
Mapping Benthic Algae and Cyanobacteria in River Channels from Aerial Photographs and Satellite Images: A Proof-of-Concept Investigation on the Buffalo National River, AR, USA



Abstract
:1. Introduction
- Collect field measurements of water depth and benthic algal cover and acquire various kinds of remotely sensed data from stream reaches along the Buffalo National River.
- Produce spectrally based bathymetric maps from aerial photographs and multispectral satellite images and evaluate the potential of spatially distributed depth information to facilitate mapping of benthic algae.
- Evaluate the feasibility of characterizing benthic algal blooms via remote sensing by applying a machine learning-based classification algorithm to different types of image data, including aerial orthophotos and multispectral satellite images, and assessing the accuracy of the resulting classifications via comparison to field observations of algal density.
2. Materials and Methods
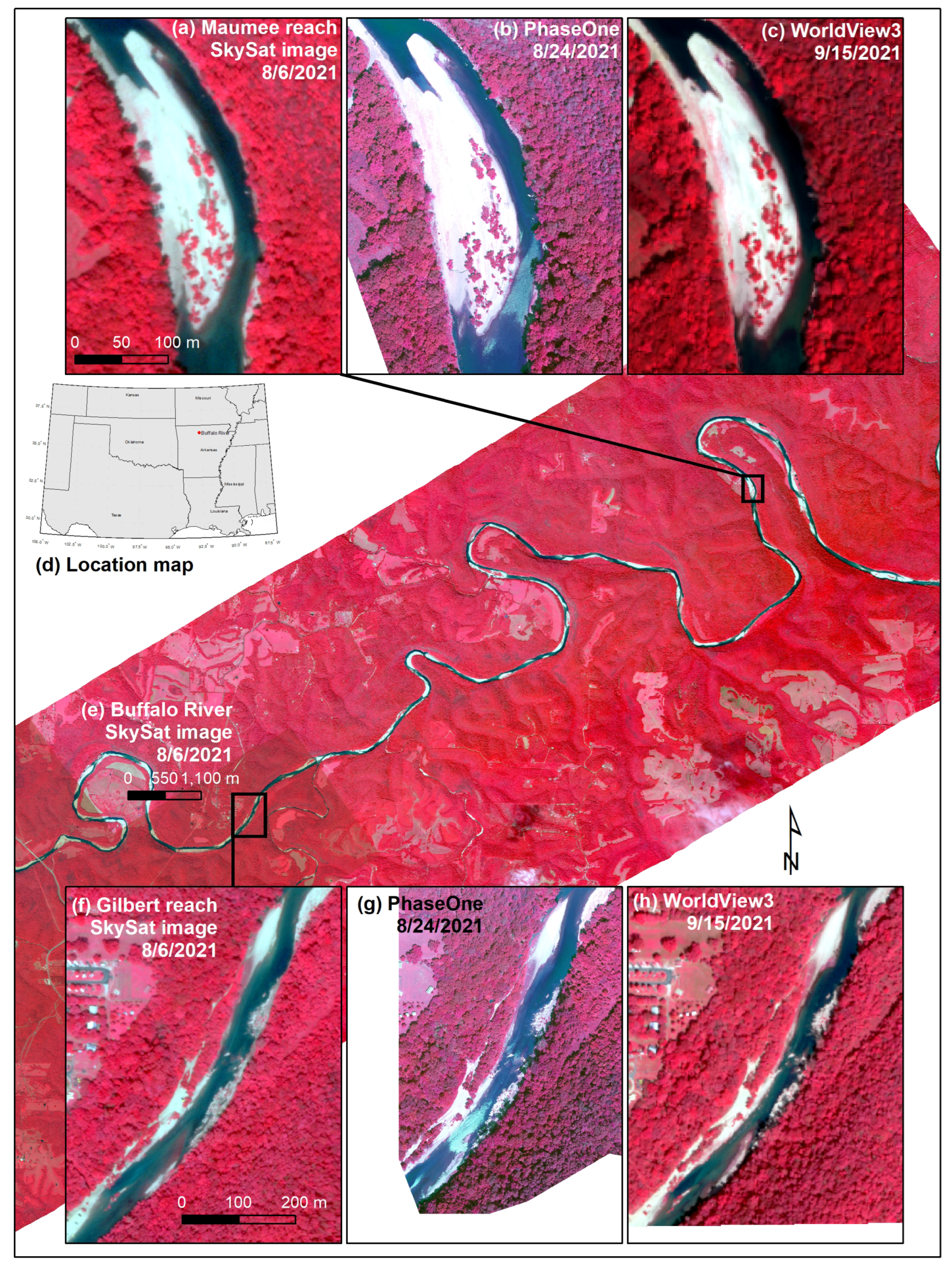
2.1. Study Area
2.2. Field Data Collection
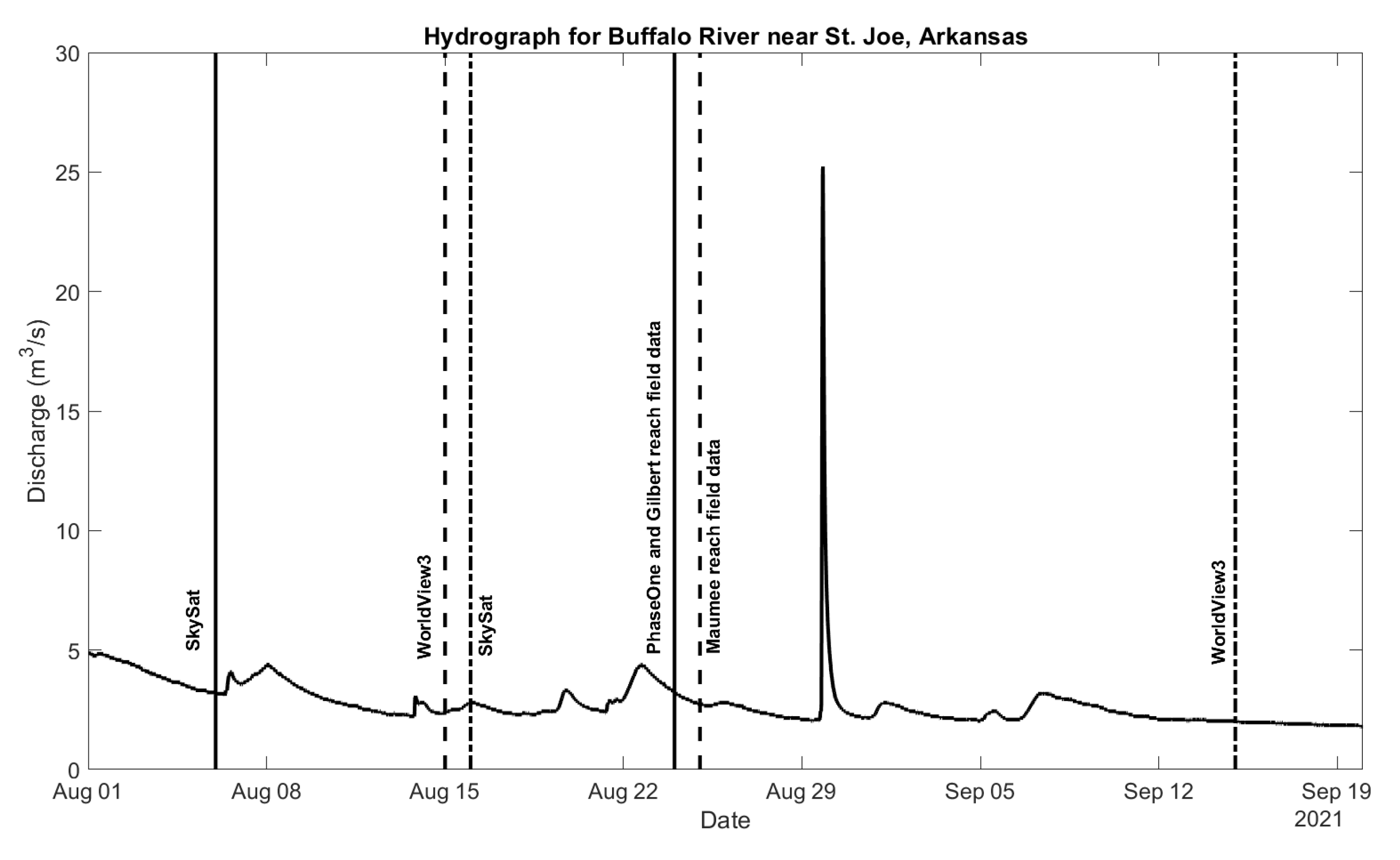
2.3. Remotely Sensed Data
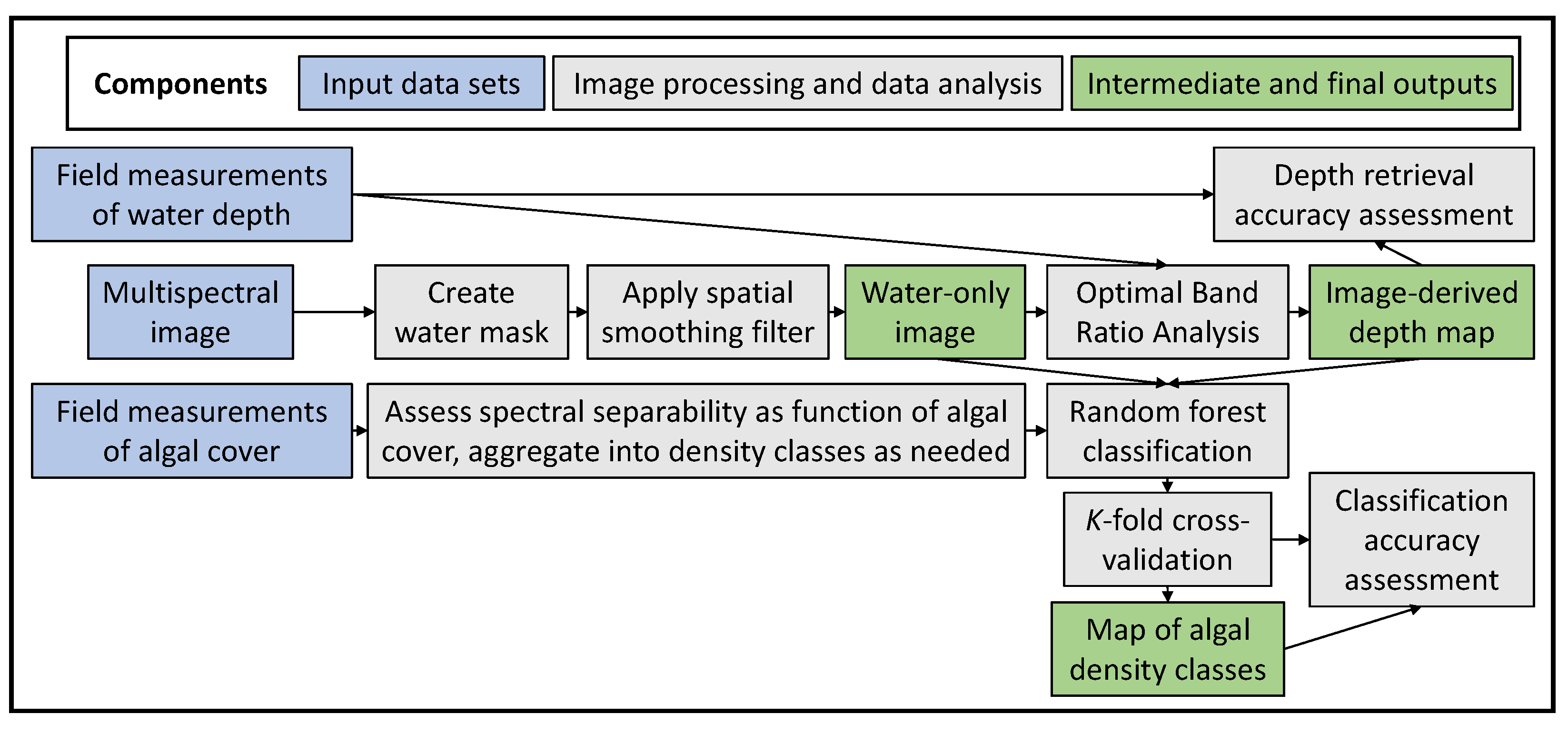
2.4. Workflow for Mapping Benthic Algal Blooms from Remotely Sensed Data
2.4.1. Spectrally Based Depth Retrieval
2.4.2. Machine Learning-Based Classification of Benthic Algal Cover
3. Results
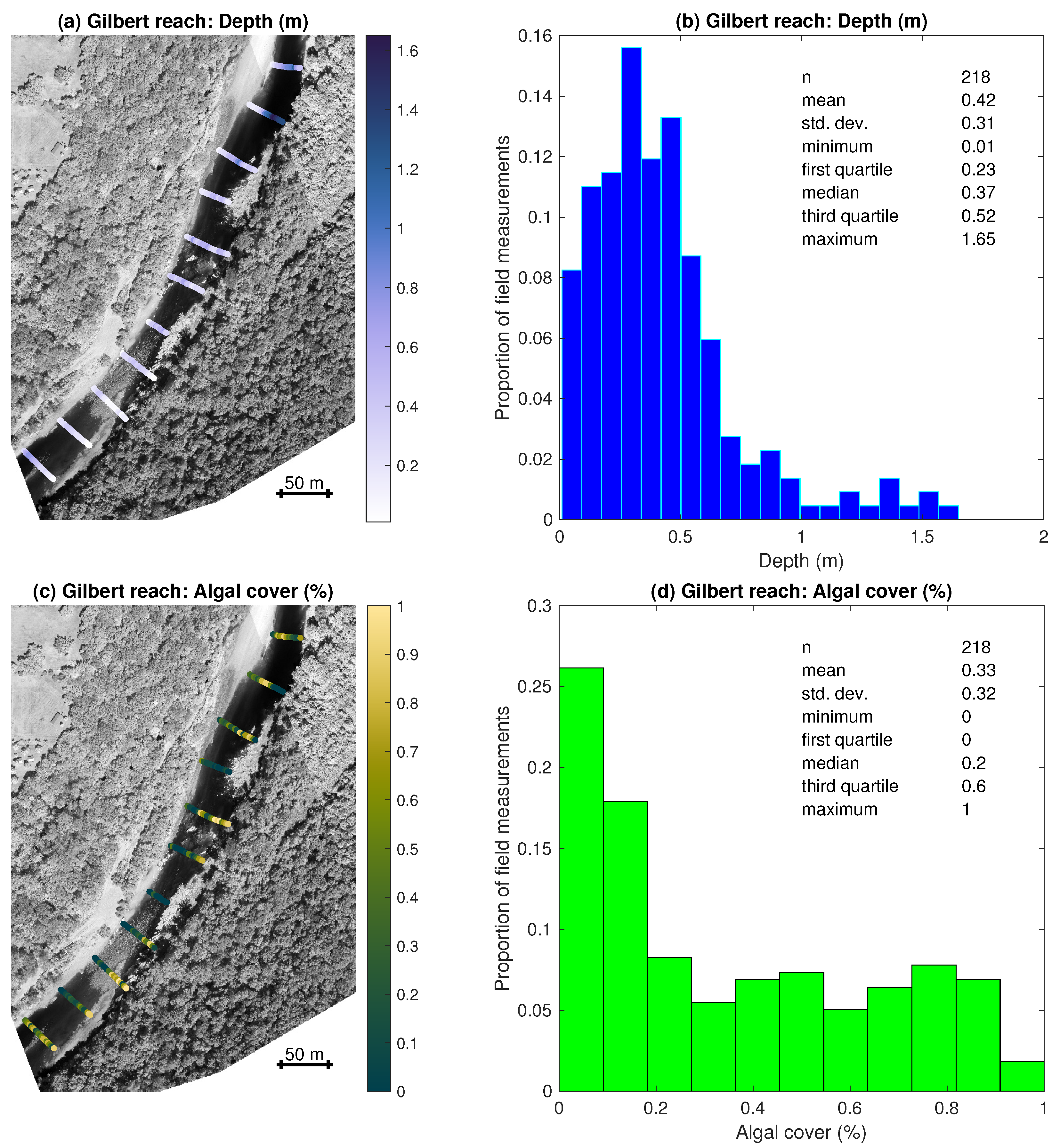
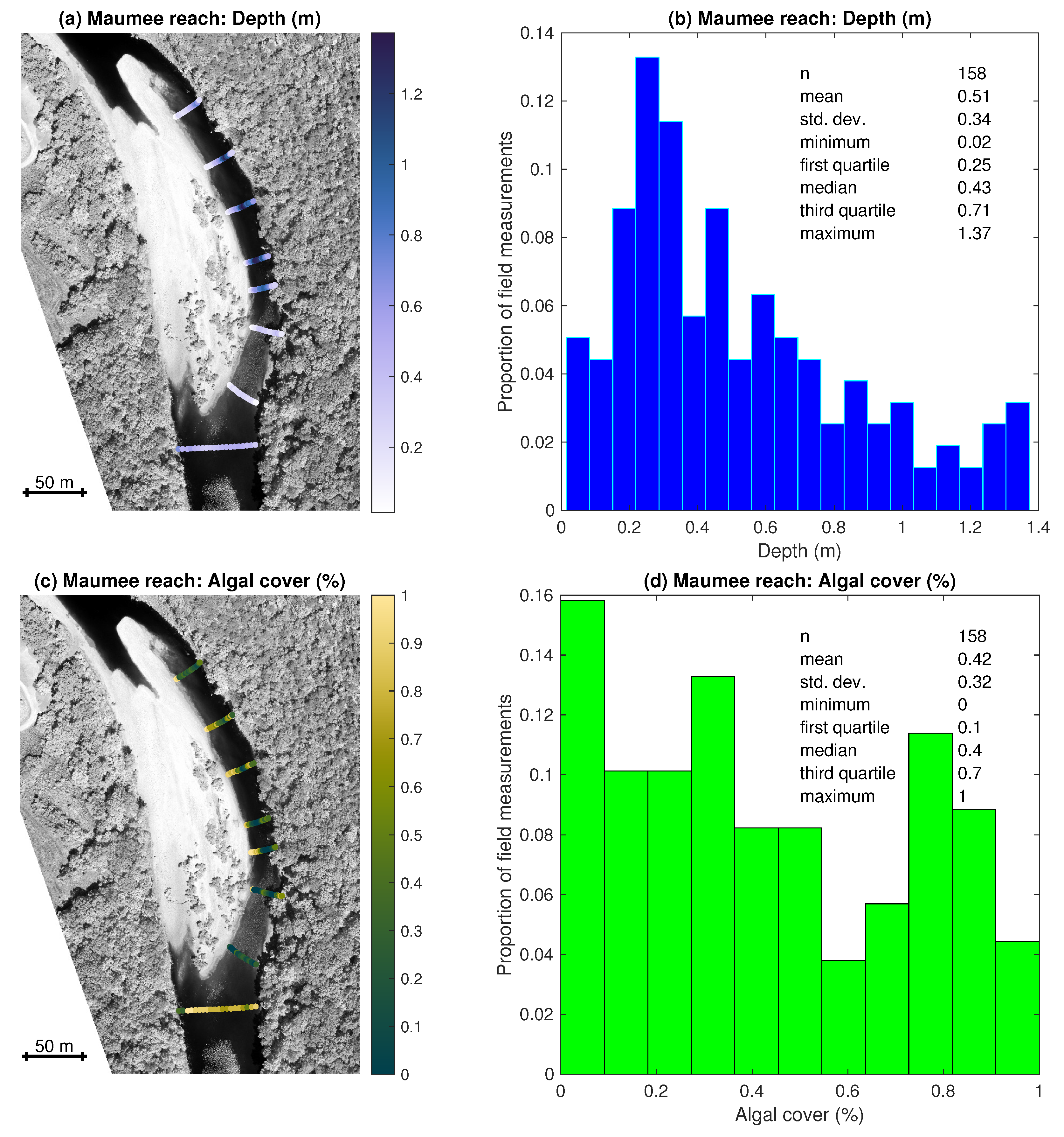
3.1. Field Observations of Depth and Algal Cover
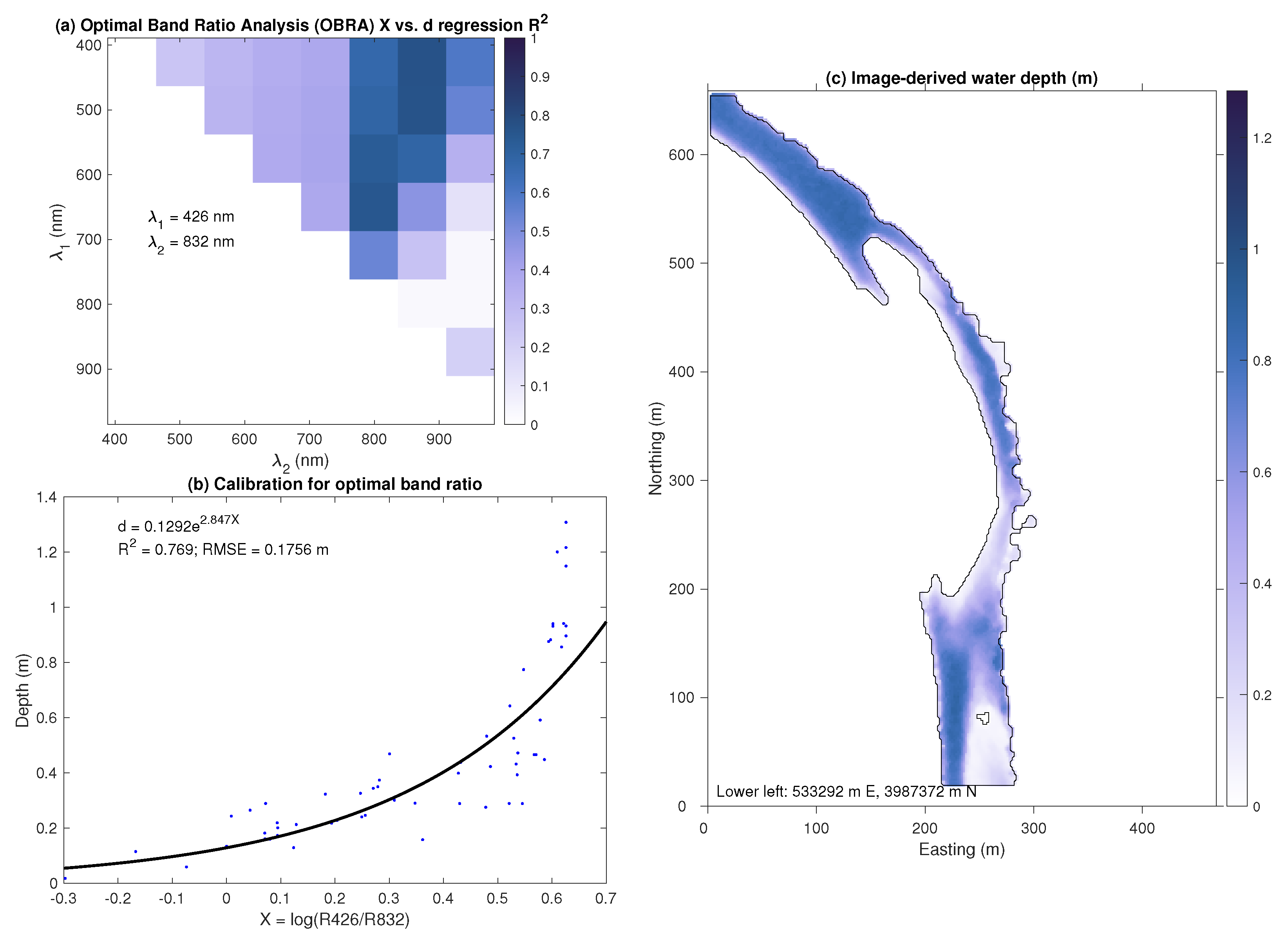
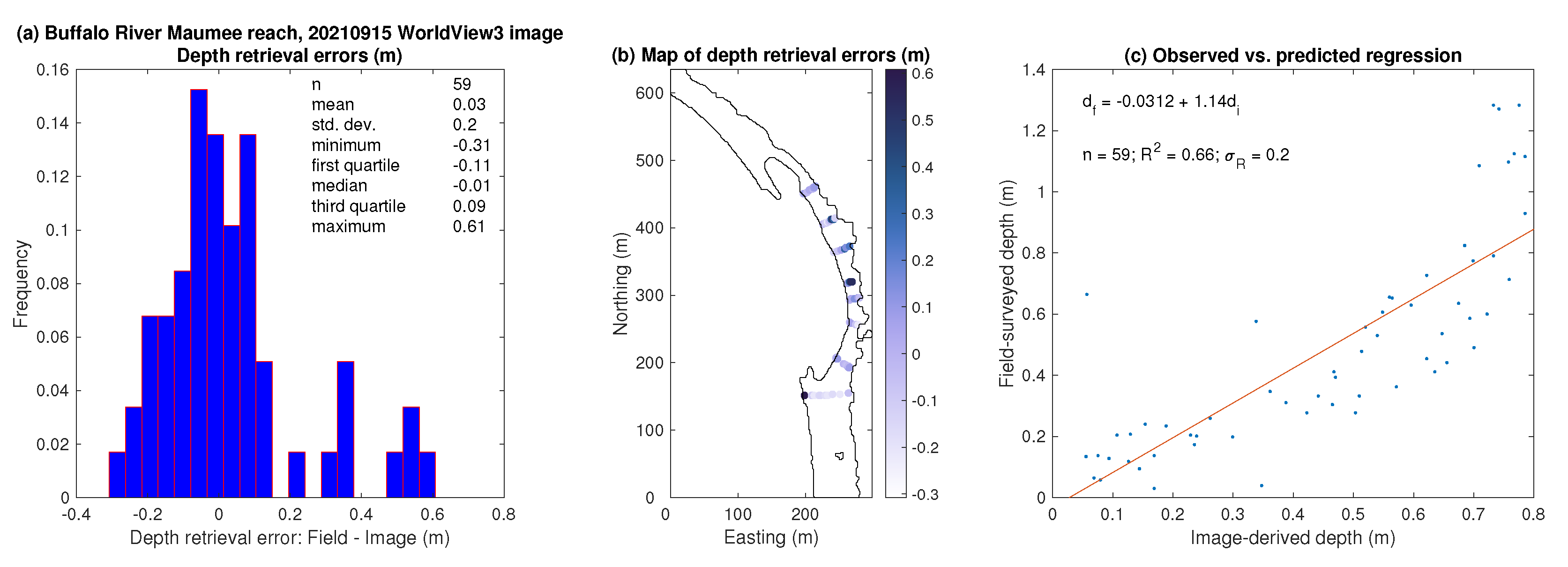
3.2. Bathymetric Mapping via OBRA
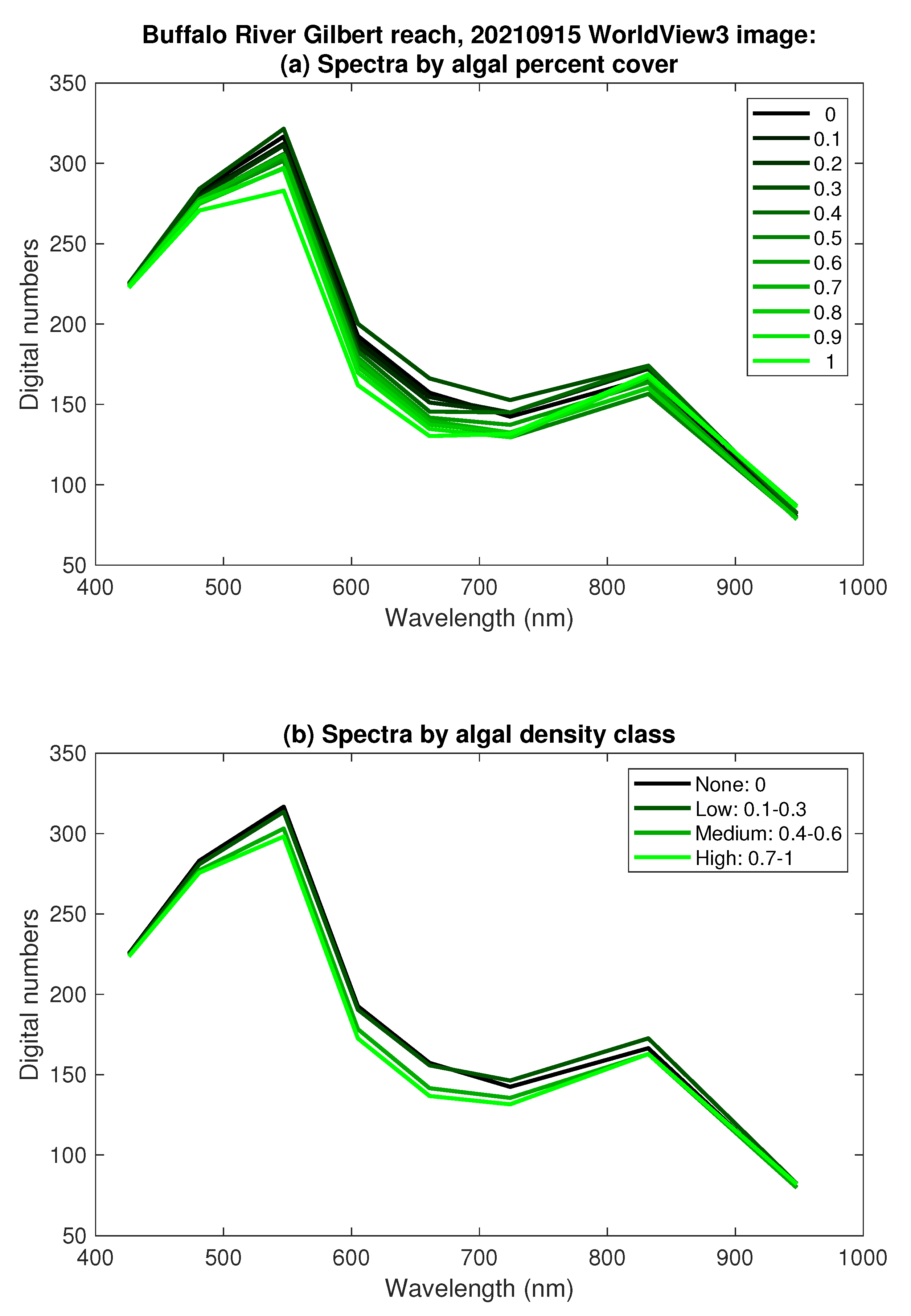
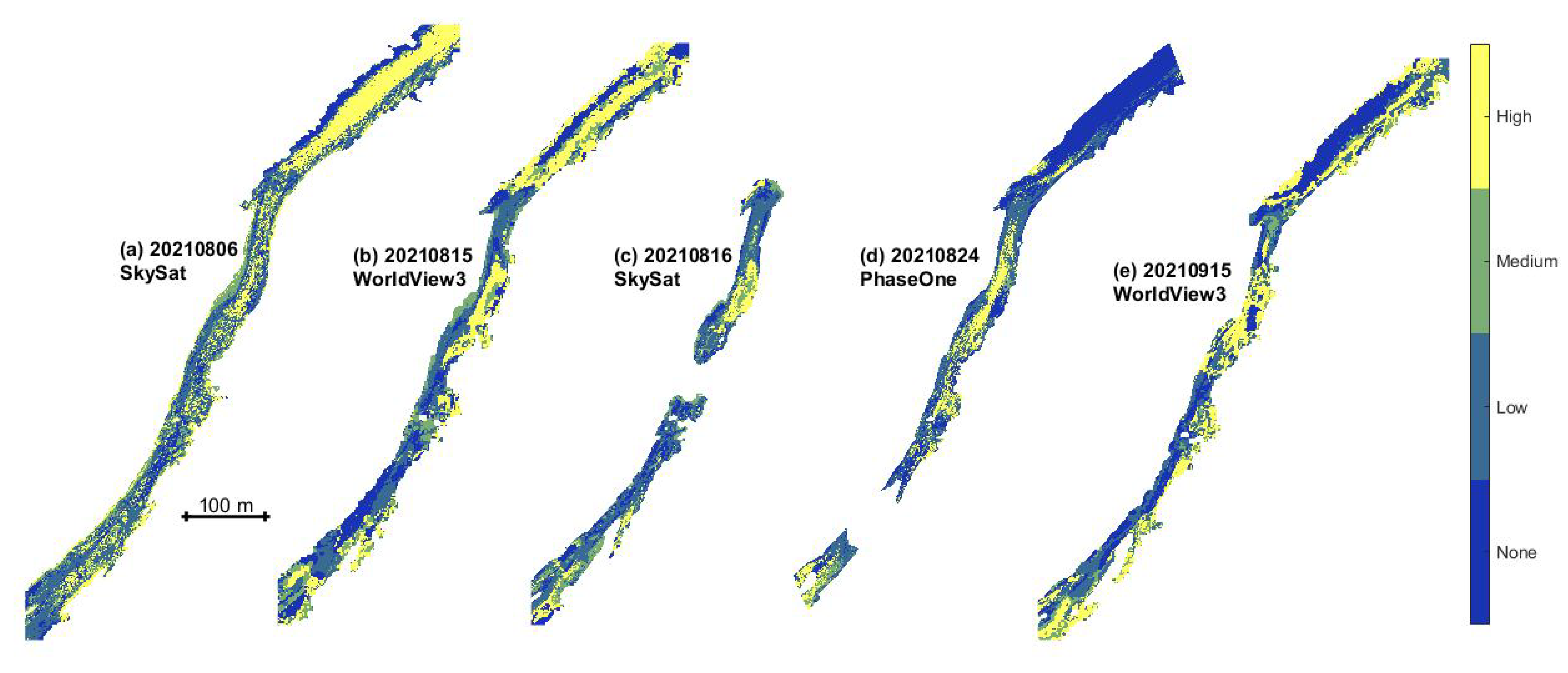
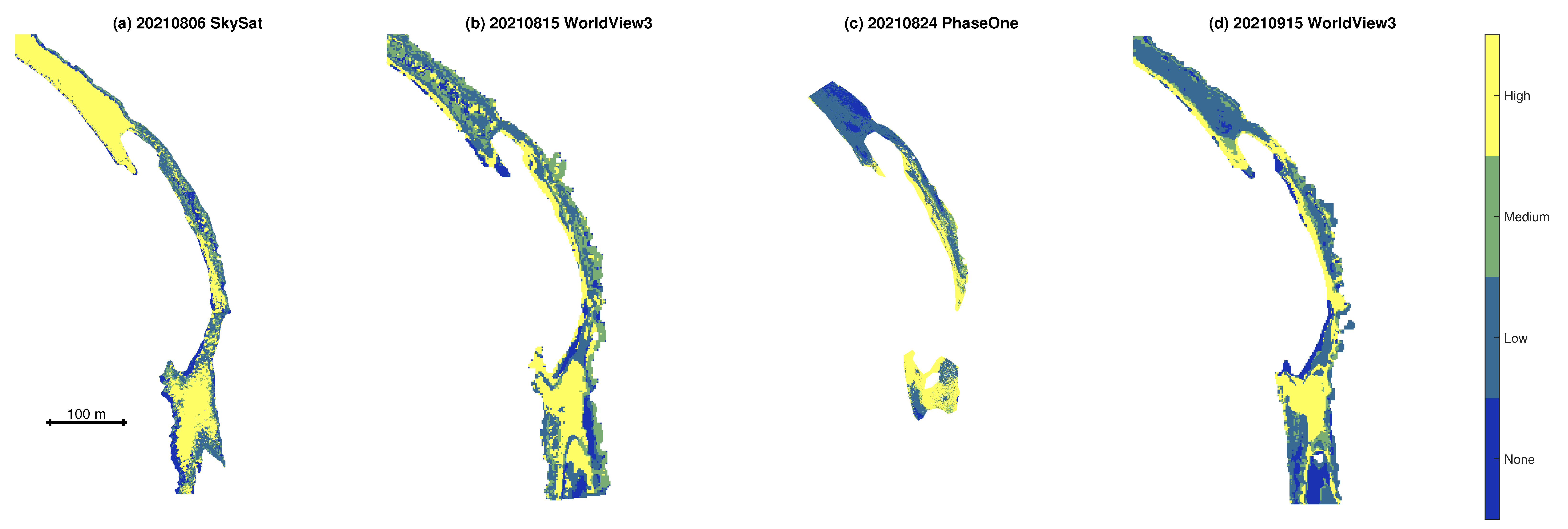
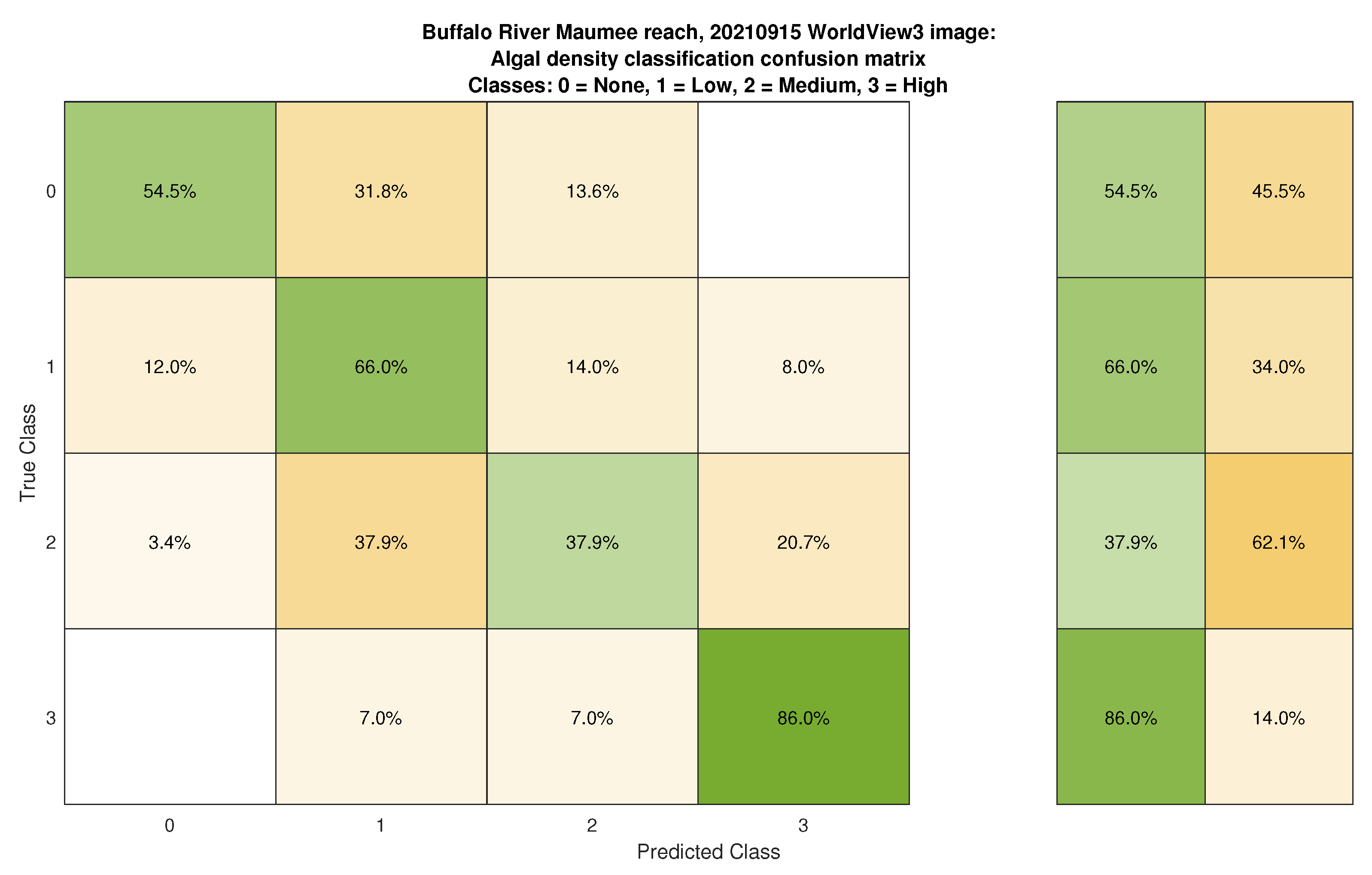
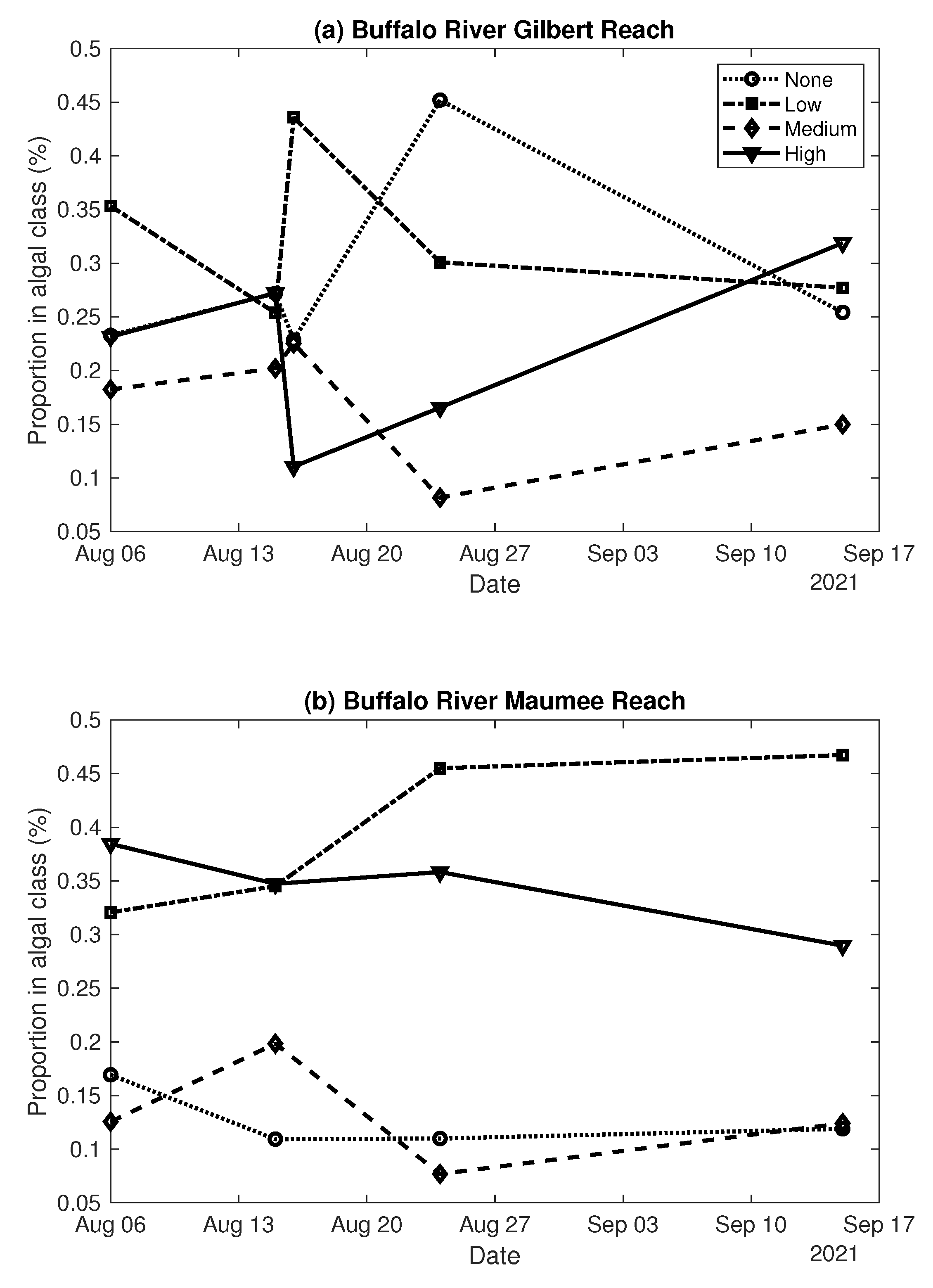
3.3. Classification and Mapping of Benthic Algae
4. Discussion
4.1. Monitoring Benthic Algae via Remote Sensing
4.2. Limitations and Lessons Learned
4.3. Advancing the Remote Sensing of Benthic Algae
5. Conclusions
- Field measurements of water depth and algal percent cover from two stream reaches along the Buffalo River captured a range of channel morphologies, with pools up to 1.65 m deep, and algal densities, with complete absence in some places and 100% coverage in others. Benthic algae tended to be more abundant in shallower areas.
- A spectrally based algorithm provided accurate depth estimates, with observed vs. predicted values up to 0.88, when applied to multispectral satellite images and orthophotos acquired from an aircraft. Image-derived depths were biased shallow, however, and only moderately precise, with typical mean biases on the order of 5% of the reach-averaged mean depth and root mean square error values from 30 to 42% of the reach-averaged mean depth.
- Spectra extracted from the images at the locations of field measurements were similar across a range of algal percent cover from 0 to 100%, with no clear, consistent distinctions between the 11 discrete levels. Aggregating the data into four algal density classes (none, low, medium, and high) only slightly improved spectral separability, especially for the imaging systems with four bands in the visible and near-infrared.
- Classified maps of algal density were produced by augmenting the original spectral bands with depth as an additional predictor variable and then training a bagged trees machine learning algorithm. Although the resulting classifications revealed some consistent spatial patterns and plausible trends over time, overall accuracies were modest, up to 64.6%.
- An important constraint on the reliability of the algal density classifications was the limited spectral resolution of the sensors employed in this study. Hyperspectral techniques capable of exploiting characteristic algal absorption features could enable a shift away from a classification-based framework toward a more quantitative approach focused on estimating algal biomass.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ward, J.V.; Tockner, K.; Arscott, D.B.; Claret, C. Riverine landscape diversity. Freshw. Biol. 2002, 47, 517–539. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and flow regulation of the world’s large river systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, J.C.; Michalak, A.M.; Pahlevan, N. Widespread global increase in intense lake phytoplankton blooms since the 1980s. Nature 2019, 574, 667–670. [Google Scholar] [CrossRef]
- Kislik, C.; Dronova, I.; Kelly, M. UAVs in Support of Algal Bloom Research: A Review of Current Applications and Future Opportunities. Drones 2018, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- US Environmental Protection Agency. Learn about Cyanobacteria and Cyanotoxins. Available online: https://www.epa.gov/cyanohabs/learn-about-cyanobacteria-and-cyanotoxins (accessed on 28 December 2021).
- Cyanotoxin—Zion National Park. Available online: https://www.nps.gov/zion/learn/news/cyanotoxin.htm (accessed on 2 February 2022).
- Priority Project: Harmful Algal Blooms. Available online: https://doimspp.sharepoint.com/sites/nps-coast/SitePages/Priority-Project--Harmful-Algal-Blooms.aspx#habs-products-proposals (accessed on 1 February 2022).
- Dodds, W.K.; Bouska, W.W.; Eitzmann, J.L.; Pilger, T.J.; Pitts, K.L.; Riley, A.J.; Schloesser, J.T.; Thornbrugh, D.J. Eutrophication of U.S. freshwaters: Analysis of potential economic damages. Environ. Sci. Technol. 2009, 43, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Burford, M.A.; Carey, C.C.; Hamilton, D.P.; Huisman, J.; Paerl, H.W.; Wood, S.A.; Wulff, A. Perspective: Advancing the research agenda for improving understanding of cyanobacteria in a future of global change. Harmful Algae 2020, 91, 101601. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.W.; Lazorchak, J.M.; Howard, M.D.; Johnson, M.V.V.; Morton, S.L.; Perkins, D.A.; Reavie, E.D.; Scott, G.I.; Smith, S.A.; Steevens, J.A. Are harmful algal blooms becoming the greatest inland water quality threat to public health and aquatic ecosystems? Environ. Toxicol. Chem. 2016, 35, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Dierssen, H.; Bracher, A.; Brando, V.; Loisel, H.; Ruddick, K. Data needs for hyperspectral detection of algal diversity across the globe. Oceanography 2020, 33, 74–79. [Google Scholar] [CrossRef]
- Cruz, R.C.; Reis Costa, P.; Vinga, S.; Krippahl, L.; Lopes, M.B. A Review of Recent Machine Learning Advances for Forecasting Harmful Algal Blooms and Shellfish Contamination. J. Mar. Sci. Eng. 2021, 9, 283. [Google Scholar] [CrossRef]
- Fetscher, A.E.; Howard, M.D.; Stancheva, R.; Kudela, R.M.; Stein, E.D.; Sutula, M.A.; Busse, L.B.; Sheath, R.G. Wadeable streams as widespread sources of benthic cyanotoxins in California, USA. Harmful Algae 2015, 49, 105–116. [Google Scholar] [CrossRef]
- Graham, J.L.; Dubrovsky, N.M.; Foster, G.M.; King, L.R.; Loftin, K.A.; Rosen, B.H.; Stelzer, E.A. Cyanotoxin occurrence in large rivers of the United States. Inland Waters 2020, 10, 109–117. [Google Scholar] [CrossRef]
- Wood, S.A.; Kelly, L.T.; Bouma-Gregson, K.; Humbert, J.F.; Laughinghouse, H.D.; Lazorchak, J.; McAllister, T.G.; McQueen, A.; Pokrzywinski, K.; Puddick, J.; et al. Toxic benthic freshwater cyanobacterial proliferations: Challenges and solutions for enhancing knowledge and improving monitoring and mitigation. Freshw. Biol. 2020, 65, 1824–1842. [Google Scholar] [CrossRef] [PubMed]
- Berkman, J.A.H.; Canova, M.G. Chapter A7. Section 7.4. Algal Biomass Indicators; Technical Report; U.S. Geological Survey: Reston, VA, USA, 2007. [CrossRef]
- Coffer, M.M.; Schaeffer, B.A.; Salls, W.B.; Urquhart, E.; Loftin, K.A.; Stumpf, R.P.; Werdell, P.J.; Darling, J.A. Satellite remote sensing to assess cyanobacterial bloom frequency across the United States at multiple spatial scales. Ecol. Indic. 2021, 128, 107822. [Google Scholar] [CrossRef]
- Khan, R.M.; Salehi, B.; Mahdianpari, M.; Mohammadimanesh, F.; Mountrakis, G.; Quackenbush, L.J. A Meta-Analysis on Harmful Algal Bloom (HAB) Detection and Monitoring: A Remote Sensing Perspective. Remote Sens. 2021, 13, 4347. [Google Scholar] [CrossRef]
- Visser, F.; Wallis, C.; Sinnott, A.M. Optical remote sensing of submerged aquatic vegetation: Opportunities for shallow clearwater streams. Limnologica 2013, 43, 388–398. [Google Scholar] [CrossRef]
- Visser, F.; Buis, K.; Verschoren, V.; Meire, P. Depth Estimation of Submerged Aquatic Vegetation in Clear Water Streams Using Low-Altitude Optical Remote Sensing. Sensors 2015, 15, 25287. [Google Scholar] [CrossRef] [Green Version]
- Legleiter, C.J.; Stegman, T.K.; Overstreet, B.T. Spectrally based mapping of riverbed composition. Geomorphology 2016, 264, 61–79. [Google Scholar] [CrossRef] [Green Version]
- Maritorena, S.; Morel, A.; Gentili, B. Diffuse-reflectance of oceanic shallow waters—Influence of water depth and bottom albedo. Limnol. Oceanogr. 1994, 39, 1689–1703. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Zimmerman, R.C.; Leathers, R.A.; Downes, T.V.; Davis, C.O. Ocean color remote sensing of seagrass and bathymetry in the Bahamas Banks by high-resolution airborne imagery. Limnol. Oceanogr. 2003, 48, 444–455. [Google Scholar] [CrossRef]
- Mobley, C.D.; Sundman, L.K.; Davis, C.O.; Bowles, J.H.; Downes, T.V.; Leathers, R.A.; Montes, M.J.; Bissett, W.P.; Kohler, D.D.R.; Reid, R.P.; et al. Interpretation of hyperspectral remote-sensing imagery by spectrum matching and look-up tables. Appl. Opt. 2005, 44, 3576–3592. [Google Scholar] [CrossRef]
- Slonecker, T.; Kalaly, S.; Young, J.; Furedi, M.A.; Maloney, K.; Hamilton, D.; Richard, E.; Zinecker, E. A Preliminary Assessment of Hyperspectral Remote Sensing Technology for Mapping Submerged Aquatic Vegetation in the Upper Delaware River National Parks (USA). Adv. Remote Sens. 2018, 7, 290–312. [Google Scholar] [CrossRef] [Green Version]
- Kislik, C.; Genzoli, L.; Lyons, A.; Kelly, M. Application of UAV Imagery to Detect and Quantify Submerged Filamentous Algae and Rooted Macrophytes in a Non-Wadeable River. Remote Sens. 2020, 12, 3332. [Google Scholar] [CrossRef]
- Legleiter, C.; Hodges, S. Remotely Sensed Data and Field Measurements of Water Depth and Percent Cover of Benthic Algae from Two Reaches of the Buffalo National River in Arkansas Acquired in August and September 2021; U.S. Geological Survey Data Release: Reston, VA, USA, 2022. [CrossRef]
- Algae and Nurient Sourcing. Available online: https://www.nps.gov/articles/000/algae-and-nutrient-sourcing.htm (accessed on 7 February 2022).
- Algae Bloom Complaint Form|DEQ. Available online: https://www.adeq.state.ar.us/complaints/forms/algae_complaint.aspx (accessed on 7 February 2022).
- Fishing—Buffalo National River. Available online: https://www.nps.gov/buff/planyourvisit/fishing.htm (accessed on 7 February 2022).
- Office of the Federal Register, National Archives and Records Administration. 80 FR 24961—Endangered Species; Marine Mammals; Receipt of Applications for Permit; Office of the Federal Register, National Archives and Records Administration: Washington, DC, USA, 2015.
- Berkman, D.N.; Lauraas, J. Water Resources Management Plan: Buffalo National River, Arkansas; Technical Report; U.S. National Park Service: Harrison, AR, USA, 2004.
- U.S. Geological Survey. USGS Water Data for the Nation: U.S. Geological Survey National Water Information System Database. USGS 07056000 Buffalo River near St. Joe, AR. Available online: https://waterdata.usgs.gov/ar/nwis/inventory/?site_no=07056000 (accessed on 22 December 2021). [CrossRef]
- Leica Zeno GG04 Plus Smart Antenna for High Accuracy Everywhere. Available online: https://leica-geosystems.com/en-us/products/gis-collectors/smart-antennas/leica-zeno-gg04-plus (accessed on 31 January 2022).
- High-Resolution Imagery with Planet Satellite Tasking. Available online: https://www.planet.com/products/hi-res-monitoring/ (accessed on 7 February 2022).
- Maxar—Archive Search & Discovery. Available online: https://discover.digitalglobe.com/ (accessed on 7 February 2022).
- Vanhellemont, Q.; Ruddick, K. Atmospheric correction of metre-scale optical satellite data for inland and coastal water applications. Remote Sens. Environ. 2018, 216, 586–597. [Google Scholar] [CrossRef]
- Pahlevan, N.; Mangin, A.; Balasubramanian, S.V.; Smith, B.; Alikas, K.; Arai, K.; Barbosa, C.; Bélanger, S.; Binding, C.; Bresciani, M.; et al. ACIX-Aqua: A global assessment of atmospheric correction methods for Landsat-8 and Sentinel-2 over lakes, rivers, and coastal waters. Remote Sens. Environ. 2021, 258, 112366. [Google Scholar] [CrossRef]
- Lyzenga, D.R. Passive Remote-Sensing Techniques for Mapping Water Depth and Bottom Features. Appl. Opt. 1978, 17, 379–383. [Google Scholar] [CrossRef]
- Niroumand-Jadidi, M.; Vitti, A.; Lyzenga, D.R. Multiple Optimal Depth Predictors Analysis (MODPA) for river bathymetry: Findings from spectroradiometry, simulations, and satellite imagery. Remote Sens. Environ. 2018, 218, 132–147. [Google Scholar] [CrossRef]
- Tomsett, C.; Leyland, J. Remote sensing of river corridors: A review of current trends and future directions. River Res. Appl. 2019, 35, 779–803. [Google Scholar] [CrossRef]
- Legleiter, C.; Roberts, D.A.; Lawrence, R.L. Spectrally based remote sensing of river bathymetry. Earth Surf. Process. Landforms 2009, 34, 1039–1059. [Google Scholar] [CrossRef]
- Legleiter, C.; Harrison, L.R. Remote Sensing of River Bathymetry: Evaluating a Range of Sensors, Platforms, and Algorithms on the Upper Sacramento River, California, USA. Water Resour. Res. 2019, 55, 2142–2169. [Google Scholar] [CrossRef]
- Legleiter, C.J. The optical river bathymetry toolkit. River Res. Appl. 2021, 37, 555–568. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Stackelberg, P.E.; Belitz, K.; Brown, C.J.; Erickson, M.L.; Elliott, S.M.; Kauffman, L.J.; Ransom, K.M.; Reddy, J.E. Machine Learning Predictions of pH in the Glacial Aquifer System, Northern USA. Groundwater 2021, 59, 352–368. [Google Scholar] [CrossRef] [PubMed]
- Congalton, R.G.; Green, K. Assessing the Acuracy of Remotely Sensed Data: Principles and Practices; CRC Press: Boca Raton, FL, USA, 1999; p. 137. [Google Scholar]
- Glibert, P.M. Eutrophication, harmful algae and biodiversity—Challenging paradigms in a world of complex nutrient changes. Mar. Pollut. Bull. 2017, 124, 591–606. [Google Scholar] [CrossRef] [PubMed]
- Overstreet, B.T.; Legleiter, C. Removing sun glint from optical remote sensing images of shallow rivers. Earth Surf. Process. Landforms 2017, 42, 318–333. [Google Scholar] [CrossRef]
- Legleiter, C.; Mobley, C.D.; Overstreet, B.T. A framework for modeling connections between hydraulics, water surface roughness, and surface reflectance in open channel flows. J. Geophys. Res. Earth Surf. 2017, 122, 1715–1741. [Google Scholar] [CrossRef]
- Aleksandra, K.; Fantina, M.; Marco, S.; Ferrarin, C.; Giacomo, M.G. Assessment of submerged aquatic vegetation abundance using multibeam sonar in very shallow and dynamic environment. The Lagoon of Venice (Italy) case study. In Proceedings of the 2015 IEEE/OES Acoustics in Underwater Geosciences Symposium (RIO Acoustics), Rio de Janeiro, Brazil, 29–31 July 2015; pp. 1–7. [Google Scholar] [CrossRef]
- Held, P.; von Deimling, J.S. New feature classes for acoustic habitat mapping—A multibeam echosounder point cloud analysis for mapping submerged aquatic vegetation (SAV). Geosciences 2019, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Legleiter, C.; Fosness, R. Defining the Limits of Spectrally Based Bathymetric Mapping on a Large River. Remote Sens. 2019, 11, 665. [Google Scholar] [CrossRef] [Green Version]
- Rousso, B.Z.; Bertone, E.; Stewart, R.; Hamilton, D.P. A systematic literature review of forecasting and predictive models for cyanobacteria blooms in freshwater lakes. Water Res. 2020, 182, 115959. [Google Scholar] [CrossRef]
- Iiames, J.S.; Salls, W.B.; Mehaffey, M.H.; Nash, M.S.; Christensen, J.R.; Schaeffer, B.A. Modeling Anthropogenic and Environmental Influences on Freshwater Harmful Algal Bloom Development Detected by MERIS Over the Central United States. Water Resour. Res. 2021, 57, e2020WR028946. [Google Scholar] [CrossRef]
- Paine, E.C.; Slonecker, E.T.; Simon, N.S.; Rosen, B.H.; Resmini, R.G.; Allen, D.W. Optical characterization of two cyanobacteria genera, Aphanizomenon and Microcystis, with hyperspectral microscopy. J. Appl. Remote Sens. 2018, 12, 036013. [Google Scholar] [CrossRef]
- Slonecker, T.; Bufford, B.; Graham, J.; Carpenter, K.; Opstal, D.; Simon, N.; Hall, N. Hyperspectral Reflectance Characteristics of Cyanobacteria. Adv. Remote Sens. 2021, 10, 66–77. [Google Scholar] [CrossRef]
- Slonecker, T.; Simon, N.; Graham, J.; Allen, D.; Bufford, B.; Evans, M.; Carpenter, K.; Griffin, D.; Hall, N.; Jones, D.; et al. Hyperspectral Characterization of Common Cyanobacteria Associated with Harmful Algal Blooms (Ver. 2.0, October 2020); U.S. Geological Survey Data Release: Reston, VA, USA, 2020. [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensor | Operator | Platform | Pixel Size (m) | Spectral Bands and Center Wavelengths (nm) |
|---|---|---|---|---|
| SkySat | Planet Labs | Satellite | 0.5 | 4: B (482.5), G (555), R (650), NIR 1 (820) |
| WorldView3 | DigitalGlobe | Satellite | 1.81, 2 2 | 8: Coastal blue (426), B (481), G (547), Yellow (605), R (661), Red edge (724), NIR1 (832), NIR2 (948) |
| PhaseOne | USFWS 3 | Aircraft | 0.088 | 4: B (450), G (550), R (650), NIR (750) |
| Date | Sensor | Reach | OBRA | Mean Error (m) | Normalized Mean Error | RMSE (m) | Normalized RMSE | Observed vs. Predicted |
|---|---|---|---|---|---|---|---|---|
| 6 August 2021 | SkySat | Gilbert | 0.50 | 0.03 | 0.065 | 0.22 | 0.478 | 0.51 |
| 15 August 2021 | WorldView3 | Gilbert | 0.72 | 0.01 | 0.022 | 0.22 | 0.478 | 0.45 |
| 16 August 2021 | SkySat | Gilbert | 0.42 | 0.04 | 0.087 | 0.21 | 0.457 | 0.49 |
| 24 August 2021 | PhaseOne | Gilbert | 0.81 | 0 | 0.000 | 0.14 | 0.304 | 0.88 |
| 15 September 2021 | WorldView3 | Gilbert | 0.72 | 0.03 | 0.065 | 0.2 | 0.435 | 0.77 |
| 6 August 2021 | SkySat | Maumee | 0.67 | 0.09 | 0.161 | 0.24 | 0.429 | 0.58 |
| 15 August 2021 | WorldView3 | Maumee | 0.85 | −0.03 | −0.054 | 0.17 | 0.304 | 0.67 |
| 24 August 2021 | PhaseOne | Maumee | 0.90 | 0.01 | 0.018 | 0.19 | 0.339 | 0.73 |
| 15 September 2021 | WorldView3 | Maumee | 0.77 | 0.03 | 0.054 | 0.2 | 0.357 | 0.66 |
| Algal Density Class | Producer’s Accuracy | User’s Accuracy |
|---|---|---|
| None | 54.5 | 63.1 |
| Low | 66 | 61.1 |
| Medium | 37.9 | 45.8 |
| High | 86.5 | 78.7 |
| Reach | Date | Sensor | Overall Accuracy without Depth (%) | Overall Accuracy with Depth (%) |
|---|---|---|---|---|
| Gilbert | 6 August 2021 | SkySat | 38.5 | 37.6 |
| Gilbert | 15 August 2021 | WorldView3 | 47.1 | 54.4 |
| Gilbert | 16 August 2021 | SkySat | 39 | 38.3 |
| Gilbert | 24 August 2021 | PhaseOne | 47.8 | 49 |
| Gilbert | 15 September 2021 | WorldView3 | 50.2 | 54.3 |
| Maumee | 6 August 2021 | SkySat | 49.2 | 55.4 |
| Maumee | 15 August 2021 | WorldView3 | 59.3 | 62 |
| Maumee | 24 August 2021 | PhaseOne | 50 | 63.7 |
| Maumee | 15 September 2021 | WorldView3 | 61.1 | 64.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legleiter, C.J.; Hodges, S.W. Mapping Benthic Algae and Cyanobacteria in River Channels from Aerial Photographs and Satellite Images: A Proof-of-Concept Investigation on the Buffalo National River, AR, USA. Remote Sens. 2022, 14, 953. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14040953
Legleiter CJ, Hodges SW. Mapping Benthic Algae and Cyanobacteria in River Channels from Aerial Photographs and Satellite Images: A Proof-of-Concept Investigation on the Buffalo National River, AR, USA. Remote Sensing. 2022; 14(4):953. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14040953
Chicago/Turabian StyleLegleiter, Carl J., and Shawn W. Hodges. 2022. "Mapping Benthic Algae and Cyanobacteria in River Channels from Aerial Photographs and Satellite Images: A Proof-of-Concept Investigation on the Buffalo National River, AR, USA" Remote Sensing 14, no. 4: 953. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14040953