Phenology and Seasonal Ecosystem Productivity in an Amazonian Floodplain Forest

 , , ,
, , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
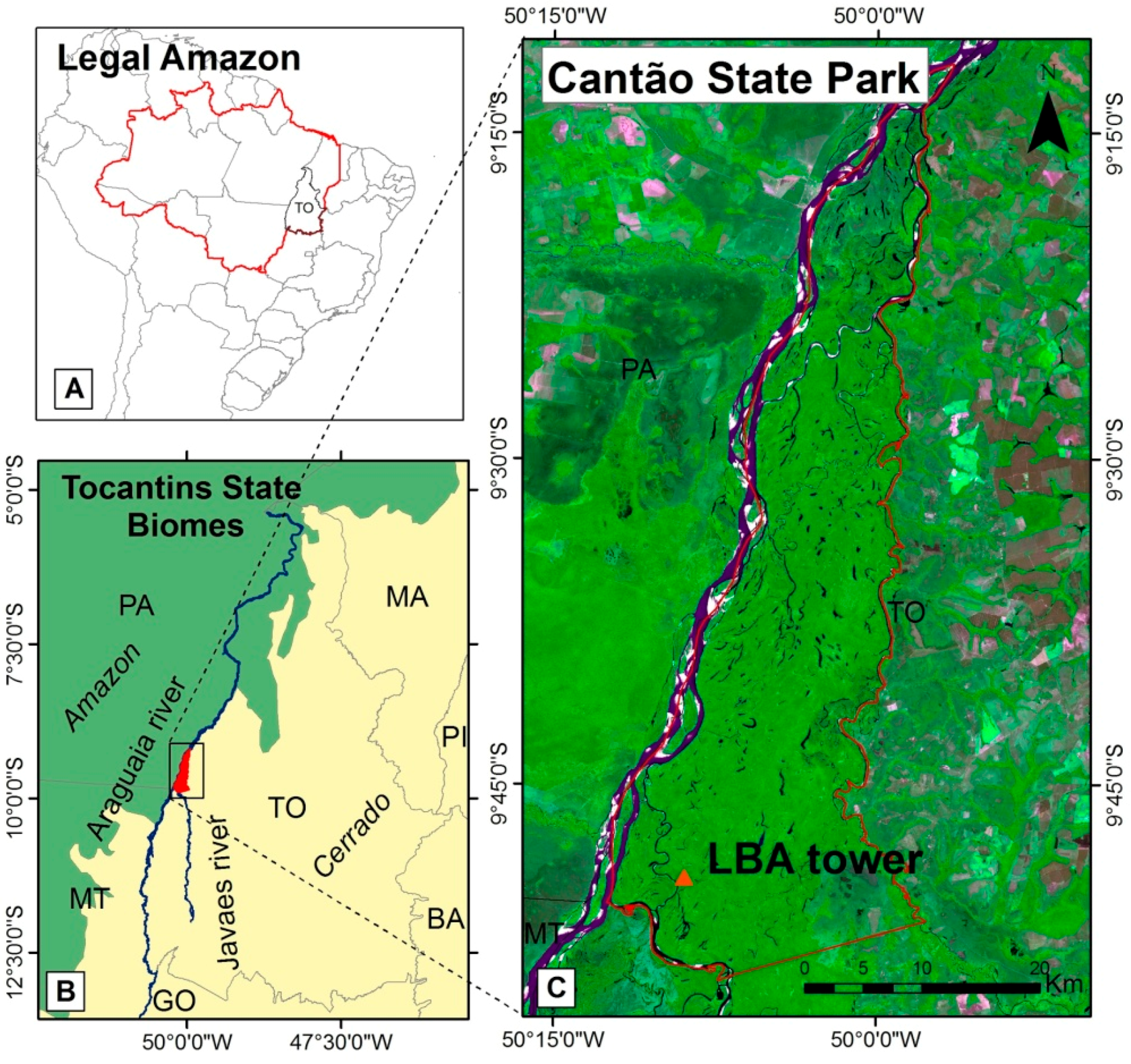
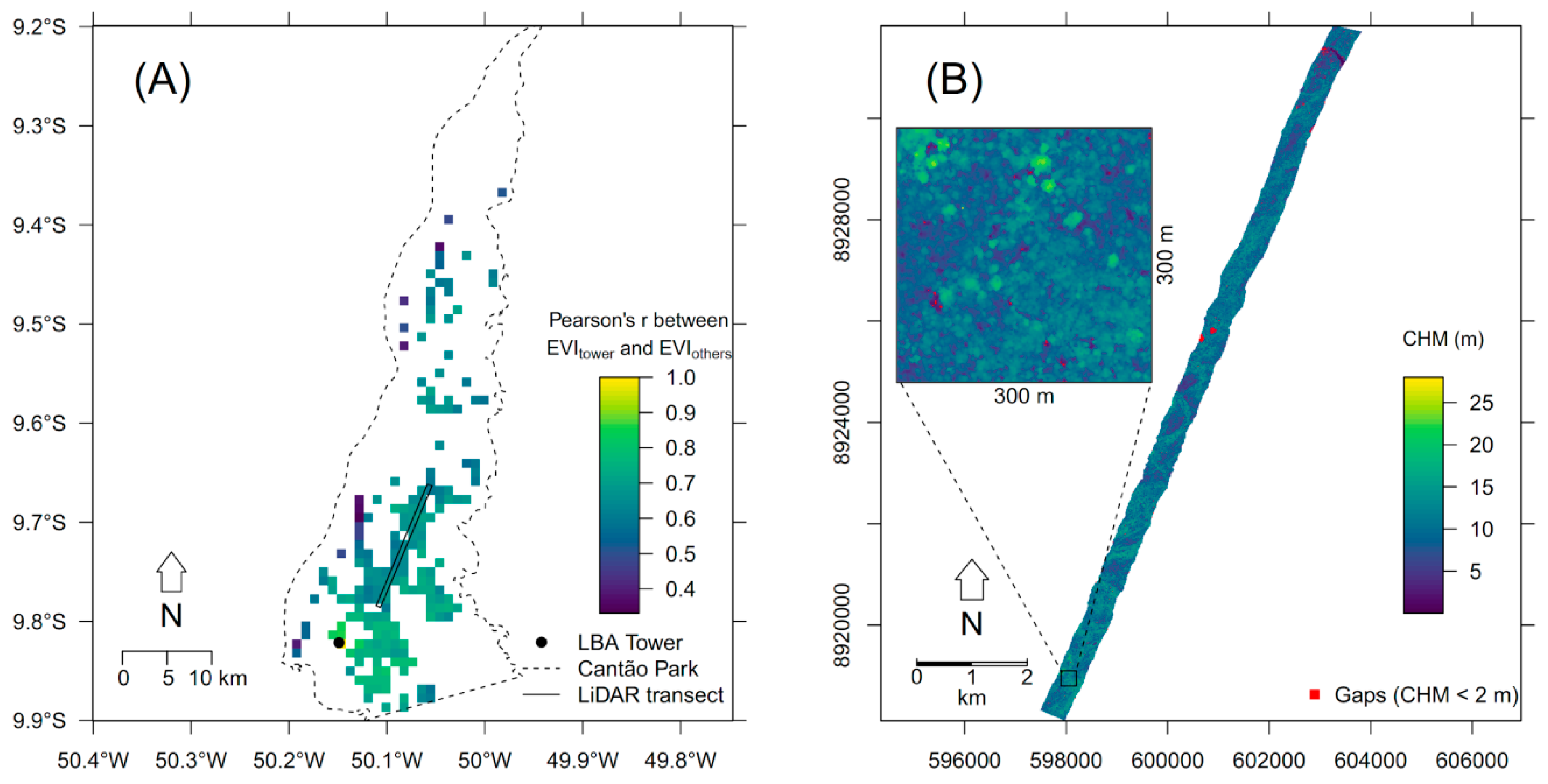
2.1. Study Site
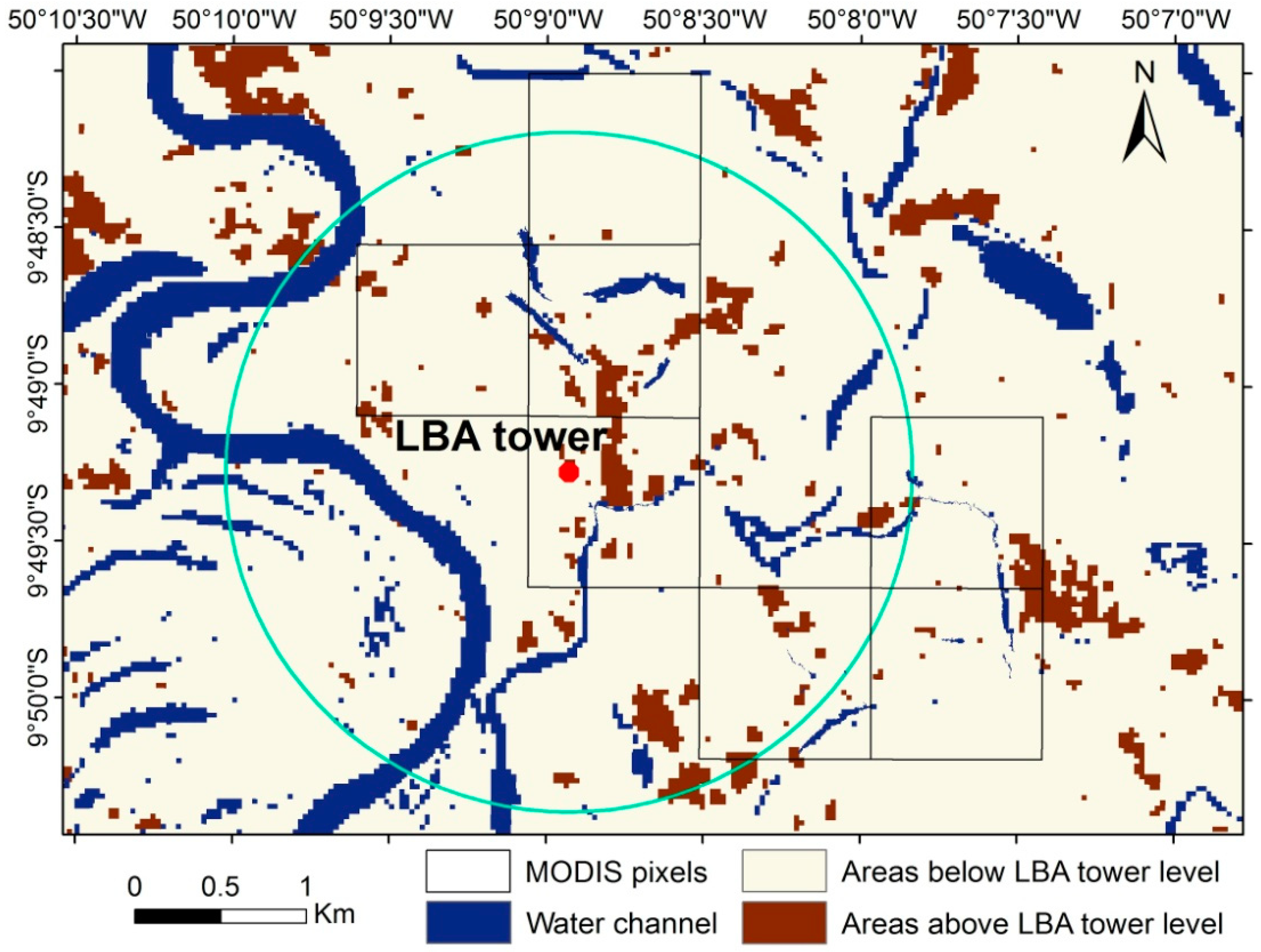
2.2. Flux Tower and Field Data
where λ (J kg−1) = 103 ∗ (2500 − 2.37 ∗ Ta)
Gap Filling of CO2 Estimates
2.3. Litterfall Collection
2.4. Remote-Sensing Data and Products
If CWDm > 0 then CWDm = 0
2.5. Statistical Analysis
3. Results
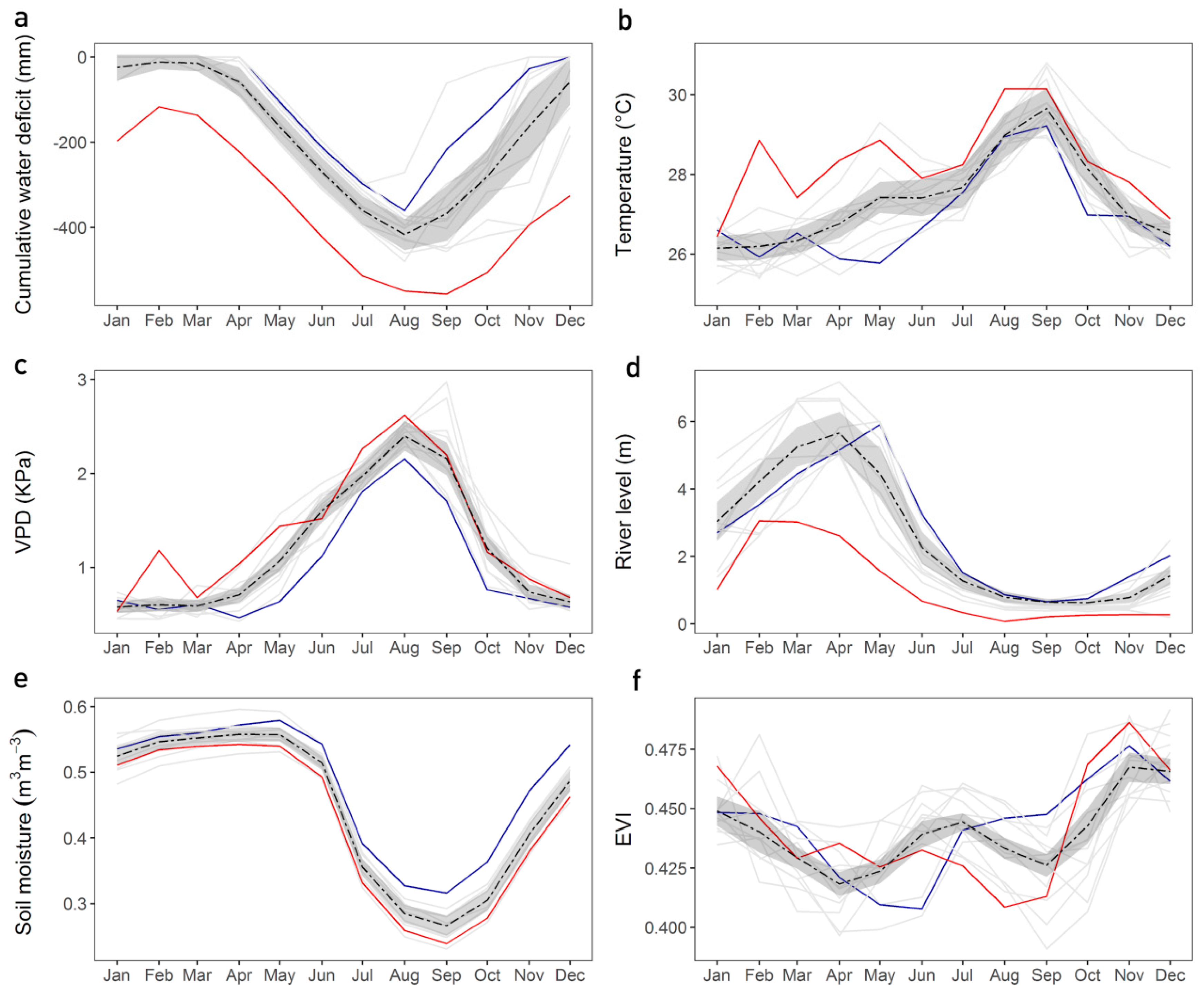
3.1. Seasonal Meteorological, Gross Primary Productivity (GPP) and Enhanced Vegetation Index (EVI) Patterns
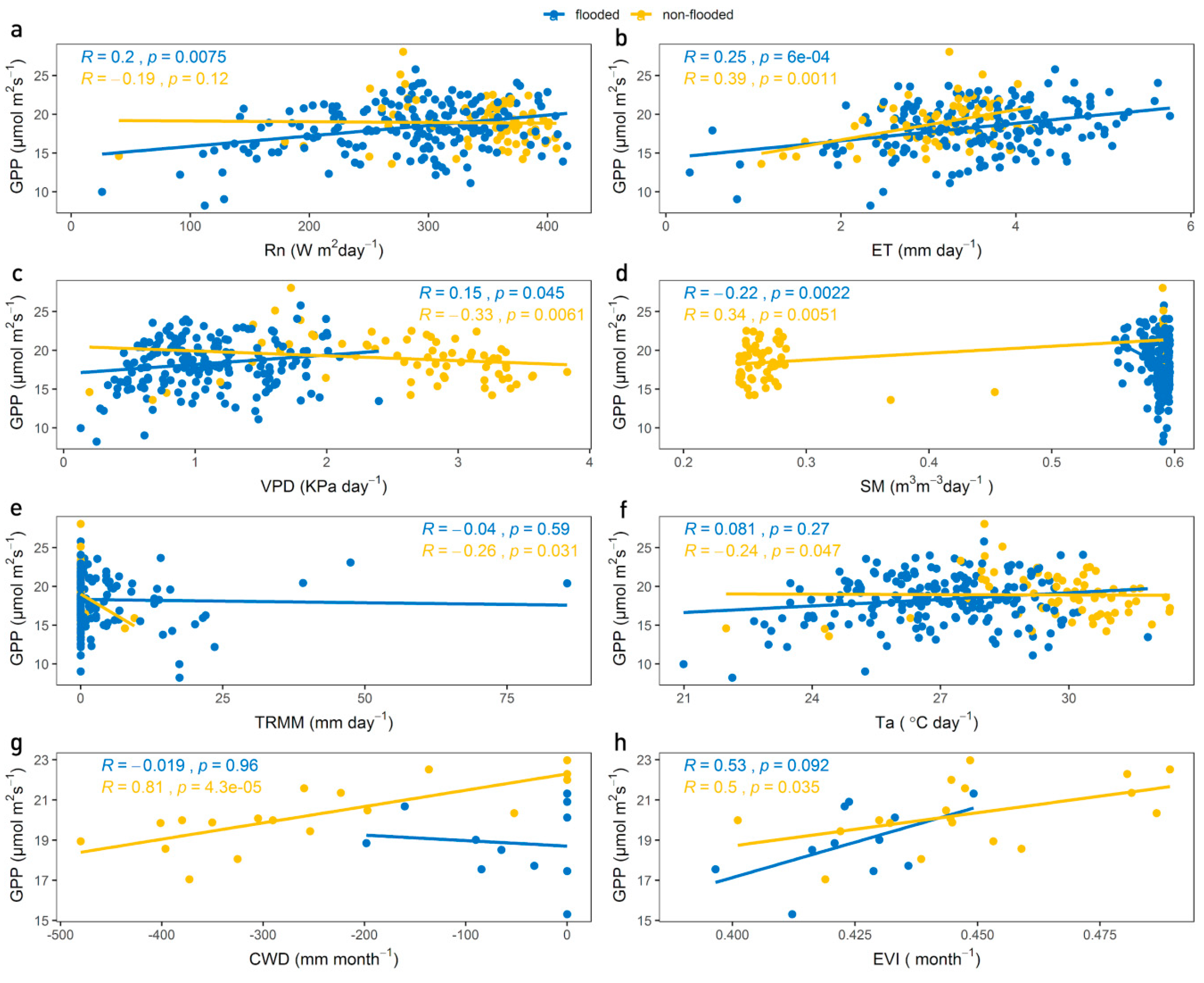
3.2. Correlation Between GPP, Climatic Variables and EVI
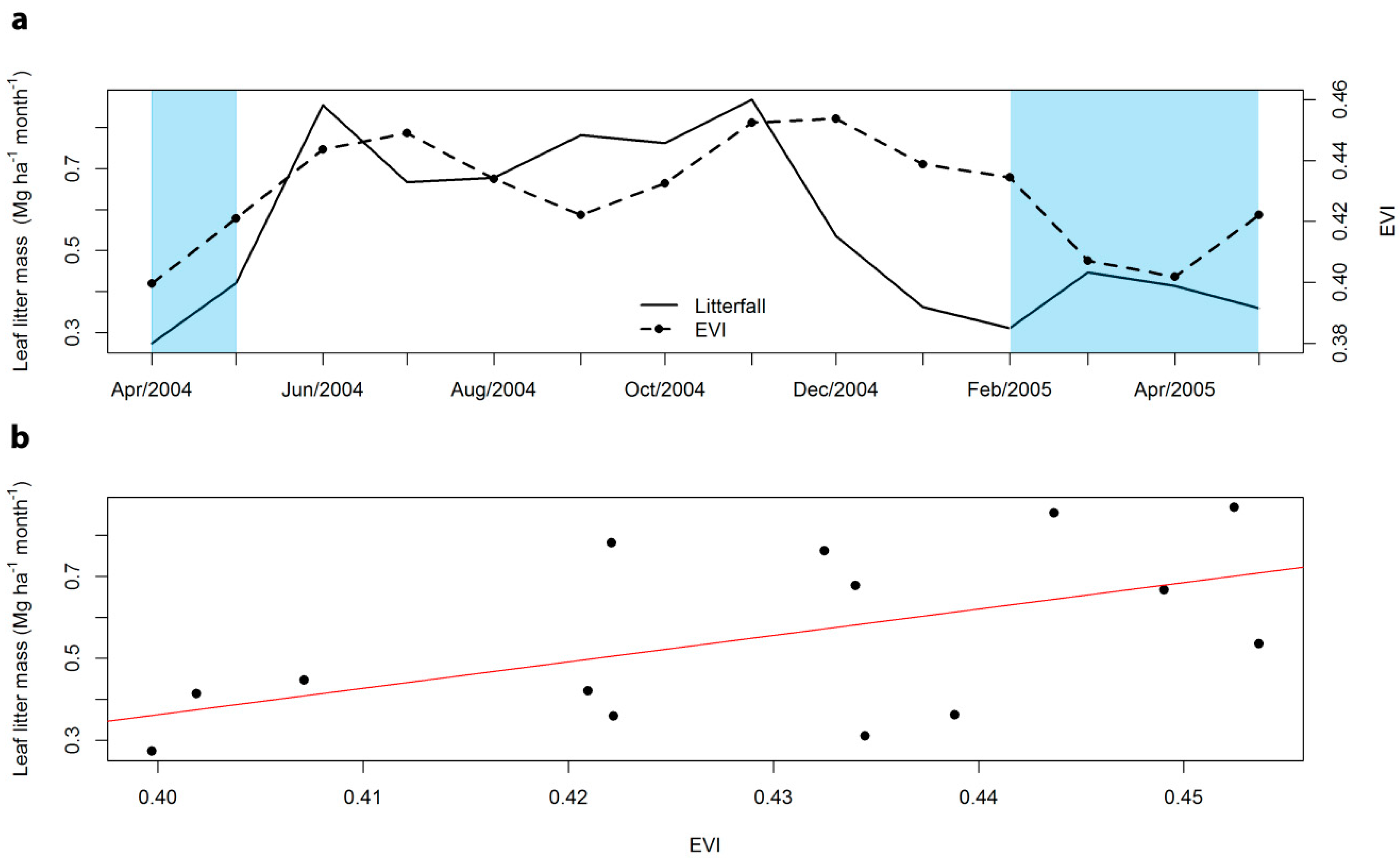
3.3. Seasonal Phenology Patterns and Analysis of Forest Canopy Gaps
3.4. Inter-Annual Variation of Seasonal Drivers and EVI-Multi-Angle Implementation Correction (MAIAC)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Espírito-Santo, F.D.B.; Gloor, M.; Keller, M.; Malhi, Y.; Saatchi, S.; Nelson, B.; Junior, R.C.O.; Pereira, C.; Lloyd, J.; Frolking, S.; et al. Size and frequency of natural forest disturbances and the Amazon forest carbon balance. Nat. Commun. 2014, 5, 3434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, O.L.; Aragão, L.E.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; López-gonzález, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A.; et al. Drought sensitivity of the Amazon rainforest. Science 2009, 323, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Brienen, R.J.W.; Phillips, O.L.; Feldpausch, T.R.; Gloor, E.; Baker, T.R.; Lloyd, J.; Lopez-Gonzalez, G.; Monteagudo-Mendoza, A.; Malhi, Y.; Lewis, S.L.; et al. Long-term decline of the Amazon carbon sink. Nature 2015, 519, 344–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldpausch, T.R.; Phillips, O.L.; Brienen, R.J.W.; Gloor, E.; Lloyd, J.; Malhi, Y.; Alarcón, A.; Dávila, E.Á.; Andrade, A.; Aragao, L.E.O.C.; et al. Amazon forest response to repeated droughts. Glob. Biogeochem. Cycles 2016, 30, 964–982. [Google Scholar] [CrossRef]
- Gatti, L.V.; Gloor, M.; Miller, J.B.; Doughty, C.E.; Malhi, Y.; Domingues, L.G.; Basso, L.S.; Martinewski, A.; Correia, C.S.C.; Borges, V.F.; et al. Drought sensitivity of Amazonian carbon balance revealed by atmospheric measurements. Nature 2014, 506, 76–80. [Google Scholar] [CrossRef]
- Restrepo-Coupe, N.; da Rocha, H.R.; Hutyra, L.R.; da Araujo, A.C.; Borma, L.S.; Christoffersen, B.; Cabral, O.M.R.; de Camargo, P.B.; Cardoso, F.L.; da Costa, A.C.L.; et al. What drives the seasonality of photosynthesis across the Amazon basin? A cross-site analysis of eddy flux tower measurements from the Brasil flux network. Agric. For. Meteorol. 2013, 182, 128–144. [Google Scholar] [CrossRef]
- Wagner, F.H.; Hérault, B.; Rossi, V.; Hilker, T.; Maeda, E.E.; Sanchez, A.; Lyapustin, A.I.; Galvão, L.S.; Wang, Y.; Aragão, L.E.O.C. Climate drivers of the Amazon forest greening. PLoS ONE 2017, 12, e0180932. [Google Scholar] [CrossRef]
- Chave, J.; Navarrete, D.; Almeida, S.; Álvarez, E.; Aragão, L.E.O.C.; Bonal, D.; Châtelet, P.; Silva-Espejo, J.E.; Goret, J.-Y.; von Hildebrand, P.; et al. Regional and seasonal patterns of litterfall in tropical South America. Biogeosciences 2010, 7, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Albert, L.P.; Lopes, A.P.; Restrepo-Coupe, N.; Hayek, M.; Wiedemann, K.T.; Guan, K.; Stark, S.C.; Christoffersen, B.; Prohaska, N.; et al. Leaf development and demography explain photosynthetic seasonality in Amazon evergreen forests. Science 2016, 351, 972–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Kobayashi, H.; Stark, S.C.; Meng, R.; Guan, K.; Tran, N.N.; Gao, S.; Yang, W.; Restrepo-Coupe, N.; Miura, T.; et al. Biological processes dominate seasonality of remotely sensed canopy greenness in an Amazon evergreen forest. New Phytol. 2018, 217, 1507–1520. [Google Scholar] [CrossRef] [PubMed]
- Huete, A.R.; Didan, K.; Shimabukuro, Y.E.; Ratana, P.; Saleska, S.R.; Hutyra, L.R.; Yang, W.; Nemani, R.R.; Myneni, R. Amazon rainforests green-up with sunlight in dry season. Geophys. Res. Lett. 2006, 33, 2–5. [Google Scholar] [CrossRef]
- Saleska, S.R.; Didan, K.; Huete, A.R.; da Rocha, H.R. BREVIA Amazon Forests Green-Up During 2005 Drought. Science 2007, 318, 612. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.O.; Malhi, Y.; Aragão, L.E.; Ladle, R.; Arai, E.; Barbier, N.; Phillips, O.; Anderson, L.O.; Ladle, R.; Arai, E.; et al. Remote sensing detection in Amazonian of droughts forest canopies. New Phytol. 2010, 187, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Brando, P.M.; Goetz, S.J.; Baccini, A.; Nepstad, D.C.; Beck, P.S.A.; Christman, M.C. Seasonal and interannual variability of climate and vegetation indices across the Amazon. Proc. Natl. Acad. Sci. USA 2010, 107, 14685–14690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvão, L.S.; dos Santos, J.R.; Roberts, D.A.; Breunig, F.M.; Toomey, M.; de Moura, Y.M. On intra-annual EVI variability in the dry season of tropical forest: A case study with MODIS and hyperspectral data. Remote Sens. Environ. 2011, 115, 2350–2359. [Google Scholar] [CrossRef]
- Morton, D.C.; Rubio, J.; Cook, B.D.; Gastellu-Etchegorry, J.-P.; Longo, M.; Choi, H.; Hunter, M.O.; Keller, M. Amazon forest structure generates diurnal and seasonal variability in light utilization. Biogeosci. Discuss. 2016, 12, 19043–19072. [Google Scholar] [CrossRef]
- Soudani, K.; François, C. Remote sensing: A green illusion. Nature 2014, 506, 165–166. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Samanta, A.; Costa, M.H.; Ganguly, S.; Nemani, R.R.; Myneni, R.B. Widespread decline in greenness of Amazonian vegetation due to the 2010 drought. Geophys. Res. Lett. 2011, 38, 2–5. [Google Scholar] [CrossRef]
- Samanta, A.; Ganguly, S.; Myneni, R.B. MODIS enhanced vegetation index data do not show greening of amazon forests during the 2005 drought. New Phytol. 2011, 189, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Hilker, T.; Lyapustin, A.I.; Tucker, C.J.; Hall, F.G.; Myneni, R.B.; Wang, Y.; Bi, J.; Mendes de Moura, Y.; Sellers, P.J. Vegetation dynamics and rainfall sensitivity of the Amazon. Proc. Natl. Acad. Sci. USA 2014, 111, 16041–16046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyapustin, A.I.; Wang, Y.; Laszlo, I.; Hilker, T.; Hall, F.G.; Sellers, P.J.; Tucker, C.J.; Korkin, S.V. Multi-angle implementation of atmospheric correction for MODIS (MAIAC): 3. Atmospheric correction. Remote Sens. Environ. 2012, 127, 385–393. [Google Scholar] [CrossRef]
- Lopes, A.; Walker, B.; Wu, J.; Maurício, P.; De Alencastro, L.; Valentim, J.; Prohaska, N.; Augusto, G.; Saleska, S.R. Remote Sensing of Environment Leaf fl ush drives dry season green-up of the Central Amazon. Remote Sens. Environ. 2016, 182, 90–98. [Google Scholar] [CrossRef]
- Maeda, E.E.; Moura, Y.M.; Wagner, F.; Hilker, T.; Lyapustin, A.I.; Wang, Y.; Chave, J.; Mõttus, M.; Aragão, L.E.O.C.; Shimabukuro, Y. Consistency of vegetation index seasonality across the Amazon rainforest. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 42–53. [Google Scholar] [CrossRef]
- de Moura, Y.M.; Hilker, T.; Lyapustin, A.I.; Galvão, L.S.; dos Santos, J.R.; Anderson, L.O.; de Sousa, C.H.R.; Arai, E. Seasonality and drought effects of Amazonian forests observed from multi-angle satellite data. Remote Sens. Environ. 2015, 171, 278–290. [Google Scholar] [CrossRef]
- Maeda, E.E.; Ma, X.; Wagner, F.H.; Kim, H.; Oki, T.; Eamus, D.; Huete, A. Evapotranspiration seasonality across the Amazon Basin. Earth Syst. Dyn. 2017, 8, 439–454. [Google Scholar] [CrossRef] [Green Version]
- Hess, L.L.; Melack, J.M.; Novo, E.M.L.M.; Barbosa, C.C.F.; Gastil, M. Dual-season mapping of wetland inundation and vegetation for the central Amazon basin. Remote Sens. Environ. 2003, 87, 404–428. [Google Scholar] [CrossRef]
- Junk, W.J.; Piedade, M.T.F.; Wittmann, F.; Schöngart, J.; Parolin, P. Amazonian Floodplain Forests; 2011; Volume 53, ISBN 9788578110796. [Google Scholar]
- De Simone, O.; Junk, W.J.; Schmidt, W. Central Amazon floodplain forests: Root adaptations to prolonged flooding. Russ. J. Plant Physiol. 2003, 50, 848–855. [Google Scholar] [CrossRef]
- Parolin, P.; De Simone, O.; Haase, K.; Waldhoff, D. Central Amazonian floodplain forests: Tree adaptations in a pulsing system. Bot. Rev. 2004, 70, 357–380. [Google Scholar] [CrossRef]
- Parolin, P.; Lucas, C.; Piedade, M.T.F.; Wittmann, F. Drought responses of flood-tolerant trees in Amazonian floodplains. Ann. Bot. 2010, 105, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Finlayson, M.; Lévêque, C.; Randy Milton, G.; Peterson, G.; Pritchard, D.; Ratner, B.D.; Reid, W.V.; Revenga, C.; Rivera, M.; Schutyser, F.; et al. A Report of the Millennium Ecosystem Assessment. Available online: https://www.millenniumassessment.org/documents/document.356.aspx.pdf. (accessed on 20 February 2019).
- Schongart, J.; Piedade, M.T.F.; Horna, V.; Worbes, M. Phenology and stem-growth periodicity of tree species in Amazonian floodplain forests. J. Trop. Ecol. 2002, 18, 581–597. [Google Scholar] [CrossRef] [Green Version]
- Schongart, J.; Junk, W.J.; Piedade, M.T.F.; Ayres, J.M.; Hüttermann, A.; Worbes, M. Teleconnection between tree growth in the Amazonian floodplains and the El Niño—Southern Oscillation effect. Glob. Chang. Biol. 2004, 10, 683–692. [Google Scholar] [CrossRef]
- Teresa Fernandez Piedade, M.; Junk, W.J.; Parolin, P. The flood pulse and photosynthetic response of trees in a white water floodplain (várzea) of the Central Amazon, Brazil. SIL Proc. 1922–2010 2000, 27, 1734–1739. [Google Scholar] [CrossRef]
- Nobre, C.A.; Sampaio, G.; Borma, L.S.; Castilla-Rubio, J.C.; Silva, J.S.; Cardoso, M. Land-use and climate change risks in the Amazon and the need of a novel sustainable development paradigm. Proc. Natl. Acad. Sci. USA 2016, 113, 10759–10768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmagro, H.J.; Zanella de Arruda, P.H.; Vourlitis, G.L.; Lathuillière, M.J.; de S. Nogueira, J.; Couto, E.G.; Johnson, M.S. Radiative forcing of methane fluxes offsets net carbon dioxide uptake for a tropical flooded forest. Glob. Chang. Biol. 2019, 4, 1967–1981. [Google Scholar] [CrossRef] [PubMed]
- de Resende, A.F.; Schöngart, J.; Streher, A.S.; Ferreira-Ferreira, J.; Piedade, M.T.F.; Silva, T.S.F. Massive tree mortality from flood pulse disturbances in Amazonian floodplain forests: The collateral effects of hydropower production. Sci. Total Environ. 2019, 659, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Mesquita, R.C.G.; Flores, B.M.; Holmgren, M.; Jakovac, C.C.; Xu, C.; van Nes, E.H. Floodplains as an Achilles’ heel of Amazonian forest resilience. Proc. Natl. Acad. Sci. USA 2017, 114, 4442–4446. [Google Scholar]
- Malhi, Y.; Wright, J. Spatial patterns and recent trends in the climate of tropical rainforest regions. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 311–329. [Google Scholar] [CrossRef]
- Borma, L.S.; Da Rocha, H.R.; Cabral, O.M.; Von Randow, C.; Collicchio, E.; Kurzatkowski, D.; Brugger, P.J.; Freitas, H.; Tannus, R.; Oliveira, L.; et al. Atmosphere and hydrological controls of the evapotranspiration over a floodplain forest in the Bananal Island region, Amazonia. J. Geophys. Res. Biogeosci. 2009, 114, G01003. [Google Scholar] [CrossRef]
- Costa, G.P. Fluxos de energia, CO2 e CH4 Sobre a Floresta em Planície de Inundação da Ilha do Bananal. Ph.D. Dissertation, Federal University of São Paulo, São Paulo, Brazil, 2015. Available online: http://www.teses.usp.br/teses/disponiveis/91/91131/tde-28092015-111609/ (accessed on 15 October 2018).
- Homeier, J.; Kurzatkowski, D.; Leuschner, C. Stand dynamics of the drought-affected floodplain forests of Araguaia River, Brazilian Amazon. For. Ecosyst. 2017, 4, 10. [Google Scholar] [CrossRef]
- Da Rocha, H.R.; Manzi, A.O.; Cabral, O.M.; Miller, S.D.; Goulden, M.L.; Saleska, S.R.; Coupe, N.R.; Wofsy, S.C.; Borma, L.S.; Artaxo, R.; et al. Patterns of water and heat flux across a biome gradient from tropical forest to savanna in brazil. J. Geophys. Res. Biogeosci. 2009, 114, 1–8. [Google Scholar] [CrossRef]
- Hayek, N.H.; Longo, M.; Wu, J.; Smith, M.N.; Restrepo-coupe, N.; Tapajós, R.; Da Silva, R.; Fitzjarrald, D.R.; Camargo, P.B.; Hutyra, L.R.; et al. Carbon exchange in an Amazon forest: From hours to years. Biogeosciences 2018, 15, 4833–4848. [Google Scholar] [CrossRef]
- Cabral, O.M.R.; Gash, J.H.C.; Rocha, H.R.; Marsden, C.; Ligo, M.A.V.; Freitas, H.C.; Tatsch, J.D.; Gomes, E. Fluxes of CO2 above a plantation of Eucalyptus in southeast Brazil. Agric. For. Meteorol. 2011, 151, 49–59. [Google Scholar] [CrossRef]
- Hutyra, L.R.; Munger, J.W.; Saleska, S.R.; Gottlieb, E.; Daube, B.C.; Dunn, A.L.; Amaral, D.F.; de Camargo, P.B.; Wofsy, S.C. Seasonal controls on the exchange of carbon and water in an Amazonian rain forest. J. Geophys. Res. Biogeosci. 2007, 112. [Google Scholar] [CrossRef]
- Moreira, K.S.; Rocha, H.R.; Kurzatkowski, D.; Ribeiro da Mata, R.; Pinto, A.S. Avaliação na Queda de Liteira em Ecótonos no Entorno da Ilha do Bananal. In Proceedings of the 2nd Congress of Students and Scholars of the LBA Experiment, Manaus, Brazil, 1–13 July 2005; Available online: http://lba2.inpa.gov.br/lbaconferencias/2005_lba_student_conf/index.htm (accessed on 12 February 2019).
- Dalagnol, R.; Wagner, F.H.; Galvão, L.S.; Nelson, B.W.; Oliveira, E. Life cycle of bamboo in southwestern Amazon and its relation to fire events. Biogeosciences 2018, 15, 6087–6104. [Google Scholar] [CrossRef]
- Dalagnol, R.; Wagner, F.H.; Galvão, L.S.; Aragão, L.E.O.eC. The MANVI Product: MODIS (MAIAC) Nadir-Solar Adjusted Vegetation Indices (EVI and NDVI) for South America, Version v1; Zenodo, 2019. Available online: https://0-doi-org.brum.beds.ac.uk/10.5281/zenodo.3159488 (accessed on 25 June 2019).
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.; Gao, X.; Ferreira, L. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 850, 2011–2014. [Google Scholar] [CrossRef]
- Dee, D.P.; Uppala, S.M.; Simmons, A.J.; Berrisford, P.; Poli, P.; Kobayashi, S.; Andrae, U.; Balmaseda, M.A.; Balsamo, G.; Bauer, P.; et al. The ERA-Interim reanalysis: Configuration and performance of the data assimilation system. Q. J. R. Meteorol. Soc. 2011, 137, 553–597. [Google Scholar] [CrossRef]
- Aragão; Luiz Eduardo, O.C.; Malhi, Y.; Roman-Cuesta, R.M.; Saatchi, S.; Anderson, L.O.; Shimabukuro, Y.E. Spatial patterns and fire response of recent Amazonian droughts. Geophys. Res. Lett. 2007, 34, 1–5. [Google Scholar] [CrossRef]
- Hunter, M.O.; Keller, M.; Morton, D.; Cook, B.; Lefsky, M.; Ducey, M.; Saleska, S.; De Oliveira, R.C.; Schietti, J.; Zang, R. Structural dynamics of tropical moist forest gaps. PLoS ONE 2015, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tavares, I.B.; Borma, L.S.; Fonseca, L.D.M.; Collicchio, E.; Domingues, T.F.; Rocha, H.R. The growth pattern of the forest located in a southeast Amazonian floodplain during the 2015/2016 ENSO year. Ecohydrology 2019. under review. [Google Scholar]
- Gloor, M.; Barichivich, J.; Ziv, G.; Brienen, R.; Schöngart, J.; Peylin, P. Recent Amazon climate as background for possible ongoing Special Section. Glob. Biogeochem. Cycles 2015, 29, 1384–1399. [Google Scholar] [CrossRef]
- Jiménez-Muñoz, J.C.; Mattar, C.; Barichivich, J.; Santamaría-Artigas, A.; Takahashi, K.; Malhi, Y.; Sobrino, J.A.; Schrier, G. Van Der Record-breaking warming and extreme drought in the Amazon rainforest during the course of El Niño 2015–2016. Sci. Rep. 2016, 6, 33130. [Google Scholar] [CrossRef] [PubMed]
- Schöngart, J.; Haugaasen, T.; Wittmann, F.; Piedade, M.T.F.; Bredin, Y.K.; de Assis, R.L.; Nobre Quesada, C.A. Above-ground woody biomass distribution in Amazonian floodplain forests: Effects of hydroperiod and substrate properties. For. Ecol. Manag. 2018, 432, 365–375. [Google Scholar]
- Piedade, M.T.F.; Schöngart, J.; Wittmann, F.; Parolin, P.; Junk, W.J. Impactos ecológicos da inundação e seca na vegetação de áreas alagáveis amazônicas. In Eventos Climáticos Extremos na Amazônia Causas e Conseqüências; Oficina de Textos: São Paulo, Brazil, 2013; pp. 268–304. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Acquisitions | Start Year | End Year | Usage |
|---|---|---|---|---|
| LE | Tower | 2004 | 2014 | ET computation |
| PAR | Tower | 2011 | 2013 | NEE computation |
| Press | Tower | 2004 | 2014 | Relative humidity (RH) computation |
| q | Tower | 2004 | 2016 | RH and VPD computation |
| Rn | Tower | 2004 | 2014 | Correlation variable |
| GPP | Tower | 2011 | 2013 | Productivity estimate/Correlation variable |
| ET | Tower | 2004 | 2014 | Correlation variable |
| VPD | Tower/Satellite | 2004 | 2016 | Correlation variable |
| Ta | Tower/Satellite | 2004 | 2016 | VPD and ET computation/Correlation variable |
| Rainfall/TRMM | Tower/Satellite | 2004 | 2014 | Correlation variable |
| CWD | Tower/Satellite | 2004 | 2016 | Correlation variable |
| EVI | Satellite | 2004 | 2016 | Phenology and productivity proxy/Correlation variable |
| Soil moisture | Field | 2014 | 2016 | Correlation variable |
| Litterfall | Field | 2004 | 2005 | Phenology proxy/Correlation variable |
| Flood height | Tower | 2004 | 2016 | Define seasonal flooding |
| Plots | Number of Individuals | Tree Mean Height (m) | 5% Percentile (m) | 95% Percentile (m) | Maximum (m) |
|---|---|---|---|---|---|
| BAN1 | 86 | 11.79 | 6.84 | 18.86 | 28.36 |
| BAN2 | 84 | 12.48 | 4.5 | 19.77 | 38.89 |
| LiDAR | - | 10.2 | 4.8 | 14.9 | 38 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonseca, L.D.M.; Dalagnol, R.; Malhi, Y.; Rifai, S.W.; Costa, G.B.; Silva, T.S.F.; Da Rocha, H.R.; Tavares, I.B.; Borma, L.S. Phenology and Seasonal Ecosystem Productivity in an Amazonian Floodplain Forest. Remote Sens. 2019, 11, 1530. https://0-doi-org.brum.beds.ac.uk/10.3390/rs11131530
Fonseca LDM, Dalagnol R, Malhi Y, Rifai SW, Costa GB, Silva TSF, Da Rocha HR, Tavares IB, Borma LS. Phenology and Seasonal Ecosystem Productivity in an Amazonian Floodplain Forest. Remote Sensing. 2019; 11(13):1530. https://0-doi-org.brum.beds.ac.uk/10.3390/rs11131530
Chicago/Turabian StyleFonseca, Letícia D. M., Ricardo Dalagnol, Yadvinder Malhi, Sami W. Rifai, Gabriel B. Costa, Thiago S. F. Silva, Humberto R. Da Rocha, Iane B. Tavares, and Laura S. Borma. 2019. "Phenology and Seasonal Ecosystem Productivity in an Amazonian Floodplain Forest" Remote Sensing 11, no. 13: 1530. https://0-doi-org.brum.beds.ac.uk/10.3390/rs11131530