
Remotely Sensed Spatiotemporal Variation in Crude Protein of Shortgrass Steppe Forage

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
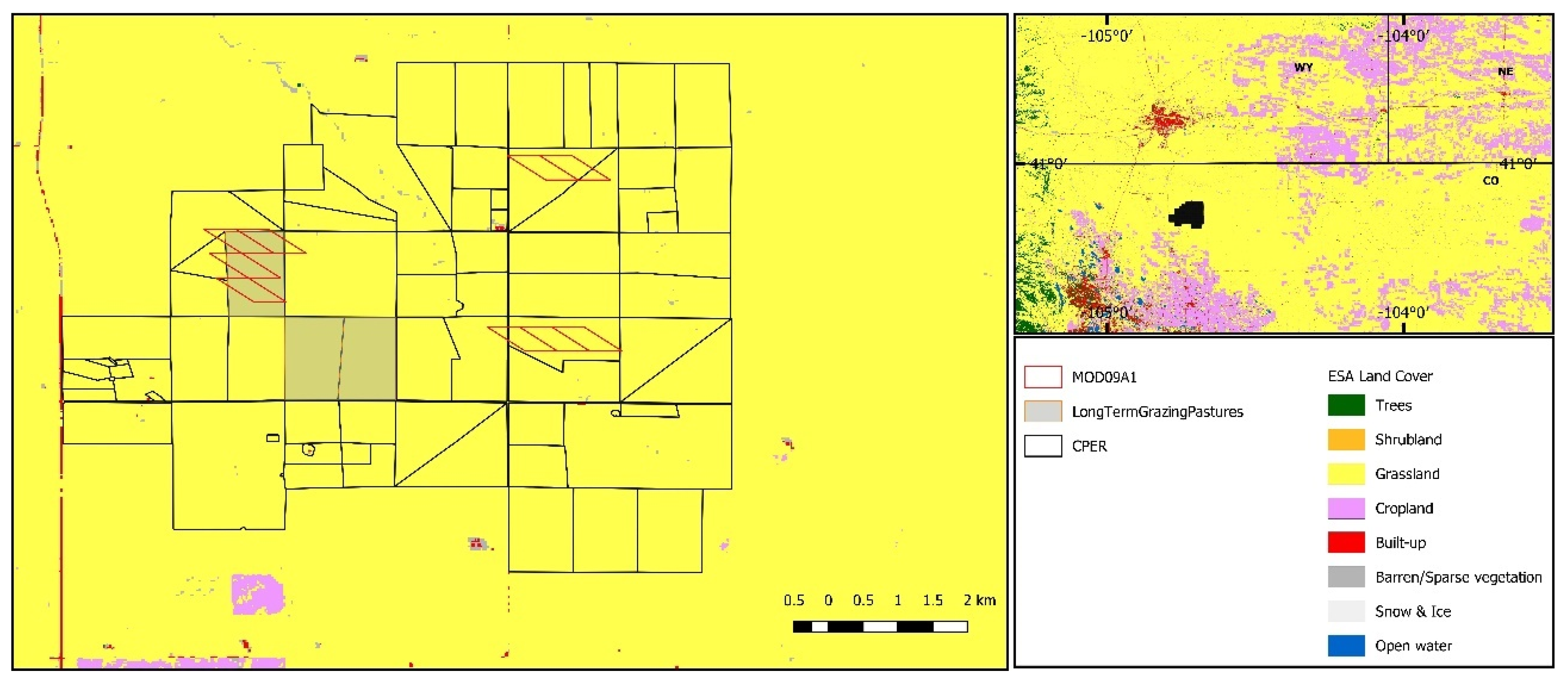
2.1. Site Description
2.2. Model Calibration
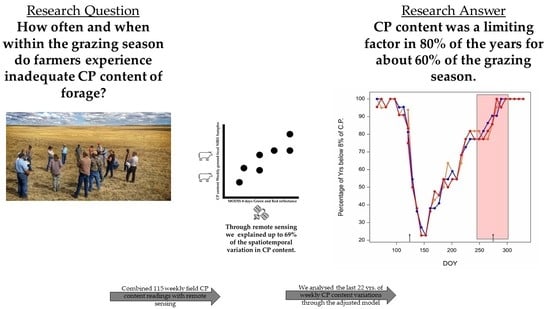
2.3. CP Content Risk Assessment
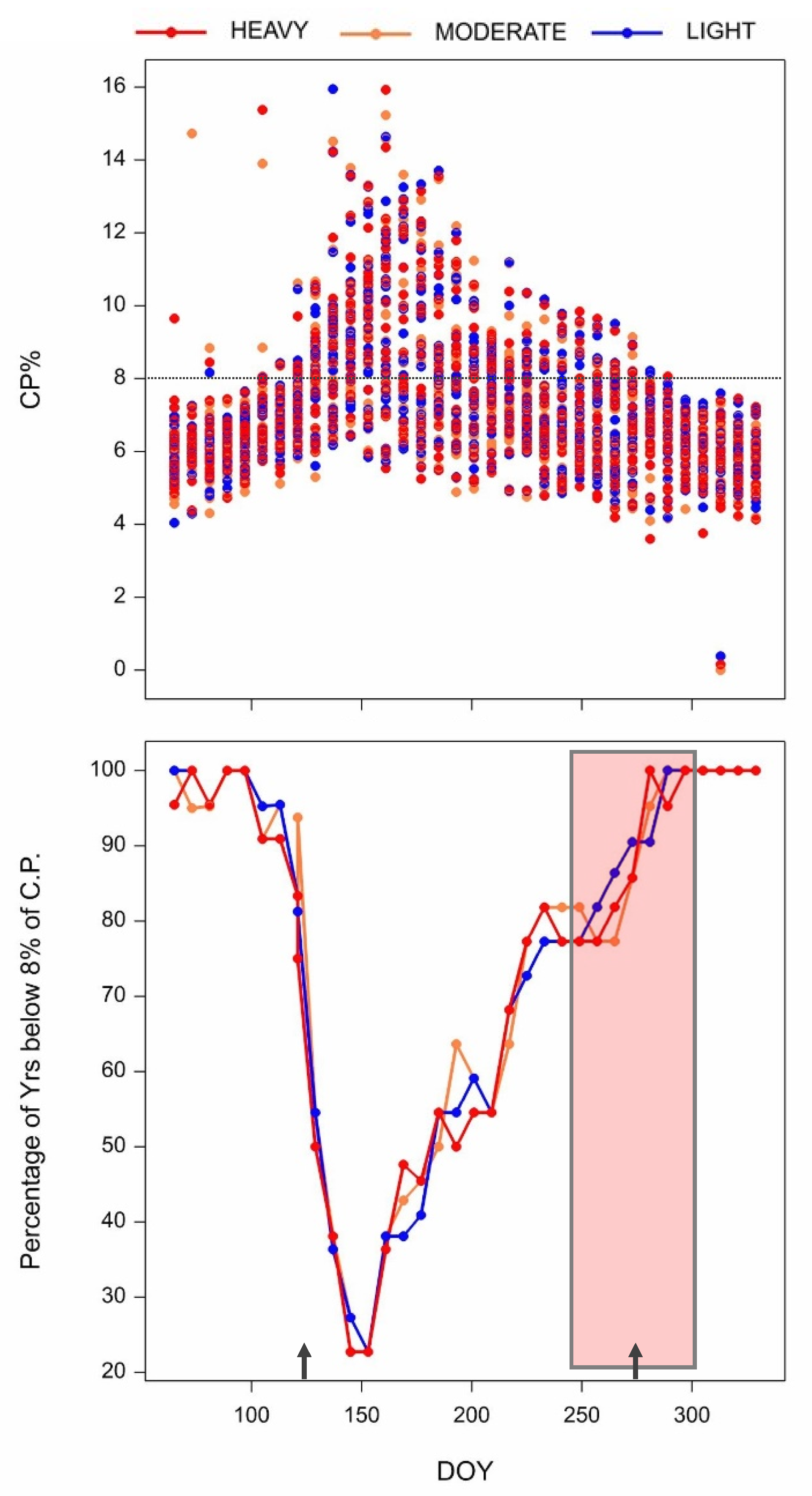
3. Results
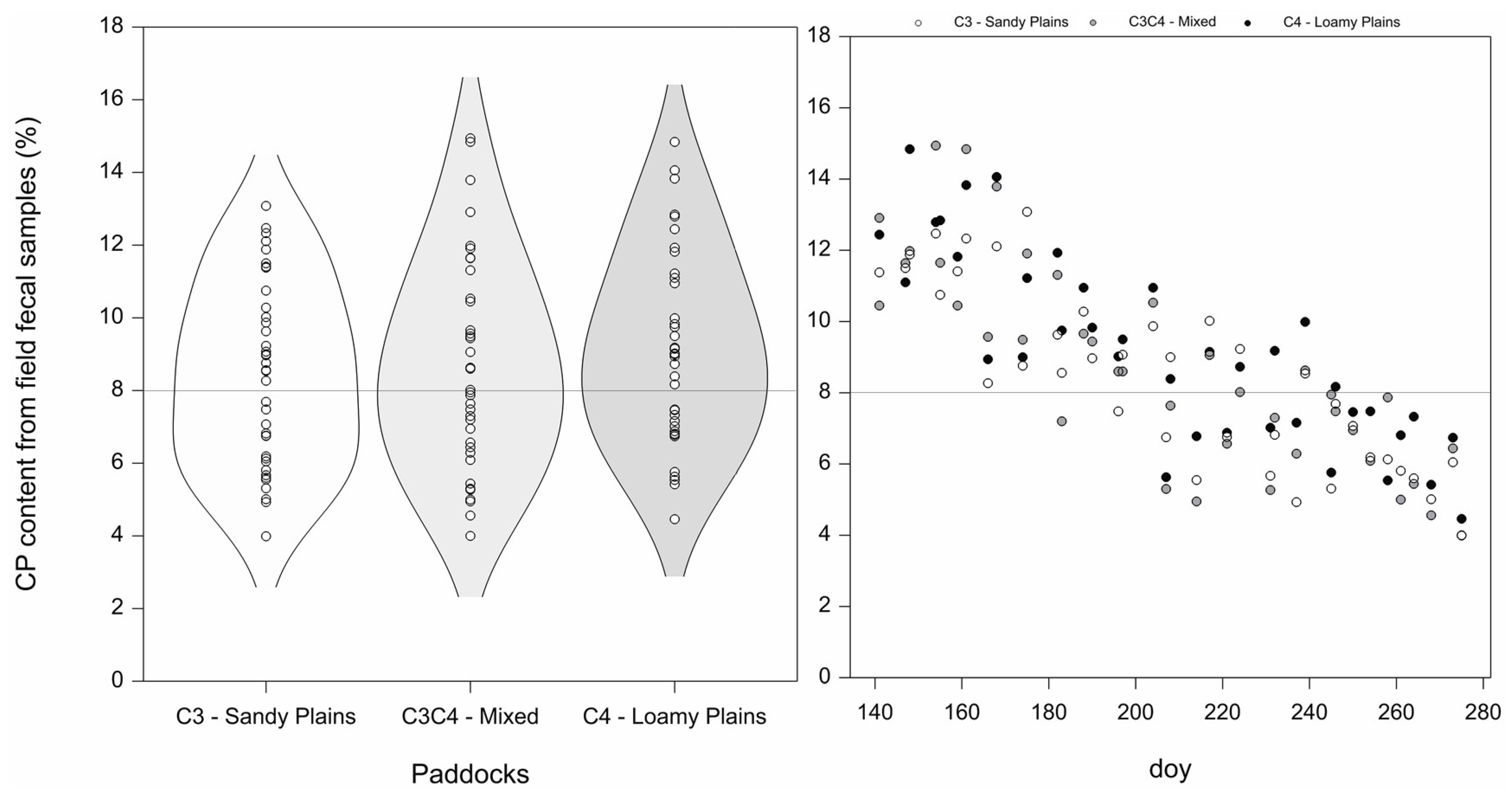
3.1. Variability in Field Estimates of CP Content
3.2. Crude Protein Content Calibration with Remotely Sensed Data
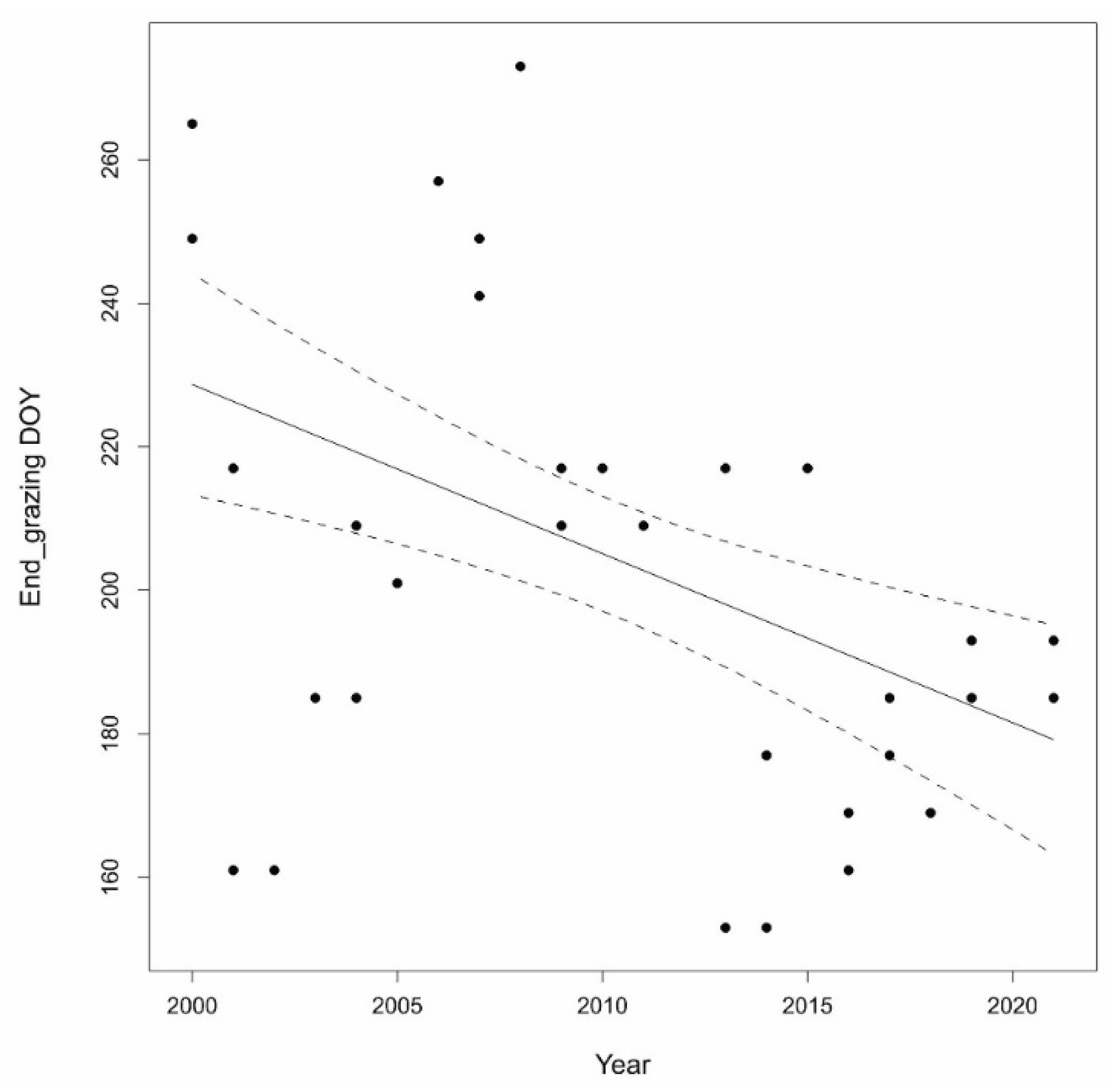
3.3. Risk Assessment of the Lack of Crude Protein Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holechek, J.L.; Vavra, M.; Arthijn, D. Relationships between performance, intake, diet nutritive quality and fecal nutritive quality of cattle on mountain range. J. Range Manag. 1978, 741–744. [Google Scholar] [CrossRef] [Green Version]
- Holechek, J.L. An approach for setting the stocking rate. Area 1988, 10, 10–14. [Google Scholar]
- Hart, R.H.; Waggoner, J.W.; Dunn, T.G.; Kaltenbach, C.C.; Adams, L.D. Optimal stocking rate for cow-calf enterprises on native range and complementary improved pastures. J. Range Manag. 1988, 41, 435. [Google Scholar] [CrossRef]
- Derner, J.D.; Hart, R.H.; Smith, M.A.; Waggoner, J.W. Long-term cattle gain responses to stocking rate and grazing systems in northern mixed-grass prairie. Livest. Sci. 2008, 117, 60–69. [Google Scholar] [CrossRef]
- Bement, R.E. A stocking-rate guide for beef production on Blue-Grama range. J. Range Manag. 1969, 22, 83. [Google Scholar] [CrossRef] [Green Version]
- Raynor, E.J.; Derner, J.D.; Baldwin, T.; Ritten, J.P.; Augustine, D.J. Multidecadal directional shift in shortgrass stocking rates. Rangel. Ecol. Manag. 2021, 74, 72–80. [Google Scholar] [CrossRef]
- Wilmer, H.; Augustine, D.J.; Derner, J.D.; Fernández-Giménez, M.E.; Briske, D.D.; Roche, L.M.; Tate, K.W.; Miller, K.E. Diverse management strategies produce similar ecological outcomes on ranches in western Great Plains: Social-ecological assessment. Rangel. Ecol. Manag. 2018, 71, 626–636. [Google Scholar] [CrossRef]
- Raynor, E.J.; Derner, J.D.; Hoover, D.L.; Parton, W.J.; Augustine, D.J. Large-scale and local climatic controls on large herbivore productivity: Implications for adaptive rangeland management. Ecol. Appl. 2019, 30, e02053. [Google Scholar] [CrossRef]
- Reeves, J.L.; Derner, J.D.; Sanderson, M.A.; Hendrickson, J.R.; Kronberg, S.L.; Petersen, M.K.; Vermeire, L.T. Seasonal weather influences on yearling beef steer production in C3-dominated Northern Great Plains rangeland. Agric. Ecosyst. Environ. 2014, 183, 110–117. [Google Scholar] [CrossRef]
- Reeves, J.L.; Derner, J.D.; Sanderson, M.A.; Petersen, M.K.; Vermeire, L.T.; Hendrickson, J.R.; Kronberg, S.L. Temperature and precipitation affect steer weight gains differentially by stocking rate in northern Mixed-Grass Prairie. Rangel. Ecol. Manag. 2013, 66, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Dunn, B.H.; Smart, A.J.; Gates, R.N.; Johnson, P.S.; Beutler, M.K.; Diersen, M.A.; Janssen, L.L. Long-term production and profitability from grazing cattle in the northern mixed grass prairie. Rangel. Ecol. Manag. 2010, 63, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, T.W.; Ritten, J.P.; Bastian, C.T.; Derner, J.D.; Tanaka, J.A. Economic impacts of increasing seasonal precipitation variation on southeast Wyoming cow-calf enterprises. Rangel. Ecol. Manag. 2016, 69, 465–473. [Google Scholar] [CrossRef]
- Irisarri, J.G.; Derner, J.D.; Ritten, J.P.; Peck, D.E. Beef production and net revenue variability from grazing systems on semiarid grasslands of North America. Livest. Sci. 2019, 220, 93–99. [Google Scholar] [CrossRef]
- Ritten, J.P.; Frasier, W.M.; Bastian, C.T.; Gray, S.T. Optimal rangeland stocking decisions under stochastic and climate-impacted weather. Am. J. Agric. Econ. 2010, 92, 1242–1255. [Google Scholar] [CrossRef]
- Augustine, D.J.; Derner, J.D.; Fernández-Giménez, M.E.; Porensky, L.M.; Wilmer, H.; Briske, D.D. Adaptive, multipaddock rotational grazing management: A ranch-scale assessment of effects on vegetation and livestock performance in semiarid rangeland. Rangel. Ecol. Manag. 2020, 73, 796–810. [Google Scholar] [CrossRef]
- Dieguez Cameroni, F.; Fort, H. Towards scientifically based management of extensive livestock farming in terms of ecological predator-prey modeling. Agric. Syst. 2017, 153, 127–137. [Google Scholar] [CrossRef]
- Russell, M.L.; Bailey, D.W.; Thomas, M.G.; Witmore, B.K. Grazing distribution and diet quality of Angus, Brangus, and Brahman cows in the Chihuahuan desert. Rangel. Ecol. Manag. 2012, 65, 371–381. [Google Scholar] [CrossRef]
- Tolleson, D.R.; Schafer, D.W. Application of fecal near-infrared spectroscopy and nutritional balance software to monitor diet quality and body condition in beef cows grazing Arizona rangeland. J. Anim. Sci. 2014, 92, 349–358. [Google Scholar] [CrossRef]
- Augustine, D.J.; Blumenthal, D.M.; Springer, T.L.; Lecain, D.R.; Gunter, S.A.; Derner, J.D. Elevated CO2 induces substantial and persistent declines in forage quality irrespective of warming in mixedgrass prairie. Ecol. Appl. 2018, 28, 721–735. [Google Scholar] [CrossRef]
- Gaffney, R.; Porensky, L.M.; Gao, F.; Irisarri, J.G.; Derner, J.D.; Augustine, D.J. Using APAR to predict aboveground plant productivity in semi-arid rangelands: Spatial and temporal relationships differ. Remote Sens. 2018, 10, 1474. [Google Scholar] [CrossRef] [Green Version]
- Grigera, G.; Oesterheld, M.; Pacín, F.; Pacin, F. Monitoring forage production for farmers’ decision making. Agric. Syst. 2007, 94, 637–648. [Google Scholar] [CrossRef]
- Reeves, M.C.; Moreno, A.L.; Bagne, K.E.; Running, S.W. Estimating climate change effects on net primary production of rangelands in the United States. Clim. Change 2014, 126, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Reeves, M.C.; Zhao, M.; Running, S.W. Special Feature: Applications of geospatial techniques applying improved estimates of MODIS productivity to characterize grassland vegetation dynamics. Earth Sci. 2006, 59, 1–10. [Google Scholar]
- Reeves, M.C.; Winslow, J.C.; Running, S.W. Mapping weekly rangeland vegetation productivity using MODIS algorithms. J. Range Man 2001, 54, 90–105. [Google Scholar]
- Mathis, C. Protein and Energy Supplementation to Beef Cows Grazing New Mexico Rangelands. Cooperative Extension Service Circular 564; College of Agriculture and Home Economics. 2003. Available online: https://aces.nmsu.edu/pubs/_circulars/Circ564.pdf (accessed on 10 January 2022).
- Mathis, C.P.; Sawyer, J.E. Nutritional management of grazing beef cows. Vet. Clin. N. Am. Food Anim. Pract. 2007, 23, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Heldt, J.S.; Cochran, R.C.; Mathis, C.P.; Woods, B.C.; Olson, K.C.; Titgemeyer, E.C.; Nagaraja, T.G.; Vanzant, E.S.; Johnson, D.E. Effects of level and source of carbohydrate and level of degradable intake protein on intake and digestion of low-quality tallgrass-prairie hay by beef steers. J. Anim. Sci. 1999, 77, 2846–2854. [Google Scholar] [CrossRef]
- Mathis, C.P.; Cochran, R.C.; Heldt, J.S.; Woods, B.C.; Abdelgadir, I.E.O.; Olson, K.C.; Titgemeyer, E.C.; Vanzant, E.S. Effects of supplemental degradable intake protein on utilization of medium- to low-quality forages. J. Anim. Sci. 2000, 78, 224–232. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Olson, K.C.; Tolleson, D. Climate change and cattle nutritional stress. Glob. Change Biol. 2010, 16, 2901–2911. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.; Angerer, J.P. Long-term declines in dietary nutritional quality for North American cattle. Environ. Res. Lett. 2017, 12, 044019. [Google Scholar] [CrossRef] [Green Version]
- Starks, P.J.; Zhao, D.; Phillips, W.A.; Coleman, S.W. Development of canopy reflectance algorithms for real-time prediction of bermudagrass pasture biomass and nutritive values. Crop Sci. 2006, 46, 927–934. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Chen, C. Using remote sensing to estimate forage biomass and nutrient contents at different growth stages. Biomass Bioenergy 2018, 115, 74–81. [Google Scholar] [CrossRef]
- Homolova, L.; Schaepman, M.E.; Lamarque, P.; Clevers, J.G.P.W.; De Bello, F.; Thuiller, W.; Lavorel, S. Comparison of remote sensing and plant trait-based modelling to predict ecosystem services in subalpine grasslands. Ecosphere 2014, 5, 1–29. [Google Scholar] [CrossRef]
- Lugassi, R.; Zaady, E.; Goldshleger, N.; Shoshany, M.; Chudnovsky, A. Spatial and temporal monitoring of pasture ecological quality: Sentinel-2-based estimation of crude protein and neutral detergent fiber contents. Remote Sens. 2019, 11, 799. [Google Scholar] [CrossRef] [Green Version]
- Ling, B.; Goodin, D.G.; Raynor, E.J.; Joern, A. Hyperspectral analysis of leaf pigments and nutritional elements in tallgrass prairie vegetation. Front. Plant Sci. 2019, 10, 142. [Google Scholar] [CrossRef] [Green Version]
- Ferner, J.; Linstädter, A.; Rogass, C.; Südekum, K.H.; Schmidtlein, S. Towards forage resource monitoring in subtropical savanna grasslands: Going multispectral or hyperspectral? Eur. J. Remote Sens. 2021, 54, 364–384. [Google Scholar] [CrossRef]
- Jennewein, J.S.; Eitel, J.U.H.; Joly, K.; Long, R.A.; Maguire, A.J.; Vierling, L.A.; Weygint, W. Estimating integrated measures of forage quality for herbivores by fusing optical and structural remote sensing data. Environ. Res. Lett. 2021, 16, 075006. [Google Scholar] [CrossRef]
- Watanabe, O.; Wang, S. Comparing MODIS vegetation indices with AVHRR NDVI for monitoring the forage quantity and quality in Inner Mongolia grassland, China. Grassl. Sci. 2005, 51, 33–40. [Google Scholar] [CrossRef]
- Ryan, S.J.; Cross, P.C.; Winnie, J.; Hay, C.; Bowers, J.; Getz, W.M. The utility of normalized difference vegetation index for predicting African buffalo forage quality. J. Wildl. Manag. 2012, 76, 1499–1508. [Google Scholar] [CrossRef]
- Pringle, M.J.; O’Reagain, P.J.; Stone, G.S.; Carter, J.O.; Orton, T.G.; Bushell, J.J. Using remote sensing to forecast forage quality for cattle in the dry savannas of northeast Australia. Ecol. Indic. 2021, 133, 108426. [Google Scholar] [CrossRef]
- Chabalala, Y.; Adam, E.; Oumar, Z.; Ramoelo, A. Exploiting the capabilities of Sentinel-2 and RapidEye for predicting grass nitrogen across different grass communities in a protected area. Appl. Geomat. 2020, 12, 379–395. [Google Scholar] [CrossRef]
- Diaz-Solis, H.; Kothmann, M.M.; Grant, W.E.; De Luna-Villarreal, R. Use of irrigated pastures in semi-arid grazinglands: A dynamic model for stocking rate decisions. Agric. Syst. 2006, 88, 316–331. [Google Scholar] [CrossRef]
- Díaz-Solis, H.; Kothmann, M.M.; Hamilton, W.T.; Grant, W.E. A simple ecological sustainability simulator (SESS) for stocking rate management on semi-arid grazinglands. Agric. Syst. 2003, 76, 655–680. [Google Scholar] [CrossRef]
- Díaz-Solís, H.; Grant, W.E.; Kothmann, M.M.; Teague, W.R.; Díaz-García, J.A. Adaptive management of stocking rates to reduce effects of drought on cow-calf production systems in semi-arid rangelands. Agric. Syst. 2009, 100, 43–50. [Google Scholar] [CrossRef]
- Ritten, J.P.; Bastian, C.T.; Frasier, W.M. Economically optimal stocking rates: A bioeconomic grazing model. Rangel. Ecol. Manag. 2010, 63, 407–414. [Google Scholar] [CrossRef]
- Torell, L.A.; Murugan, S.; Ramirez, O.A. Economics of flexible versus conservative stocking strategies to manage climate variability risk. Rangel. Ecol. Manag. 2010, 63, 415–425. [Google Scholar] [CrossRef]
- Natural Resources Service Conservation (Ed.) Ecological site description for Loamy plains (R067BY002CO). In Ecological Site Description Catalog; USDA: Washington, DC, USA, 2007. Available online: https://edit.jornada.nmsu.edu/catalogs/esd/067B/R067BY002CO (accessed on 10 January 2022).
- Natural Resources Service Conservation (Ed.) Ecological site description for Sandy plains (R067BY024CO). In Ecological Site Description Catalog; USDA: Washington, DC, USA, 2007. Available online: https://edit.jornada.nmsu.edu/catalogs/esd/067B/R067BY024CO (accessed on 10 January 2022).
- Reynolds, A.Q.; Derner, J.D.; Augustine, D.J.; Porensky, L.M.; Wilmer, H.; Jorns, T.; Briske, D.D.; Scasta, J.D.; Fernández-Giménez, M.E. Ecological sites: Can they be managed to promote livestock production? Rangelands 2019, 41, 239–243. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Irisarri, J.G.N.; Oesterheld, M.; Paruelo, J.M.; Texeira, M.A. Patterns and controls of above-ground net primary production in meadows of Patagonia. A remote sensing approach. J. Veg. Sci. 2012, 23, 114–126. [Google Scholar] [CrossRef]
- Pellegrini, P.; Cossani, C.M.; Bella, C.M.D.; Piñeiro, G.; Sadras, V.O.; Oesterheld, M. Simple regression models to estimate light interception in wheat crops with Sentinel-2 and a handheld sensor. Crop Sci. 2020, 60, 1607–1616. [Google Scholar] [CrossRef]
- Zanaga, D.; Van De Kerchove, R.; De Keersmaecker, W.; Souverijns, N.; Brockmann, C.; Quast, R.; Wevers, J.; Grosu, A.; Paccini, A.; Vergnaud, S.; et al. ESA WorldCover 10 m 2020 v100 2021. Available online: https://developers.google.com/earth-engine/datasets/catalog/ESA_WorldCover_v100 (accessed on 10 January 2022).
- Hart, R.H.; Ashby, M.M. Grazing intensities, vegetation, and heifer gains: 55 years on shortgrass. J. Range Manag. 1998, 51, 392–398. [Google Scholar] [CrossRef] [Green Version]
- Irisarri, J.G.N.; Derner, J.D.; Porensky, L.M.; Augustine, D.J.; Reeves, J.L.; Mueller, K.E. Grazing intensity differentially regulates ANPP response to precipitation in North American semiarid grasslands. Ecol. Appl. 2016, 26, 1370–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porensky, L.M.; Derner, J.D.; Augustine, D.J.; Milchunas, D.G. Plant community composition after 75 yr of sustained grazing intensity treatments in Shortgrass Steppe. Rangel. Ecol. Manag. 2017, 70, 456–464. [Google Scholar] [CrossRef]
- Kearney, S.P.; Porensky, L.M.; Augustine, D.J.; Derner, J.D.; Gao, F. Predicting spatial-temporal patterns of diet quality and large herbivore performance using satellite time series. Ecol. Appl. 2021, e2503. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.A.; Kaufman, Y.J.; Stark, R.; Rundquist, D. Novel algorithms for remote estimation of vegetation fraction. Remote Sens. Environ. 2002, 80, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.; Arkebauer, T.; Viña, A.; Skakun, S.; Inoue, Y. Evaluating plant photosynthetic traits via absorption coefficient in the photosynthetically active radiation region. Remote Sens. Environ. 2021, 258, 11241. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing carotenoid content in plant leaves with reflectance spectroscopy. Ratio 2002, 75, 272–281. [Google Scholar]
- Xu, D.; Wang, C.; Chen, J.; Shen, M.; Shen, B.; Yan, R.; Li, Z.; Karnieli, A.; Chen, J.; Yan, Y.; et al. The superiority of the normalized difference phenology index (NDPI) for estimating grassland aboveground fresh biomass. Remote Sens. Environ. 2021, 264, 112578. [Google Scholar] [CrossRef]
- Derner, J.D.; Hart, R.H. Grazing-induced modifications to peak standing crop in Northern Mixed-grass Prairie. Rangel. Ecol. Manag. 2007, 60, 270–276. [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Sala, O.E. Long-term forage production of North American shortgrass steppe. Ecol. Appl. 1992, 2, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Oesterheld, M.; Loreti, J.; Semmartin, M.; Sala, O.E. Inter-annual variation in primary production of a semi-arid grassland related to previous-year production. J. Veg. Sci. 2001, 12, 137. [Google Scholar] [CrossRef]
- Polley, H.W.; Briske, D.D.; Morgan, J.A.; Wolter, K.; Bailey, D.W.; Brown, J.R. Climate Change and North American Rangelands: Trends, Projections, and Implications. Rangel. Ecol. Manag. 2013, 66, 493–511. [Google Scholar] [CrossRef]
- Joyce, L.A.; Briske, D.D.; Brown, J.R.; Polley, H.W.; McCarl, B.A.; Bailey, D.W. Climate change and North American rangelands: Assessment of mitigation and adaptation strategies. Rangel. Ecol. Manag. 2013, 66, 512–528. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irisarri, J.G.N.; Durante, M.; Derner, J.D.; Oesterheld, M.; Augustine, D.J. Remotely Sensed Spatiotemporal Variation in Crude Protein of Shortgrass Steppe Forage. Remote Sens. 2022, 14, 854. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14040854
Irisarri JGN, Durante M, Derner JD, Oesterheld M, Augustine DJ. Remotely Sensed Spatiotemporal Variation in Crude Protein of Shortgrass Steppe Forage. Remote Sensing. 2022; 14(4):854. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14040854
Chicago/Turabian StyleIrisarri, Jorge Gonzalo N., Martin Durante, Justin D. Derner, Martin Oesterheld, and David J. Augustine. 2022. "Remotely Sensed Spatiotemporal Variation in Crude Protein of Shortgrass Steppe Forage" Remote Sensing 14, no. 4: 854. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14040854