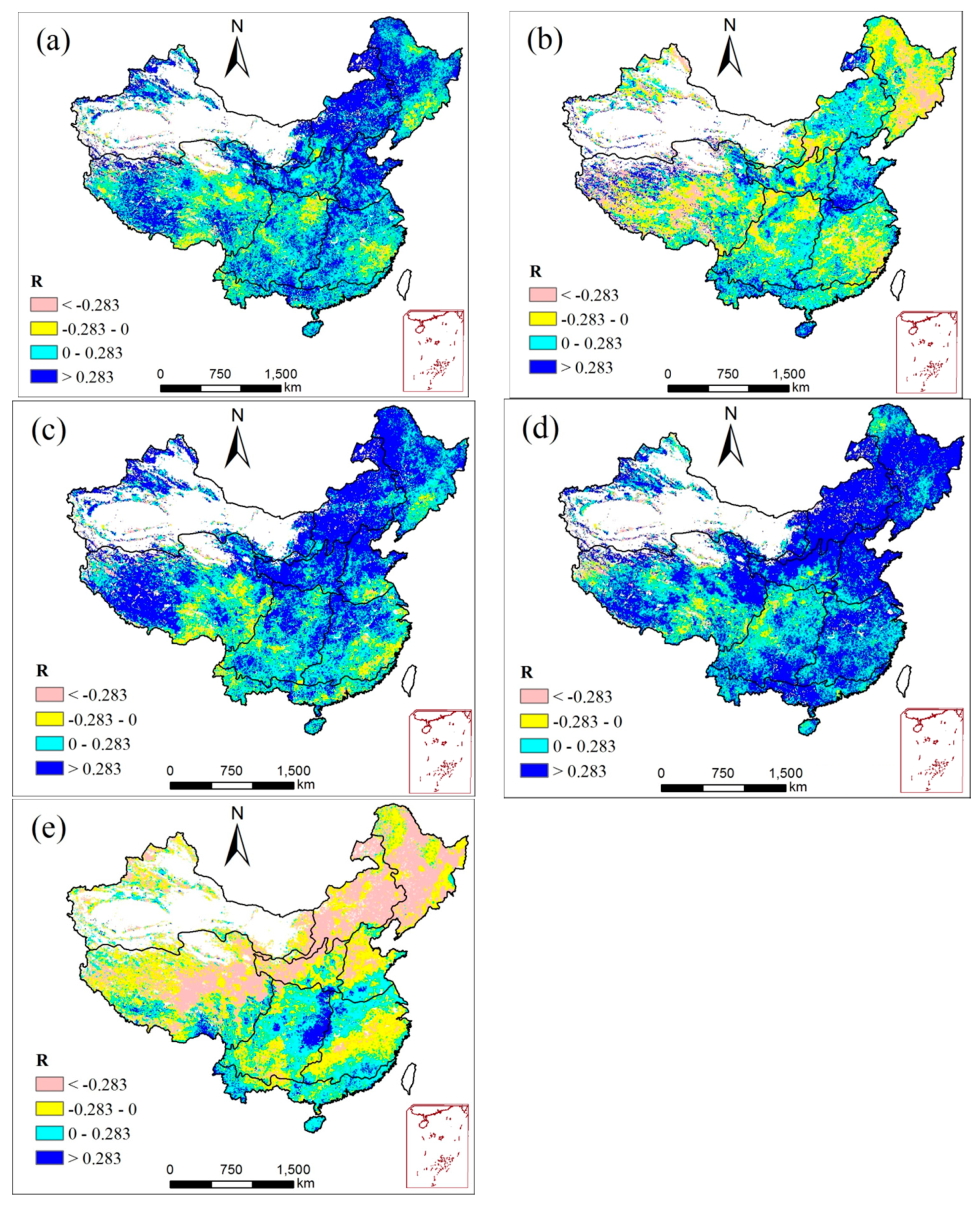
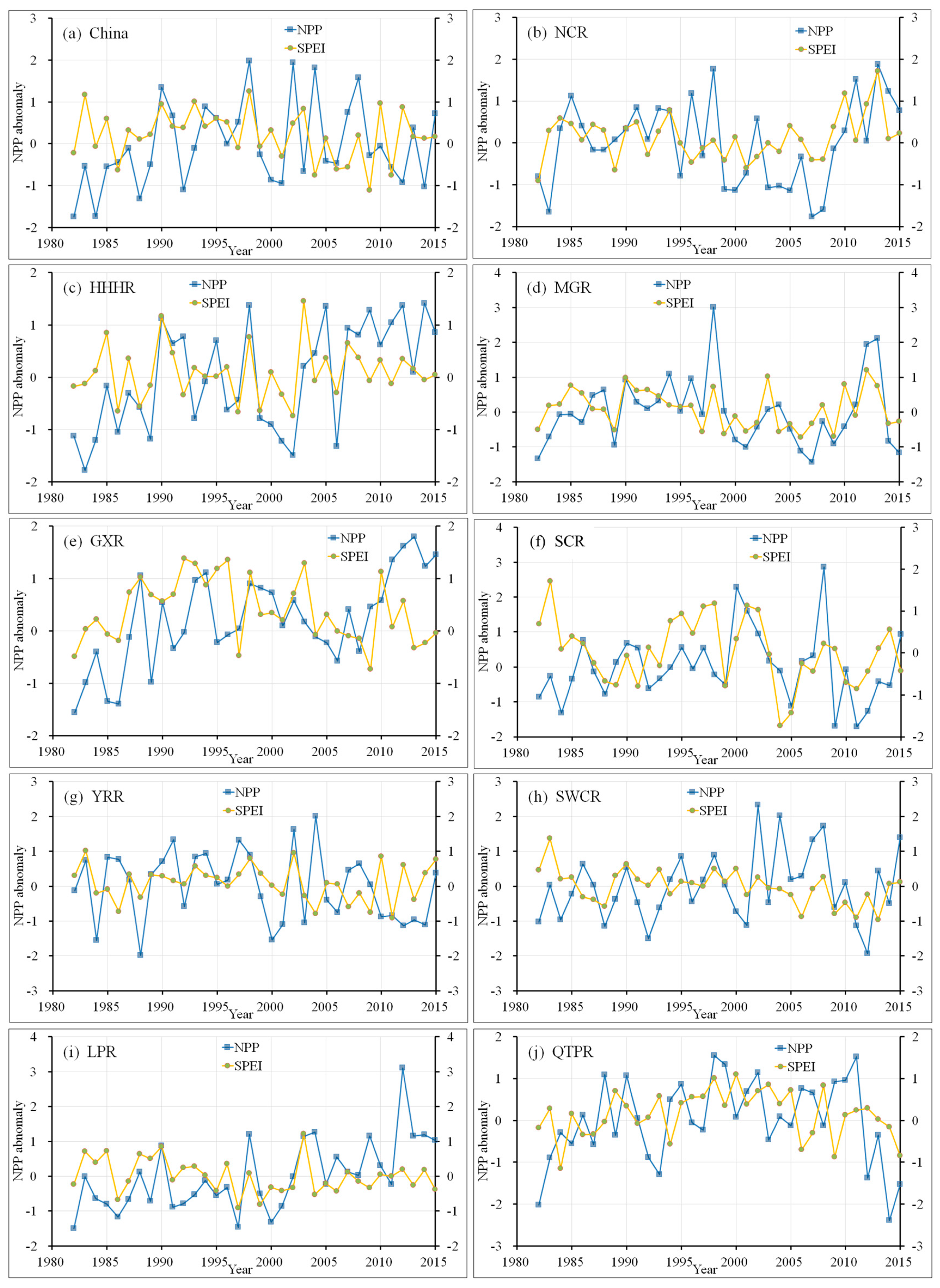
3.3.3. Relationship between NPP and Drought
4. Discussion
4.1. NPP Trends Related to Climate Variation
4.2. NPP Variation Induced by Drought
4.3. Other Potential Factors Affecting NPP Variation
5. Conclusions
- The estimate of NPP across mainland China conducted via the CASA model is satisfactory. For the temporal dimension, the annual NPP of mainland China showed a slightly increased trend from 1982 to 2015; the NPP of spring increased significantly, while the summer NPP showed a higher decreasing trend. For the spatial dimension, the annual NPP across mainland China increased over 54.9% areas and over 13.8% of them significantly increased; 45.1% areas showed a declining trend and 7.2% of them presented a significantly declining trend.
- A positive spatial correlation between annual SPEI and NPP was observed in most areas of mainland China. The temporal relationship between NPP and SPEI showed a significant positive correlation in summer and autumn, while a negative relation was detected in spring and winter. Specific to each region, annual NPP and SPEI showed a significant positive relation in NCR, HHHR, MGR, and GXR. A weak positive correlation was observed for the remaining areas, namely SCR, YRR, SWCR, LPR, and QTPR.
- Based on the SPEI index, typical drought events were identified in the nine regions and in mainland China from 1982 to 2015. There were five typical drought events in mainland China that occurred during the periods of 1986–1987, 2004–2005, 2006–2007, 2009–2010, and 2011–2012. During these drought events, more than 23% of the area of mainland China experienced drought ravage, in which NPP decreased to different extents. The NPP in most sub-regions decreased by approximately 30% during these events, while the NPP in QTPR generally decreased by about 10%. Generally, the NPP showed a reducing trend during the drought events.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Z.; Lai, C.; Wu, X.; Zeng, Z.; Chen, X. Response of net primary production to land use and land cover change in mainland China since the late 1980s. Sci. Total Environ. 2018, 639, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Kurz, W.A.; Stinson, G.; Rampley, G.J.; Dymond, C.C.; Neilson, E.T. Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proc. Natl. Acad. Sci. USA 2008, 105, 1551–1555. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.; Kane, J.M.; Anderegg, L.D. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements—FAO Irrigation and Drainage Paper 56; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998. [Google Scholar]
- Pei, F.; Li, X.; Liu, X.; Lao, C. Assessing the impacts of droughts on net primary productivity in China. J. Environ. Manag. 2013, 114, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Potter, C.S.; Randerson, J.T.; Field, C.B.; Matson, P.A.; Vitousek, P.M.; Mooney, H.A. Terrestrial ecosystem production: A process model based on global satellite and surface data. Glob. Biogeochem. Cycl. 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Roxburgh, S.H.; Berry, S.L.; Buckley, T.N.; Barnes, B.; Roderick, M.L. What is NPP? Inconsistent accounting of respiratory fluxes in the definition of net primary production. Funct. Ecol. 2005, 19, 378–382. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Lai, R.; Zhao, Y.; Jiang, W.; Chen, Q. Estimating net primary production of natural grassland and its spatio-temporal distribution in China. Sci. Total Environ. 2016, 553, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Wu, Z.; Cheng, J.; Yu, Q.; Rao, X. Impacts of urbanization on net primary productivity in the Pearl River Delta, China. Int. J. Plant Prod. 2015, 9, 581–598. [Google Scholar]
- Li, X.; Chen, G.; Liu, X.; Liang, X.; Wang, S.; Chen, Y.; Xu, X. A new global land-use and land-cover change product at a 1-km resolution for 2010 to 2100 based on human–environment interactions. J. Ann. Am. Assoc. Geogr. 2017, 107, 1040–1059. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, L.; Guo, H.; Leinenkugel, P.; Zhou, Y.; Li, L.; Shen, Q. Drought impact on vegetation productivity in the lower Mekong basin. Int. J. Remote Sens. 2014, 35, 2835–2856. [Google Scholar] [CrossRef]
- Zhang, L.; Xiao, J.; Li, J.; Wang, K.; Lei, L.; Guo, H. The 2010 spring drought reduced primary productivity in southwestern China. Environ. Res. Lett. 2012, 7, 045706. [Google Scholar] [CrossRef]
- Xiao, J.; Zhuang, Q. Drought effects on large fire activity in Canadian and Alaskan forests. Environ. Res. Lett. 2007, 2, 044003. [Google Scholar] [CrossRef] [Green Version]
- Van der Molen, M.K.; Dolman, A.J.; Ciais, P. Drought and ecosystem carbon cycling. Agric. For. Meteorol. 2011, 151, 765–773. [Google Scholar] [CrossRef]
- Law, B.E.; Falge, E.; Gu, L. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agric. For. Meteorol. 2002, 113, 97–120. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, Y.; Ju, W.; Wang, S.; Wu, X.; He, M.; Zhu, G. Impacts of droughts on carbon sequestration by China’s terrestrial ecosystems from 2000 to 2011. Biogeosciences 2014, 11, 2583–2599. [Google Scholar] [CrossRef]
- Chen, T.; Werf, G.R.; Jeu, R.A.M.; Wang, G. A global analysis of the impact of drought on net primary productivity. Hydrol. Earth Syst. Sci. 2013, 17, 3885–3894. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Zhang, B.; Wu, C.; Huete, A.R.; Gonsamo, A. Country-level net primary production distribution and response to drought and land cover change. Sci. Total Environ. 2017, 574, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Reichstein, M.; Ciais, P.; Papale, D. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: A joint flux tower, remote sensing and modelling analysis. Glob. Chang. Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Phillips, O.L.; Aragao, L.E.O.C.; Lewis, S.L. Drought sensitivity of the Amazon rainforest. Science 2009, 323, 1344–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.; Pendall, E.; Ewers, B.E.; Cleary, M.; Naithani, K. Spring drought regulates summer net ecosystem CO2 exchange in a sagebrush-steppe ecosystem. Agric. For. Meteorol. 2008, 148, 381–391. [Google Scholar] [CrossRef]
- Xiao, J.F.; Zhuang, Q.L.; Law, B.E. A continuous measure of gross primary production for the conterminous United States derived from MODIS and AmeriFlux data. Remote Sens. Environ. 2010, 114, 576–591. [Google Scholar] [CrossRef]
- Xiao, J.F.; Zhuang, Q.L.; Law, B.E. Assessing net ecosystem carbon exchange of U.S. terrestrial ecosystems by integrating eddy covariance flux measurements and satellite observations. Agric. For. Meteorol. 2011, 151, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Potter, C.; Klooster, S.; Hiatt, C.; Genovese, V.; Castillarubio, J.C. Changes in the carbon cycle of Amazon ecosystems during the 2010 drought. Environ. Res. Lett. 2011, 6, 034024. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Ciais, P.; Huang, Y. The impacts of climate change on water resources and agriculture in China. Nature 2010, 467, 43. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Yang, Y.; Fan, D.; Guan, H.; Zhang, T.; Long, D.; Zhou, Y.; Bai, D. Analysis of spatial and temporal patterns of net primary production and their climate controls in China from 1982 to 2010. Agric. For. Meteorol. 2015, 204, 22–36. [Google Scholar] [CrossRef]
- Yu, G.R.; Zhu, X.J.; Fu, Y.L. Spatial patterns and climate drivers of carbon fluxes in terrestrial ecosystems of China. Glob. Chang. Biol. 2013, 19, 798–810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, T. Drought over East Asia: A review. J. Clim. 2015, 28, 3375–3399. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.; Pan, Y.; Li, W. Spatial and temporal variability of drought in the arid region of China and its relationships to teleconnection indices. J. Hydrol. 2015, 523, 283–296. [Google Scholar] [CrossRef]
- Xu, K.; Yang, D.; Yang, H.; Li, Z.; Qin, Y.; Shen, Y. Spatio-temporal variation of drought in China during 1961–2012: A climatic perspective. J. Hydrol. 2015, 526, 253–264. [Google Scholar] [CrossRef]
- Sun, B.; Zhao, H.; Wang, X. Effects of drought on net primary productivity: Roles of temperature, drought intensity, and duration. Chin. Geogr. Sci. 2016, 26, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, J.; Lai, C.; Zeng, Z.; Zhong, R.; Chen, X.; Zhou, X. Does drought in china show a significant decreasing trend from 1961 to 2009? Sci. Total Environ. 2017, 579, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; He, B.; Quan, X.; Bai, X. Use of the standardized precipitation evapotranspiration index (SPEI) to characterize the drying trend in Southwest China from 1982–2012. Remote Sens. 2015, 7, 10917–10937. [Google Scholar] [CrossRef]
- Pei, F.; Li, X.; Liu, X.; Wang, S.; He, Z. Assessing the differences in net primary productivity between pre- and post-urban land development in China. Agric. For. Meteorol. 2013, 171–172, 174–186. [Google Scholar] [CrossRef]
- He, C.; Liu, Z.; Xu, M.; Ma, Q.; Dou, Y. Urban expansion brought stress to food security in China: Evidence from decreased cropland net primary productivity. Sci. Total Environ. 2017, 576, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Pan, Y.; Zhang, J. Estimation of net primary productivity of Chinese terrestrial vegetation based on remote sensing. J. Plant Ecol. 2007, 31, 413–424. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multi-scalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Chen, H.; Sun, J. Changes in drought characteristics over china using the standardized precipitation evapotranspiration index. J. Clim. 2015, 28, 540–5447. [Google Scholar] [CrossRef]
- Mann, H.B. Nonparametric tests against trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Hafner Publishing Co.: Oxford, UK, 1975. [Google Scholar]
- Wang, Z.; Lai, C.; Chen, X.; Yang, B.; Zhao, S.; Bai, X. Flood hazard risk assessment model based on random forest. J. Hydrol. 2015, 527, 1130–1141. [Google Scholar] [CrossRef]
- Yue, S.; Wang, C.Y. Applicability of prewhitening to eliminate the influence of serial correlation on the Mann-Kendall test. Water Resour. Res. 2002, 38, 4-1–4-7. [Google Scholar] [CrossRef]
- Luo, T.X. Patterns of Net Primary Productivity for Chinese Major Forest Types and Its Mathematical Models. Ph.D. Thesis, Chinese Academy of Sciences, Beijing, China, 1996. [Google Scholar]
- Wu, X.; Guo, S.; Yin, J.; Yang, G.; Zhong, Y.; Liu, D. On the event-based extreme precipitation across China: Time distribution patterns, trends, and return levels. J. Hydrol. 2018, 562, 305–317. [Google Scholar] [CrossRef]
- Xu, X.G.; Lin, H.P.; Hou, L.S.; Yao, X. An assessment for sustainable developing capability of integrated agricultural regionalization in China. Chin. Geogr. Sci. 2002, 12, 1–8. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, P.; Lai, C.; Chen, X.; Wu, X.; Zeng, Z. Spatiotemporal variability of reference evapotranspiration and contributing climatic factors in China during 1961–2013. J. Hydrol. 2017, 544, 97–108. [Google Scholar] [CrossRef]
- Piao, S.L.; Fang, J.Y.; Guo, Q.H. Application of CASA model to the estimation of Chinese terrestrial net primary productivity. Acta Phytoecol. Sin. 2001, 25, 603–608. [Google Scholar]
- Mohammat, A.; Wang, X.; Xu, X.; Peng, L.; Yang, Y.; Zhang, X. Drought and spring cooling induced recent decrease in vegetation growth in Inner Asia. Agric. For. Meteorol. 2013, 178, 21–30. [Google Scholar] [CrossRef]
- Wang, H.; Chen, A.; Wang, Q.; He, B. Drought dynamics and impacts on vegetation in China from 1982 to 2011. Ecol. Eng. 2015, 75, 303–307. [Google Scholar] [CrossRef]
- Mao, J.; Shi, X.; Thornton, P.E.; Wang, X. Causes of spring vegetation growth trends in the northern mid–high latitudes from 1982 to 2004. Environ. Res. Lett. 2012, 7, 014010. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ju, W.; He, H.; Wang, S.; Sun, R.; Zhang, Y. Changes of net primary productivity in China during recent 11 years detected using an ecological model driven by MODIS data. Front. Earth Sci. 2013, 7, 112–127. [Google Scholar] [CrossRef]
- Wang, Z.; Zhong, R.; Lai, C.; Chen, J. Evaluation of the GPM IMERG satellite-based precipitation products and the hydrological utility. Atmos. Res. 2017, 196, 151–163. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhong, R.; Lai, C.; Zeng, Z.; Lian, Y.; Bai, X. Climate change enhances the severity and variability of drought in the Pearl River Basin in South China in the 21st century. Agric. For. Meteorol. 2018, 249, 149–162. [Google Scholar] [CrossRef]
- Shi, Z.; Thomey, M.L.; Mowll, W.; Litvak, M.; Brunsell, N.A.; Collins, S.L.; Pockman, W.T.; Smith, M.D.; Knapp, A.K. Differential effects of extreme drought on production and respiration: Synthesis and modeling analysis. Biogeosci. Discuss. 2013, 10, 16043–16074. [Google Scholar] [CrossRef]
- Li, Z.; Liu, G.; Fu, B.; Zhang, J. The potential influence of seasonal climate variables on the net primary production of forests in Eastern China. Environ. Manag. 2011, 48, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Kumar, S. Responses to winter dormancy, temperature, and plant hormones share gene networks. Funct. Integr. Genom. 2011, 11, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Saleska, S.R.; Didan, K.; Huete, A.R.; da Rocha, H.R. Amazon forests green-up during 2005 drought. Science 2007, 318, 612. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; He, B.; Chen, A.; Wang, H.; Liu, J.; Lu, A.; Chen, Z. Drought dominates the interannual variability in global terrestrial net primary production by controlling semi-arid ecosystems. Sci. Rep. 2016, 6, 24639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Z.; An, S.; Cheng, M.; Wang, W. Plant functional traits and soil microbial biomass in different vegetation zones on the Loess Plateau. J. Plant Interact. 2014, 9, 889–900. [Google Scholar] [CrossRef] [Green Version]
- Willson, C.J.; Manos, P.S.; Jackson, R.B. Hydraulic traits are influenced by phylogenetic history in the drought-resistant, invasive genus Juniperus (Cupressaceae). Am. J. Bot. 2008, 95, 299–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Moran, M.S.; Zhao, X.; Liu, S.; Zhou, T.; Ponce-Campos, G.E.; Liu, F. Impact of prolonged drought on rainfall use efficiency using MODIS data across china in the early 21st century. Remote Sens. Environ. 2014, 150, 188–197. [Google Scholar] [CrossRef]
- Xiong, H.; Wang, S.; Rong, L.; Cheng, A.; Li, Y. Effects of extreme drought on plant species in karst area of Guizhou Province, southwest China. Chin. J. Appl. Ecol. 2011, 22, 1127–1134. [Google Scholar]
- Zhang, C.; Ju, W.; Chen, J.; Wang, X.; Yang, L.; Zheng, G. Disturbance-induced reduction of biomass carbon sinks of China’s forests in recent years. Environ. Res. Lett. 2015, 10, 114021. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Liu, X.; Wang, F.; Li, X.; Ou, J. Assessing the impacts of urban sprawl on net primary productivity using fusion of Landsat and MODIS data. Sci. Total Environ. 2018, 613, 1417–1429. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.; Kyle, G.P.; Dooley, J.J. The impact of electric passenger transport technology under an economy-wide climate policy in the United States: Carbon dioxide emissions, coal use, and carbon dioxide capture and storage. Int. J. Greenh. Gas Control. 2010, 4, 301–308. [Google Scholar] [CrossRef]
- Shochat, E.; Warren, P.S.; Faeth, S.H.; McIntyre, N.E. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 2006, 21, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Clerici, N.; Paracchini, M.L.; Maes, J. Land-cover change dynamics and insights into ecosystem services in European stream riparian zones. Ecohydrol. Hydrobiol. 2014, 14, 107–120. [Google Scholar] [CrossRef]
- Pei, F.; Li, X.; Liu, X.; Lao, C. Exploring the response of net primary productivity variations to urban expansion and climate change: A scenario analysis for Guangdong Province in China. J. Environ. Manag. 2015, 150, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Qiao, Z. Assessing the impact of the urbanization process on net primary productivity in China in 1989–2000. Environ. Pollut. 2014, 184, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Wang, W.; Zeng, W.; Zeng, H. Nitrogen deposition enhances carbon sequestration by plantations in northern China. PLoS ONE 2014, 9, e87975. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Chen, J.M.; Liu, J.; Bélair, S.; Luo, X. Assessment of SMAP soil moisture for global simulation of gross primary production. J. Geophys. Res. Biogeosci. 2017, 122, 1549–1563. [Google Scholar] [CrossRef]
- Woodward, F.I.; Kelly, C.K. Responses of global plant diversity capacity to changes in carbon dioxide concentration and climate. Ecol. Lett. 2008, 11, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, J.; Lai, C.; Wang, R.Y.; Chen, X.; Lian, Y. Drying tendency dominating the global grain production area. Glob. Food Sec. 2018, 16, 138–149. [Google Scholar] [CrossRef]
- Jeong, S.J.; HO, C.H.; GIM, H.J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | SPEI Value | Category | SPEI Value |
|---|---|---|---|
| Extreme drought | Less than −2 | Mild wet | 0.50 to 0.99 |
| Severe drought | −1.99 to −1.5 | Moderate wet | 1.0 to 1.49 |
| Moderate drought | −1.49 to −1.0 | Severe wet | 1.50 to 1.99 |
| Mild drought | −0.99 to −0.50 | Extreme wet | More than 2 |
| Near normal | −0.49 to 0.49 |
| Region | Year | Spring | Summer | Autumn | Winter | |
|---|---|---|---|---|---|---|
| MK | Slope (Pg C Year−1) | MK | MK | MK | MK | |
| NCR | 0.237 | 0.005 | 0.474 | −0.801 | 0.623 | 0.682 |
| SCR | 0.237 | 0.004 | 1.749 | −0.919 | −1.542 | 0.937 |
| YRR | −1.364 | −0.027 | 0.534 | −1.957 | −1.838 | 0.937 |
| SWCR | 1.186 | 0.020 | 2.757 | −0.919 | 1.127 | 0.511 |
| LPR | 3.795 | 0.058 | 2.935 | 2.609 | 3.172 | 0.312 |
| QTPR | 0.771 | 0.018 | 1.660 | −0.385 | 0.326 | 0.256 |
| HHHR | 3.024 | 0.058 | 4.566 | 0.504 | 2.135 | 1.988 |
| MGR | −0.623 | −0.010 | 2.520 | −1.275 | 0.949 | 0.227 |
| GXR | 1.838 | 0.058 | 1.305 | 1.275 | 2.105 | −0.966 |
| Mainland China | 0.771 | 0.025 | 3.083 | −1.156 | 0.237 | 1.25 |
| Time Scale | Non-Significant Increase (%) | Significant Increase (%, p < 0.05) | Non-Significant Decrease (%) | Significant Decrease (%, p < 0.05) |
|---|---|---|---|---|
| Year | 54.9 | 13.8 | 45.1 | 7.2 |
| Spring | 67.6 | 23.9 | 32.4 | 6.7 |
| Summer | 43.7 | 8.5 | 56.3 | 10.4 |
| Autumn | 55.9 | 11.6 | 44.1 | 5.4 |
| Winter | 60.6 | 9.8 | 39.4 | 5.9 |
| Region | Persistent Period (Month Year) | Duration (Month) | Maximum Affected Area (105 km2) | Percentage of Drought Areas (%) | Severity | Intensity |
|---|---|---|---|---|---|---|
| Mainland China | 09.2006–08.2007 | 12 | 28.9 | 33.4 | 15.56 | −1.9 |
| 07.2009–03.2010 | 9 | 32.7 | 37.8 | 11.55 | −1.8 | |
| 08.2011–02.2012 | 7 | 20.8 | 24.0 | 7.72 | −1.5 | |
| 09.1986–04.1987 | 8 | 22.4 | 25.8 | 7.38 | −1.2 | |
| 10.2004–02.2005 | 5 | 19.7 | 22.7 | 4.54 | −1.2 | |
| NCR | 09.2001–07.2002 | 11 | 7.28 | 73.2 | 19.38 | −2.1 |
| 07.2007–06.2008 | 12 | 7.01 | 70.4 | 17.46 | −2 | |
| 07.1982–05.1983 | 11 | 6.42 | 64.5 | 15.34 | −1.7 | |
| 10.2004–03.2005 | 6 | 3.88 | 39.0 | 6.01 | −1.3 | |
| 06.2000–10.2000 | 5 | 4.76 | 47.9 | 4.54 | −1.4 | |
| SCR | 10.2003–09.2004 | 12 | 2.86 | 77.2 | 19.39 | −2.2 |
| 06.2011–03.2012 | 10 | 2.60 | 70.2 | 11.80 | −1.8 | |
| 2009.10–2010.05 | 8 | 2.27 | 61.2 | 8.50 | −1.5 | |
| 08.1989–02.1990 | 7 | 1.76 | 47.6 | 8.11 | −1.5 | |
| 04.1999–08.1999 | 5 | 2.44 | 66.0 | 5.68 | −1.7 | |
| YRR | 02.2004–01.2005 | 12 | 4.96 | 61.8 | 16.26 | −1.8 |
| 05.2011–02.2012 | 10 | 4.28 | 49.8 | -14.18 | −1.8 | |
| 07.2007–05.2008 | 11 | 4.42 | 55.1 | 11.12 | −1.7 | |
| 08.1986–04.1987 | 9 | 3.76 | 46.8 | 9.76 | −1.3 | |
| 08.2009–12.2009 | 5 | 3.68 | 45.8 | 4.64 | −1.3 | |
| SWCR | 07.2006–06.2007 | 12 | 5.36 | 62.9 | 22.71 | −2.3 |
| 07.2011–06.2012 | 12 | 5.67 | 66.5 | 21.62 | −2.3 | |
| 09.2009–09.2010 | 13 | 5.14 | 60.4 | 20.13 | −2.1 | |
| 07.2013–07.2014 | 13 | 4.85 | 56.9 | 17.53 | −1.5 | |
| 11.1992–07.1993 | 9 | 4.07 | 47.7 | 8.73 | −1.5 | |
| LPR | 08.1997–06.1998 | 11 | 3.13 | 87.1 | 22.67 | −2.8 |
| 09.1986–05.1987 | 9 | 2.19 | 60.7 | 10.39 | −1.5 | |
| 05.1999–10.1999 | 6 | 2.53 | 70.3 | 7.52 | −2.3 | |
| 04.2000–09.2000 | 6 | 2.66 | 73.8 | 7.41 | −1.9 | |
| 02.1992–07.1992 | 6 | 2.92 | 81.2 | 6.91 | −1.7 | |
| QTPR | 08.1984–08.1985 | 13 | 1.14 | 58.2 | 19.86 | −2.0 |
| 08.1994–07.1995 | 12 | 1.04 | 53.1 | 17.10 | −1.8 | |
| 08.2006–06.2007 | 11 | 6.79 | 34.5 | 13.50 | −1.6 | |
| 09.1986–06.1987 | 10 | 7.44 | 37.8 | 10.50 | −1.3 | |
| 09.2009–03.2010 | 7 | 8.80 | 44.7 | 7.70 | −1.3 | |
| HHHR | 07.2002–05.2003 | 11 | 2.61 | 65.2 | 13.16 | −1.5 |
| 1982.01–1982.07 | 7 | 2.96 | 74.0 | 10.50 | −1.9 | |
| 1997.08–1998.04 | 9 | 3.30 | 82.6 | 10.16 | −1.8 | |
| 07.1999–01.2000 | 7 | 3.87 | 96.8 | 8.69 | −2.0 | |
| 11.2001–04.2002 | 6 | 2.79 | 69.7 | 7.32 | −1.8 | |
| MGR | 04.2001–05.2002 | 14 | 5.39 | 69.6 | 19.79 | −1.8 |
| 06.2007–05.2008 | 12 | 4.35 | 56.2 | 14.78 | −1.5 | |
| 03.2006–02.2007 | 12 | 4.56 | 59.0 | 13.34 | −1.3 | |
| 08.2009–03.2010 | 8 | 4.29 | 55.4 | 9.72 | −1.6 | |
| 02.2000–07.2000 | 6 | 4.90 | 63.4 | 4.90 | −1.6 |
| Period | Positive Correlation (%) | Significantly Positive Correlation (%) | Negative Correlation (%) | Significantly Negative Correlation (%) |
|---|---|---|---|---|
| Year | 82.0 | 37.7 | 18 | 1.8 |
| Spring | 51.8 | 0.98 | 48.2 | 9.2 |
| Summer | 83.8 | 45.4 | 16.2 | 1.4 |
| Autumn | 88.7 | 53.0 | 11.3 | 0.9 |
| Winter | 37.5 | 7 | 62.5 | 30 |
| Region | Year | Spring | Summer | Autumn | Winter |
|---|---|---|---|---|---|
| NCR | 0.409 | −0.374 | 0.341 | 0.548 | −0.387 |
| SCR | 0.272 | 0.052 | 0.38 | 0.505 | 0.277 |
| YRR | 0.175 | −0.147 | 0.113 | 0.432 | 0.118 |
| SWCR | 0.129 | 0.025 | 0.228 | 0.235 | 0.112 |
| LPR | 0.227 | 0.084 | 0.519 | 0.402 | −0.155 |
| QTPR | 0.137 | −0.198 | −0.132 | 0.211 | −0.096 |
| HHHR | 0.485 | 0.34 | 0.331 | 0.575 | 0.001 |
| MGR | 0.632 | 0.078 | 0.688 | 0.639 | −0.622 |
| GXR | 0.334 | −0.291 | 0.444 | 0.401 | −0.326 |
| Mainland China | 0.134 | −0.216 | 0.378 | 0.322 | −0.119 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, C.; Li, J.; Wang, Z.; Wu, X.; Zeng, Z.; Chen, X.; Lian, Y.; Yu, H.; Wang, P.; Bai, X. Drought-Induced Reduction in Net Primary Productivity across Mainland China from 1982 to 2015. Remote Sens. 2018, 10, 1433. https://0-doi-org.brum.beds.ac.uk/10.3390/rs10091433
Lai C, Li J, Wang Z, Wu X, Zeng Z, Chen X, Lian Y, Yu H, Wang P, Bai X. Drought-Induced Reduction in Net Primary Productivity across Mainland China from 1982 to 2015. Remote Sensing. 2018; 10(9):1433. https://0-doi-org.brum.beds.ac.uk/10.3390/rs10091433
Chicago/Turabian StyleLai, Chengguang, Jun Li, Zhaoli Wang, Xiaoqing Wu, Zhaoyang Zeng, Xiaohong Chen, Yanqing Lian, Haijun Yu, Peng Wang, and Xiaoyan Bai. 2018. "Drought-Induced Reduction in Net Primary Productivity across Mainland China from 1982 to 2015" Remote Sensing 10, no. 9: 1433. https://0-doi-org.brum.beds.ac.uk/10.3390/rs10091433