
Spatial-Temporal Dynamics of China’s Terrestrial Biodiversity: A Dynamic Habitat Index Diagnostic

 ,
,
Abstract
:
1. Introduction
2. Data and Methodology
2.1. Data
2.2. Dynamic Habitat Index
2.3. Regression Tendency Analysis
2.4. Decadal Mean and Inter-Annual Variability
3. Application of the Dynamic Habitat Analysis: Results for China
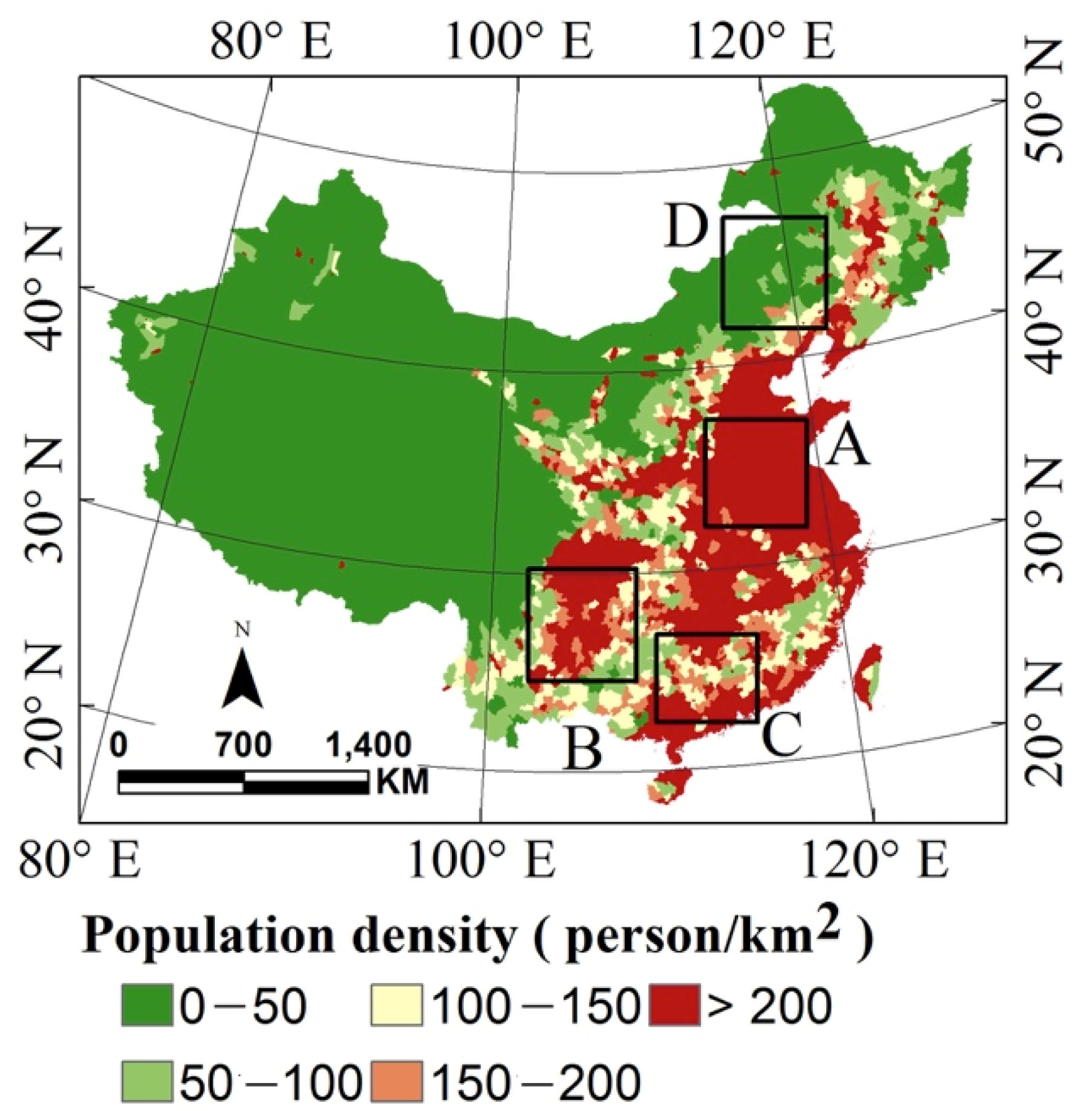
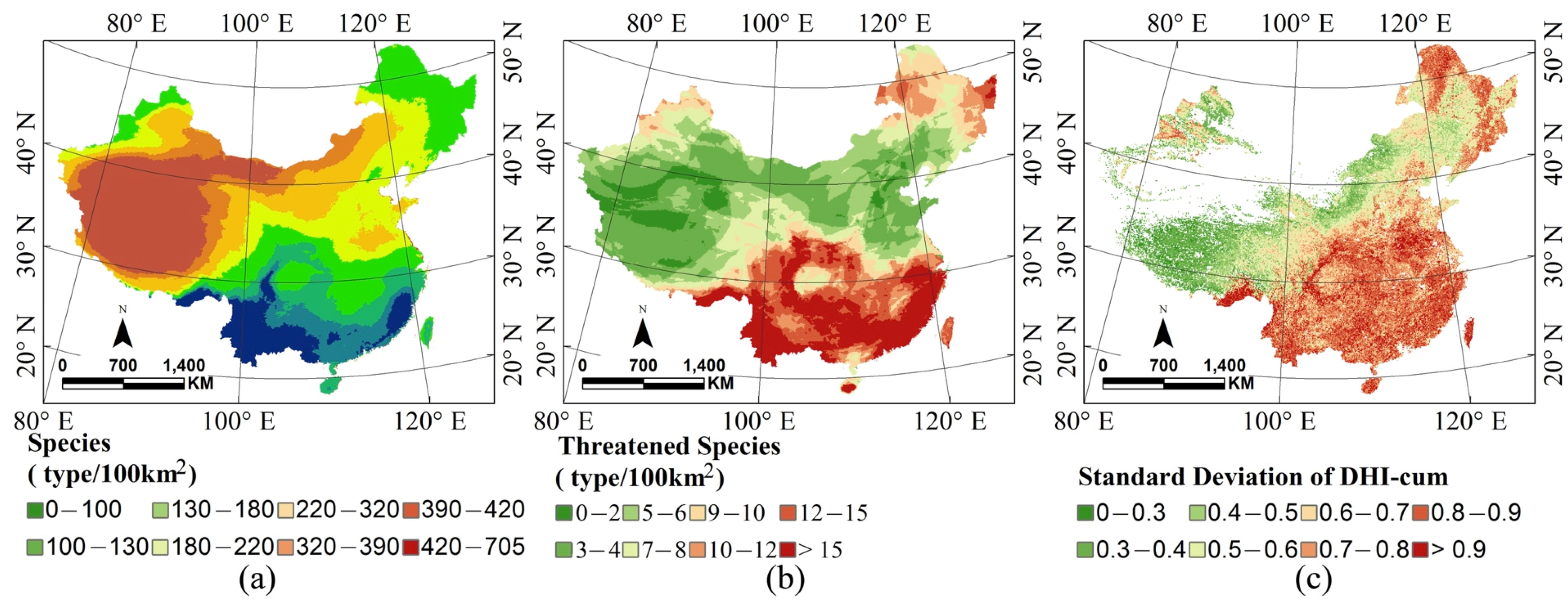
3.1. Ecological Setting
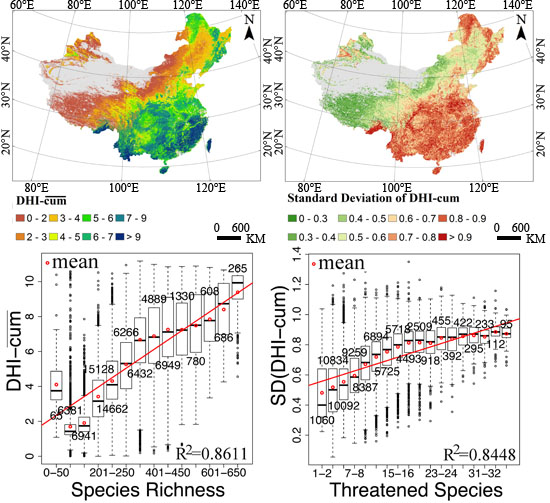
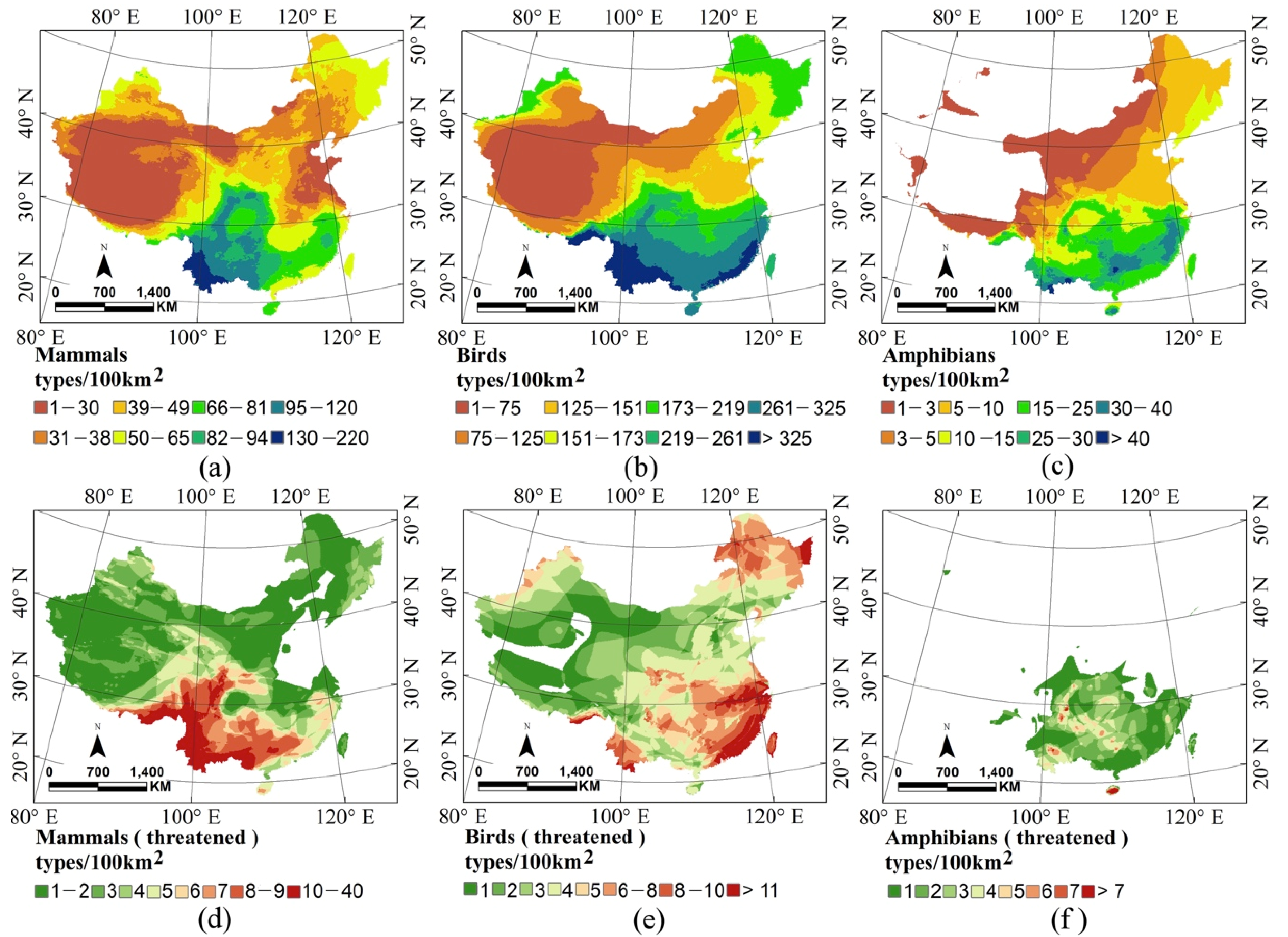
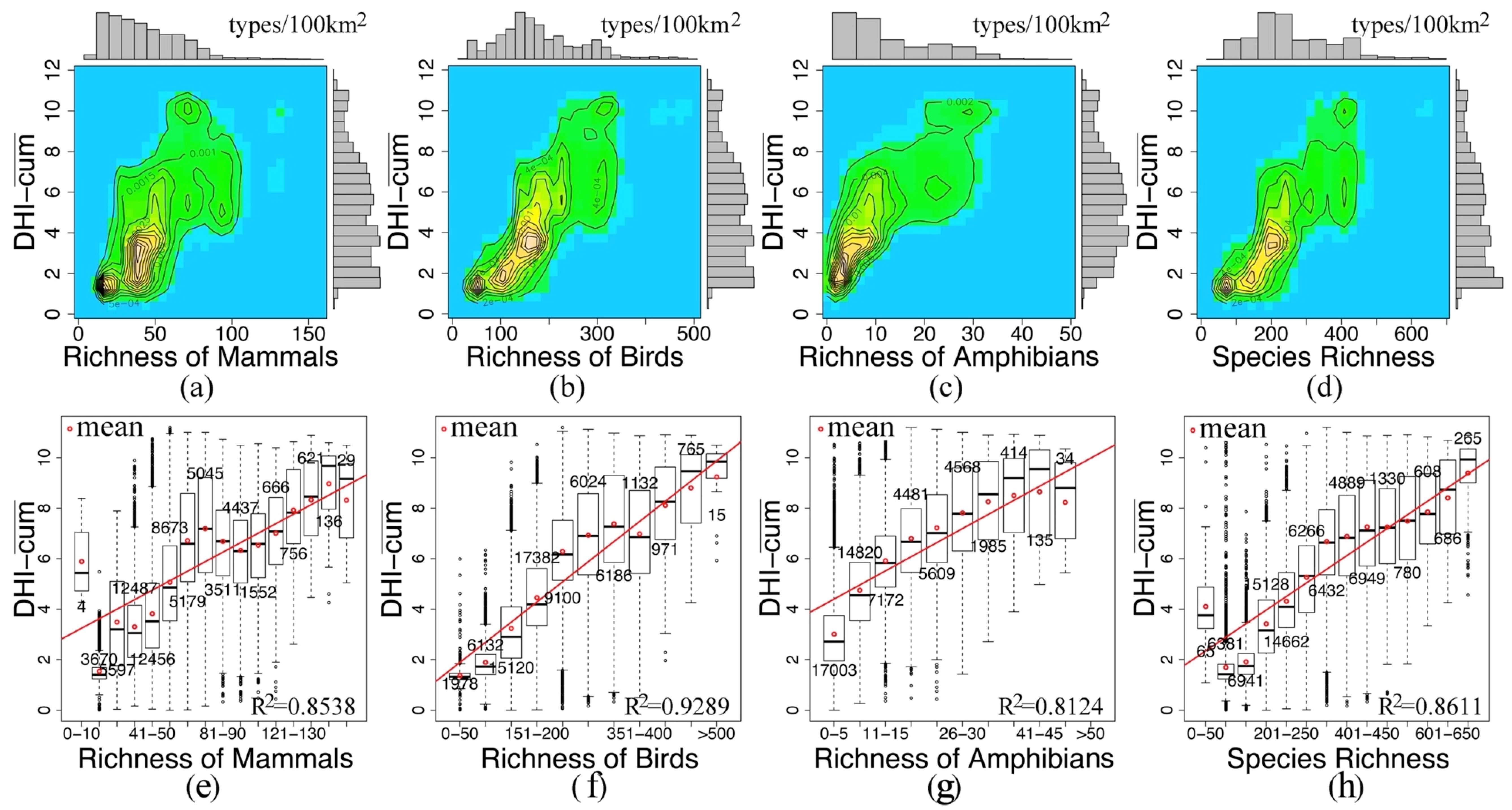
3.2. Remote Sensing Surrogates for Species and Threatened Species Richness
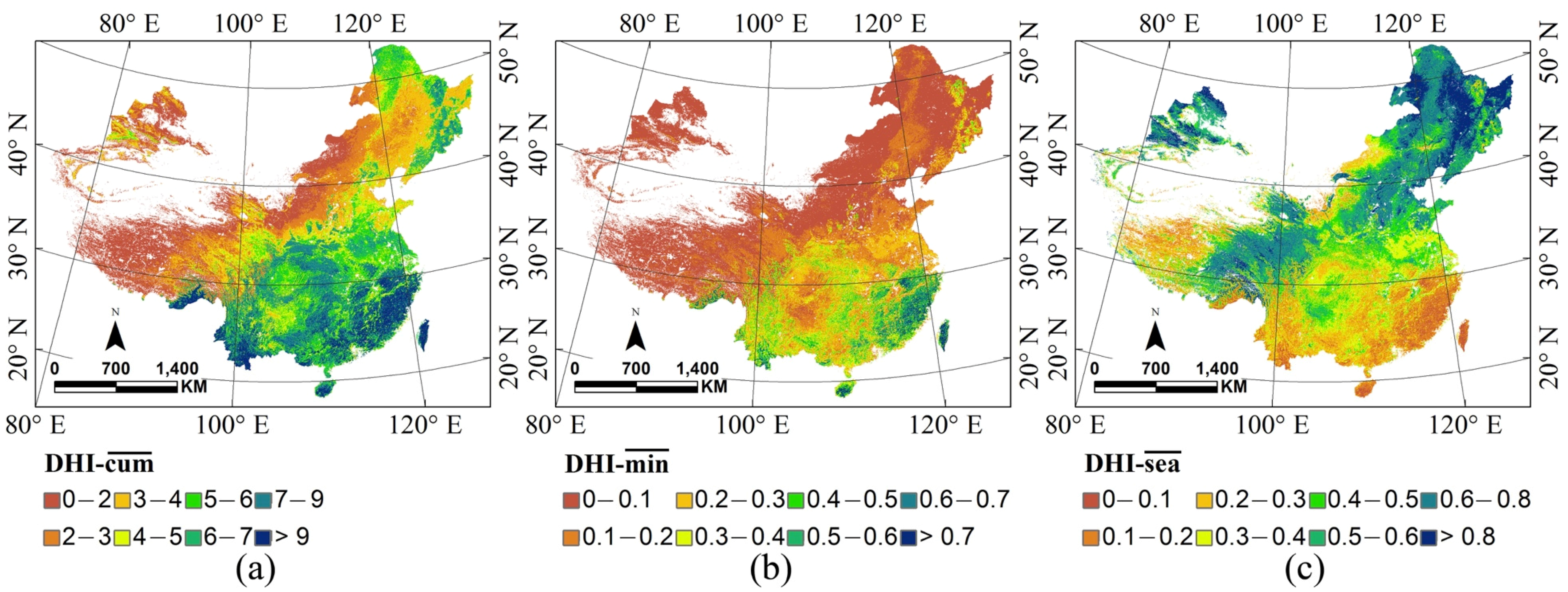
3.3. Dynamic Habitat Analysis in China
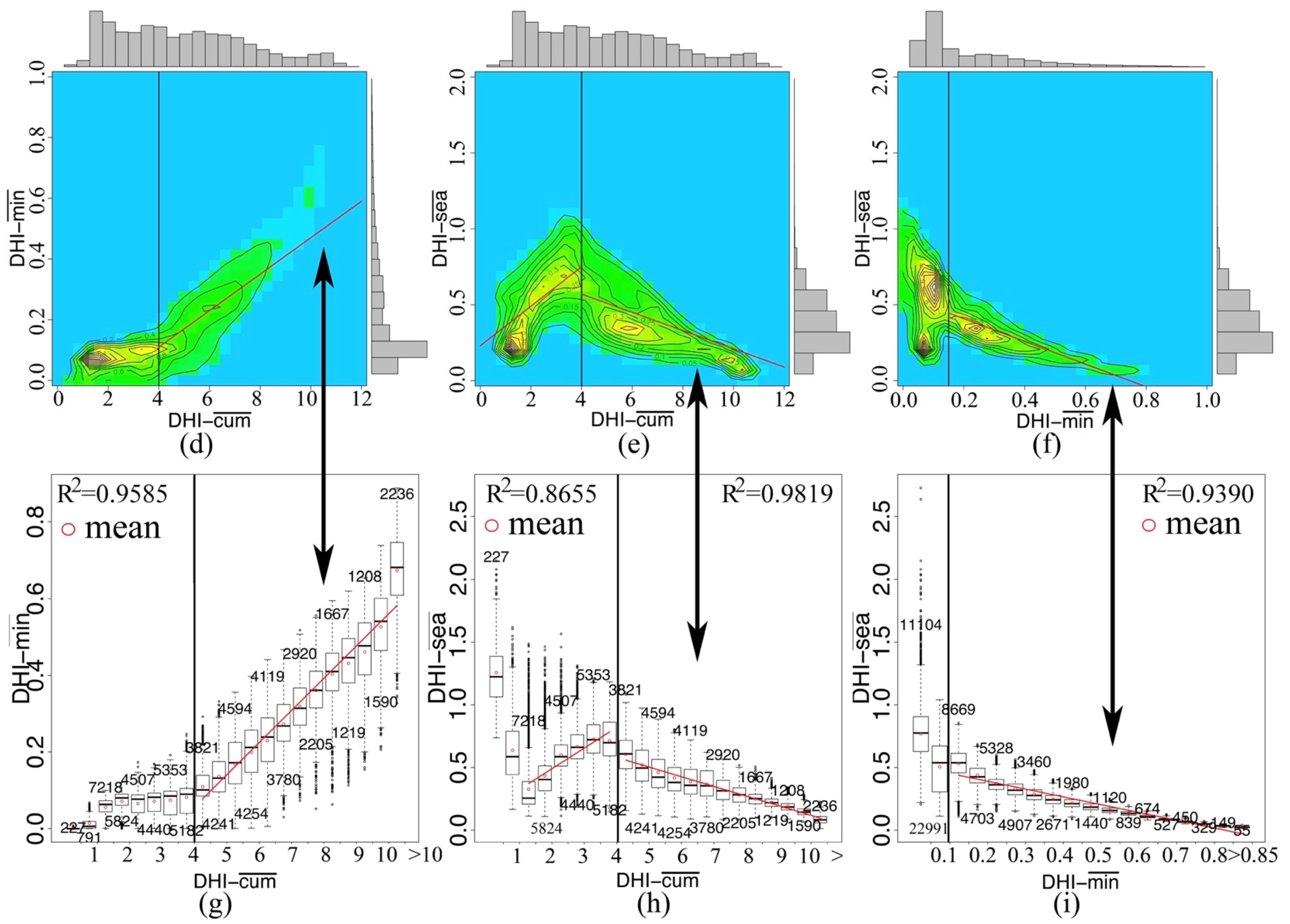
3.4. Regression and Change
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koh, L.P.; Dunn, R.R.; Sodhi, N.S.; Colwell, R.K.; Proctor, H.C.; Smith, V.S. Species coextinctions and the biodiversity crisis. Science 2004, 305, 1632–1634. [Google Scholar] [CrossRef] [PubMed]
- Dirzo, R.; Raven, P.H. Global state of biodiversity and loss. Ann. Rev. Environ. Resour. 2003, 28, 137–167. [Google Scholar] [CrossRef]
- Ehrlich, P.R.; Daily, G.C. Population extinction and saving biodiversity. Ambio 1993, 22, 64–68. [Google Scholar]
- Hughes, J.B.; Daily, G.C.; Ehrlich, P.R. Population diversity: Its extent and extinction. Science 1997, 278, 689–692. [Google Scholar] [CrossRef] [PubMed]
- May, R.M.; Tregonning, K. Global conservation and UK government policy. In Conservation in a Changing World; Cambridge University Press: London, UK, 1998; pp. 287–301. [Google Scholar]
- Daily, G. Nature’s Services: Societal Dependence on Natural Ecosystems; Island Press: Washington, DC, USA, 1997. [Google Scholar]
- Larigauderie, A.; Prieur-Richard, A.-H.; Mace, G.M.; Lonsdale, M.; Mooney, H.A.; Brussaard, L.; Cooper, D.; Cramer, W.; Daszak, P.; Díaz, S. Biodiversity and ecosystem services science for a sustainable planet: The diversitas vision for 2012–2020. Curr. Opin. Environ. Sust. 2012, 4, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, D.J. Six biological reasons why the endangered species act doesn’t work—And what to do about it. Conserv. Biol. 1991, 5, 273–282. [Google Scholar] [CrossRef]
- Telford, L. Update on endangered species protection in Canada. Endanger. Spec. Update 2000, 94–99. [Google Scholar]
- Zhang, P.; Shao, G.; Zhao, G.; Le Master, D.C.; Parker, G.R.; Dunning, J.B.; Li, Q. China’s forest policy for the 21st century. Science 2000, 288, 2135–2136. [Google Scholar] [CrossRef] [PubMed]
- Uchida, E.; Xu, J.; Rozelle, S. Grain for green: Cost-effectiveness and sustainability of China’s conservation set-aside program. Land Econ. 2005, 81, 247–264. [Google Scholar] [CrossRef]
- Xu, W.; Ouyang, Z.; Viña, A.; Zheng, H.; Liu, J.; Xiao, Y. Designing a conservation plan for protecting the habitat for giant pandas in the Qionglai mountain range, China. Divers. Distrib. 2006, 12, 610–619. [Google Scholar] [CrossRef]
- Gong, M.; Yu, C. Study on the Corridors of Giant Panda; China Forestry Publishing House: Beijing, China, 2003. [Google Scholar]
- Zhu, C.; Feng, G. A Case Study on China’s Policy of Converting Steep Cultivated Land to Forest or Grassland; China’s Forestry Press: Beijing, China, 2002. [Google Scholar]
- Slavin, R.E. Best evidence synthesis: An intelligent alternative to meta-analysis. J. Clin. Epidemiol. 1995, 48, 9–18. [Google Scholar] [CrossRef]
- Kerr, J.T.; Southwood, T.; Cihlar, J. Remotely sensed habitat diversity predicts butterfly species richness and community similarity in Canada. Proc. Nat. Acad. Sci. USA 2001, 98, 11365–11370. [Google Scholar] [CrossRef] [PubMed]
- Livingston, M.; Shaw, W.W.; Harris, L.K. A model for assessing wildlife habitats in urban landscapes of eastern Pima county, Arizona (USA). Landsc. Urban Plann. 2003, 64, 131–144. [Google Scholar] [CrossRef]
- Pauleit, S.; Ennos, R.; Golding, Y. Modeling the environmental impacts of urban land use and land cover change—A study in Merseyside, UK. Landsc. Urban Plann. 2005, 71, 295–310. [Google Scholar] [CrossRef]
- Turner, W.; Spector, S.; Gardiner, N.; Fladeland, M.; Sterling, E.; Steininger, M. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 18, 306–314. [Google Scholar] [CrossRef]
- Coops, N.C.; Wulder, M.A.; Duro, D.C.; Han, T.; Berry, S. The development of a Canadian dynamic habitat index using multi-temporal satellite estimates of canopy light absorbance. Ecol. Indic. 2008, 8, 754–766. [Google Scholar] [CrossRef]
- Mackey, B.G.; Bryan, J.; Randall, L. Australia’s dynamic habitat template 2003. In Proceedings of the MODIS Vegetation Workshop II, Missoula, MT, American, 17 April 2004.
- Michaud, J.-S.; Coops, N.C.; Andrew, M.E.; Wulder, M.A. Characterising spatiotemporal environmental and natural variation using a dynamic habitat index throughout the province of Ontario. Ecol. Indic. 2012, 18, 303–311. [Google Scholar] [CrossRef]
- Whittaker, R.J. Meta-analyses and mega-mistakes: Calling time on meta-analysis of the species richness-productivity relationship. Ecology 2010, 91, 2522–2533. [Google Scholar] [CrossRef] [PubMed]
- Duro, D.C.; Coops, N.C.; Wulder, M.A.; Han, T. Development of a large area biodiversity monitoring system driven by remote sensing. Prog. Phys. Geog. 2007, 31, 235–260. [Google Scholar] [CrossRef]
- Fritz, S.; McCallum, I.; Schill, C.; Perger, C.; See, L.; Schepaschenko, D.; van der Velde, M.; Kraxner, F.; Obersteiner, M. Geo-wiki: An online platform for improving global land cover. Environ. Modell. Softw. 2012, 31, 110–123. [Google Scholar] [CrossRef]
- Fritz, S.; See, L.; McCallum, I.; You, L.; Bun, A.; Moltchanova, E.; Duerauer, M.; Albrecht, F.; Schill, C.; Perger, C. Mapping global cropland and field size. Glob. Change Biol. 2015, 21, 1980–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- See, L.; McCallum, I.; Fritz, S.; Perger, C.; Kraxner, F.; Obersteiner, M.; Baruah, U.D.; Mili, N.; Kalita, N.R. Mapping cropland in Ethiopia using crowdsourcing. Int. J. Geosci. 2013, 4, 6–13. [Google Scholar] [CrossRef]
- Berry, S.; Mackey, B.; Brown, T. Potential applications of remotely sensed vegetation greenness to habitat analysis and the conservation of dispersive fauna. Pac. Cons. Biol. 2007, 13, 120–127. [Google Scholar]
- Coops, N.C.; Fontana, F.M.A.; Harvey, G.K.A.; Nelson, T.A.; Wulder, M.A. Monitoring of a national-scale indirect indicator of biodiversity using a long time-series of remotely sensed imagery. Can. J. Remote Sens. 2014, 40, 179–191. [Google Scholar] [CrossRef]
- Coops, N.C.; Wulder, M.A.; Iwanicka, D. Demonstration of a satellite-based index to monitor habitat at continental-scales. Ecol. Indic. 2009, 9, 948–958. [Google Scholar] [CrossRef]
- Coops, N.C.; Waring, R.H.; Wulder, M.A.; Pidgeon, A.M.; Radeloff, V.C. Bird diversity: A predictable function of satellite-derived estimates of seasonal variation in canopy light absorbance across the United States. J. Biogeogr. 2009, 36, 905–918. [Google Scholar] [CrossRef]
- Holmes, K.R.; Nelson, T.A.; Coops, N.C.; Wulder, M.A. Biodiversity indicators show climate change will alter vegetation in parks and protected areas. Diversity 2013, 5, 352–373. [Google Scholar] [CrossRef]
- Andrew, M.E.; Wulder, M.A.; Coops, N.C.; Baillargeon, G. Beta-diversity gradients of butterflies along productivity axes. Glob. Ecol. Biogeogr. 2012, 21, 352–364. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, Y.; Knyazikhin, Y.; Myneni, R.B.; Glassy, J.M.; Dedieu, G.; Running, S.W. Prototyping of MODIS LAI and fPAR algorithm with LASUR and Landsat data. IEEE Trans. Geosci. Remote Sens. 2000, 38, 2387–2401. [Google Scholar] [CrossRef]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. PNAS 2013, 110, 2602–2610. [Google Scholar] [CrossRef] [PubMed]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Mapping the World’s Biodiversity. Available online: http://www.biodiversitymapping.org (accessed on 10 November 2014).
- Evans, K.L.; Warren, P.H.; Gaston, K.J. Species–energy relationships at the macroecological scale: A review of the mechanisms. Biol. Rev. 2005, 80, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Skidmore, A.K.; Oindo, B.O.; Said, M.Y. Biodiversity assessment by remote sensing. In Proceedings of the 30th International Symposium on Remote Sensing of the Environment: Information for Risk Management and Sustainable Development, Honolulu, HI, USA; 2003. [Google Scholar]
- Schwartz, C.C.; Haroldson, M.A.; White, G.C.; Harris, R.B.; Cherry, S.; Keating, K.A.; Moody, D.; Servheen, C. Temporal, spatial, and environmental influences on the demographics of grizzly bears in the greater Yellowstone ecosystem. Wildlife Monogr. 2006, 161, 1–8. [Google Scholar] [CrossRef]
- Gilmore, S.; Mackey, B.; Berry, S. The extent of dispersive movement behaviour in Australian vertebrate animals, possible causes, and some implications for conservation. Pac. Cons. Biol. 2007, 13, 93–103. [Google Scholar]
- McLoughlin, P.D.; Wal, E.V.; Lowe, S.J.; Patterson, B.R.; Murray, D.L. Seasonal shifts in habitat selection of a large herbivore and the influence of human activity. Basic Appl. Ecol. 2011, 12, 654–663. [Google Scholar] [CrossRef]
- Holben, B.N. Characteristics of maximum-value composite images from temporal AVHRR data. Int. J. Remote Sens. 1986, 7, 1417–1434. [Google Scholar] [CrossRef]
- Rao, C.R.; Toutenburg, H.; Shalabh, H.C.; Schomaker, M. Linear Models and Generalizations. Least Squares and Alternatives, 3rd ed.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Mamtimin, B.; Et-Tantawi, A.M.M.; Schaefer, D.; Meixner, F.X.; Domroes, M. Recent trends of temperature change under hot and cold desert climates: Comparing the Sahara (Libya) and central Asia (Xinjiang, China). J. Arid. Environ. 2011, 75, 1105–1113. [Google Scholar] [CrossRef]
- López-Pujol, J.; Zhang, F.-M.; Ge, S. Plant biodiversity in China: Richly varied, endangered, and in need of conservation. Biodivers Conserv. 2006, 15, 3983–4026. [Google Scholar] [CrossRef]
- Cai, D.; Guan, Y.; Guo, S.; Zhang, C.; Fraedrich, K. Mapping plant functional types over broad mountainous regions: A hierarchical soft time-space classification applied to the Tibetan Plateau. Remote Sens. 2014, 6, 3511–3532. [Google Scholar] [CrossRef]
- Yi, X.; Yin, Y.; Li, G.; Peng, J. Temperature variation in recent 50 years in the three-river headwaters region of Qinghai province. Acta Geogr. Sin. 2011, 66, 1451–1465. (In Chinese) [Google Scholar]
- Harkness, J. Recent trends in forestry and conservation of biodiversity in China. China Q. 1998, 156, 911–934. (In Chinese) [Google Scholar] [CrossRef]
- Nilsen, E.B.; Herfindal, I.; Linnell, J.D.C. Can intra-specific variation in carnivore home-range size be explained using remote-sensing estimates of environmental productivity? Ecoscience 2005, 12, 68–75. [Google Scholar] [CrossRef]
- Cai, D.; Fraedrich, K.; Sielmann, F.; Guan, Y.; Guo, S.; Zhang, L.; Zhu, X. Climate and vegetation: An ERA-interim and GIMMS NDVI analysis. J. Clim. 2014, 27, 5111–5118. [Google Scholar] [CrossRef]
- Cai, D.; Fraedrich, K.; Sielmann, F.; Zhang, L.; Zhu, X.; Guo, S.; Guan, Y. Vegetation dynamics on the Tibetan Plateau (1982 to 2006): An attribution by eco-hydrological diagnostics. J. Clim. 2015, 28, 4576–4584. [Google Scholar] [CrossRef]
- National Bureau of Statistics of the People’s Republic of China. Available online: http://www.stats.gov.cn (accessed on 10 June 2013).
- Jetz, W.; Fine, P.V.A. Global gradients in vertebrate diversity predicted by historical area-productivity dynamics and contemporary environment. PLoS Biol. 2012, 10, e1001292. [Google Scholar] [CrossRef] [PubMed]
- Jetz, W.; Rahbek, C. Geographic range size and determinants of avian species richness. Science 2002, 297, 1548–1551. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Kissling, W.D. Spatial scale and cross-taxon congruence of terrestrial vertebrate and vascular plant species richness in China. Ecology 2010, 91, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Waide, R.B.; Willig, M.R.; Steiner, C.F.; Mittelbach, G.; Gough, L.; Dodson, S.I.; Juday, G.P.; Parmenter, R. The relationship between productivity and species richness. Annu. Rev. Ecol. Syst. 1999, 30, 257–300. [Google Scholar] [CrossRef]
- Chase, J. Historical and contemporary factors govern global biodiversity patterns. PLoS Biol. 2012, 10, e1001294. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.E.; Stoms, D.M.; Estes, J.E.; Cayocca, K.D. Relationships between Biological Diversity and Multi-Temporal Vegetation Index Data in California; Soc Photogrammetry & Remote Sensing: Washington, DC, USA, 1992; pp. 562–571. [Google Scholar]
- Moore, J.C.; De Ruiter, P.C.; Hunt, H.W. Influence of productivity on the stability of real and model ecosystems. Sci. New York Then Wash. 1993, 261, 906–908. [Google Scholar] [CrossRef] [PubMed]
- Chown, S.L.; van Rensburg, B.J.; Gaston, K.J.; Rodrigues, A.S.L.; van Jaarsveld, A.S. Energy, species richness, and human population size: Conservation implications at a national scale. Ecol. Appl. 2003, 13, 1233–1241. [Google Scholar] [CrossRef] [Green Version]
- Mao, D.; Wang, Z.; Luo, L.; Ren, C. Integrating AVHRR and MODIS data to monitor NDVI changes and their relationships with climatic parameters in Northeast China. Int. J. Appl. Earth Obs. 2012, 18, 528–536. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, S.; Zhou, Z.; Wang, R. Vegetation map of the People’s Republic of China (1:1,000,000); Geological Publishing House: Beijing, China, 2007. (In Chinese) [Google Scholar]
- Chen, C.; Qian, C.; Deng, A.; Zhang, W. Progressive and active adaptations of cropping system to climate change in Northeast China. Eur. J. Agron. 2012, 38, 94–103. [Google Scholar] [CrossRef]
- Liu, M.; Liu, G.H.; Wu, X.; Wang, H.; Chen, L. Vegetation traits and soil properties in response to utilization patterns of grassland in Hulun Buir city, Inner Mongolia, China. Chinese Geogr. Sci. 2014, 24, 471–478. [Google Scholar] [CrossRef]
- Lu, A.; Kang, S.; Pang, D.; Wang, T.; Ge, J. Different landform effects on seasonal temperature patterns in China. Ecol. Envrion. 2008, 17, 1450–1452. [Google Scholar]
- Wang, J.; Huang, J.; Yang, J. Overview of impacts of climate change and adaptation in China’s agriculture. J. Integr. Agric. 2014, 13, 1–17. [Google Scholar] [CrossRef]
- Li, J.; Yu, Q.; Sun, X.; Tong, X.; Ren, C.; Wang, J.; Liu, E.; Zhu, Z.; Yu, G. Carbon dioxide exchange and the mechanism of environmental control in a farmland ecosystem in North China Plain. Sci. China Ser. D 2006, 49, 226–240. [Google Scholar] [CrossRef]
- Mo, X.; Liu, S.; Lin, Z. Evaluation of an ecosystem model for a wheat–maize double cropping system over the North China plain. Environ. Modell. Softw. 2012, 32, 61–73. [Google Scholar] [CrossRef]
- Song, Y.; Liu, B.; Zhong, H. Impact of global warming on the rice cultivable area in southern China in 1961–2009. Adv. Clim. Chang. Res. 2011, 7, 259–264. (In Chinese) [Google Scholar]
- Wang, J.; Wang, E.; Yang, X.; Zhang, F.; Yin, H. Increased yield potential of wheat-maize cropping system in the North China Plain by climate change adaptation. Climat. Chang. 2012, 113, 825–840. [Google Scholar] [CrossRef]
- Guo, Z. Study on Comprehensive Benefits Evaluation of Natural Forest Protection Program in Southwest China. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2011. [Google Scholar]
- Huang, C.; Zhuang, X.; Li, R.; Liu, Z.; Jiang, B.; Zhai, C. Damaged status and early recovery of tree species in Wuzhishan of Nanling Mountains, South China after the ice storm in 2008. Chin. J. Ecol. 2012, 6, 1390–1396. (In Chinese) [Google Scholar]
- Zhou, B.; Li, Z.; Wang, X.; Cao, Y.; An, Y.; Deng, Z.; Letu, G.; Wang, G.; Gu, L. Impact of the 2008 ice storm on moso bamboo plantations in Southeast China. J. Geophys. Res.: Biogeosci. 2011, 116, G00H06. [Google Scholar] [CrossRef]
- Barriopedro, D.; Gouveia, C.M.; Trigo, R.M.; Wang, L. The 2009/10 drought in China: possible causes and impacts on vegetation. J. Hydrometeorol. 2012, 13, 1251–1267. [Google Scholar] [CrossRef]
- Storch, D.; Davies, R.G.; Zajíček, S.; Orme, C.D.L.; Olson, V.; Thomas, G.H.; Ding, T.-S.; Rasmussen, P.C.; Ridgely, R.S.; Bennett, P.M.; et al. Energy, range dynamics and global species richness patterns: Reconciling mid-domain effects and environmental determinants of avian diversity. Ecol. Lett. 2006, 9, 1308–1320. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Fraedrich, K.F.; Sielmann, C.D.; Zhu, X. Climate dynamics on watershed scale: Along the rainfall-runoff chain. In The Fluid Dynamics of Climate, International Centre for Mechanical Sciences (CISM); Springer Verlag: Vienna, Austria, 2016; pp. 183–209. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
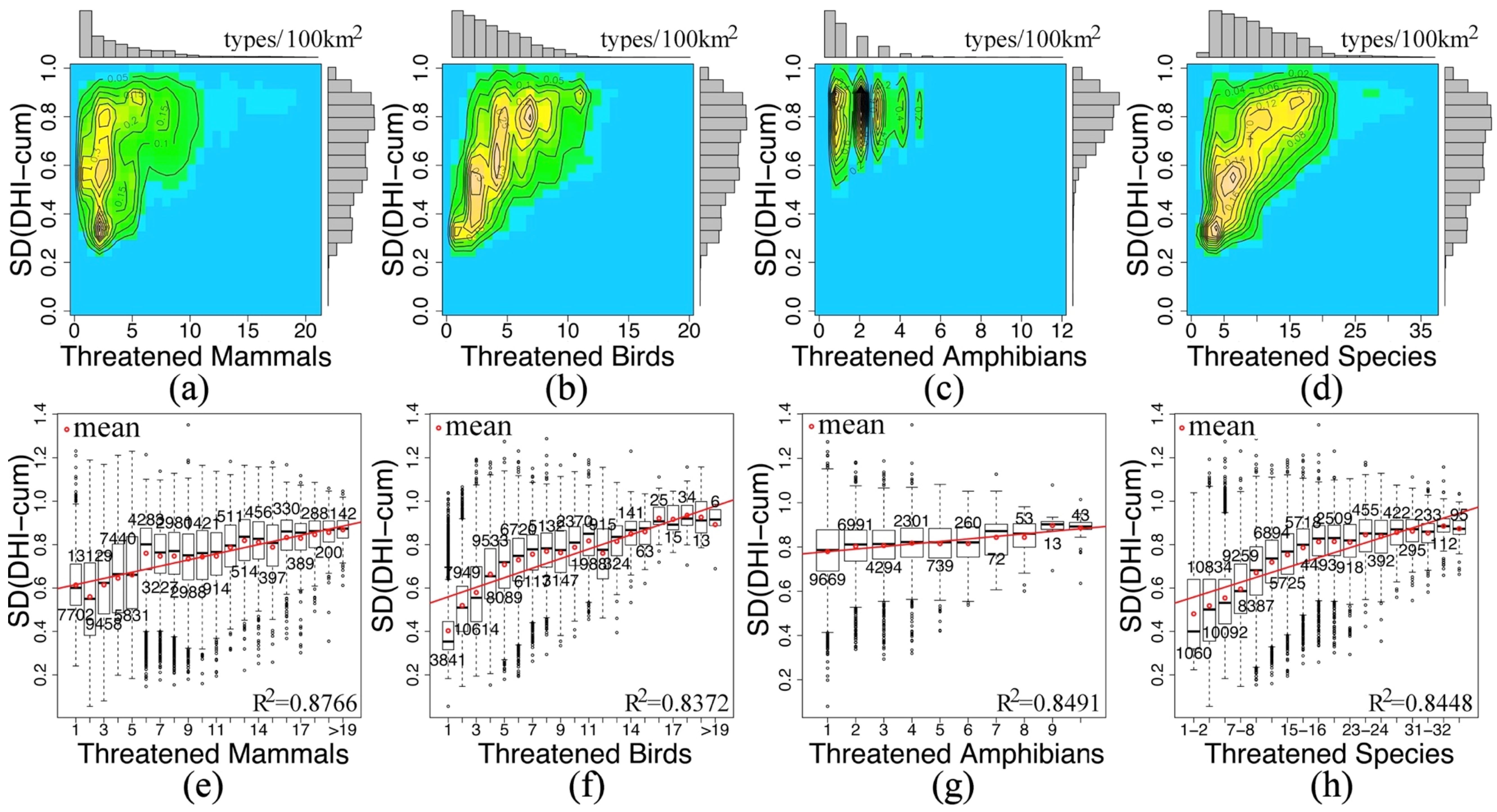
| Species Richness | Slopes | R2 | p Value | DF | Pixels |
|---|---|---|---|---|---|
| Mammals | 0.4516 | 0.8538 | 5.34e-07 | 13 | 64817 |
| Birds | 0.7980 | 0.9289 | 1.125e-06 | 9 | 64803 |
| Amphibians | 0.5586 | 0.8124 | 0.0002 | 8 | 56219 |
| ALL | 0.5393 | 0.8611 | 1.062e-06 | 12 | 71380 |
| Threatened Species | Slopes | R2 | p Value | DF | Pixels |
|---|---|---|---|---|---|
| Mammals | 0.0142 | 0.8766 | 7.989e-10 | 18 | 62598 |
| Birds | 0.0220 | 0.8372 | 9.895e-09 | 18 | 67034 |
| Amphibians | 0.0115 | 0.8491 | 9.364e-05 | 8 | 24433 |
| ALL | 0.0227 | 0.8448 | 4.367e-08 | 16 | 67891 |
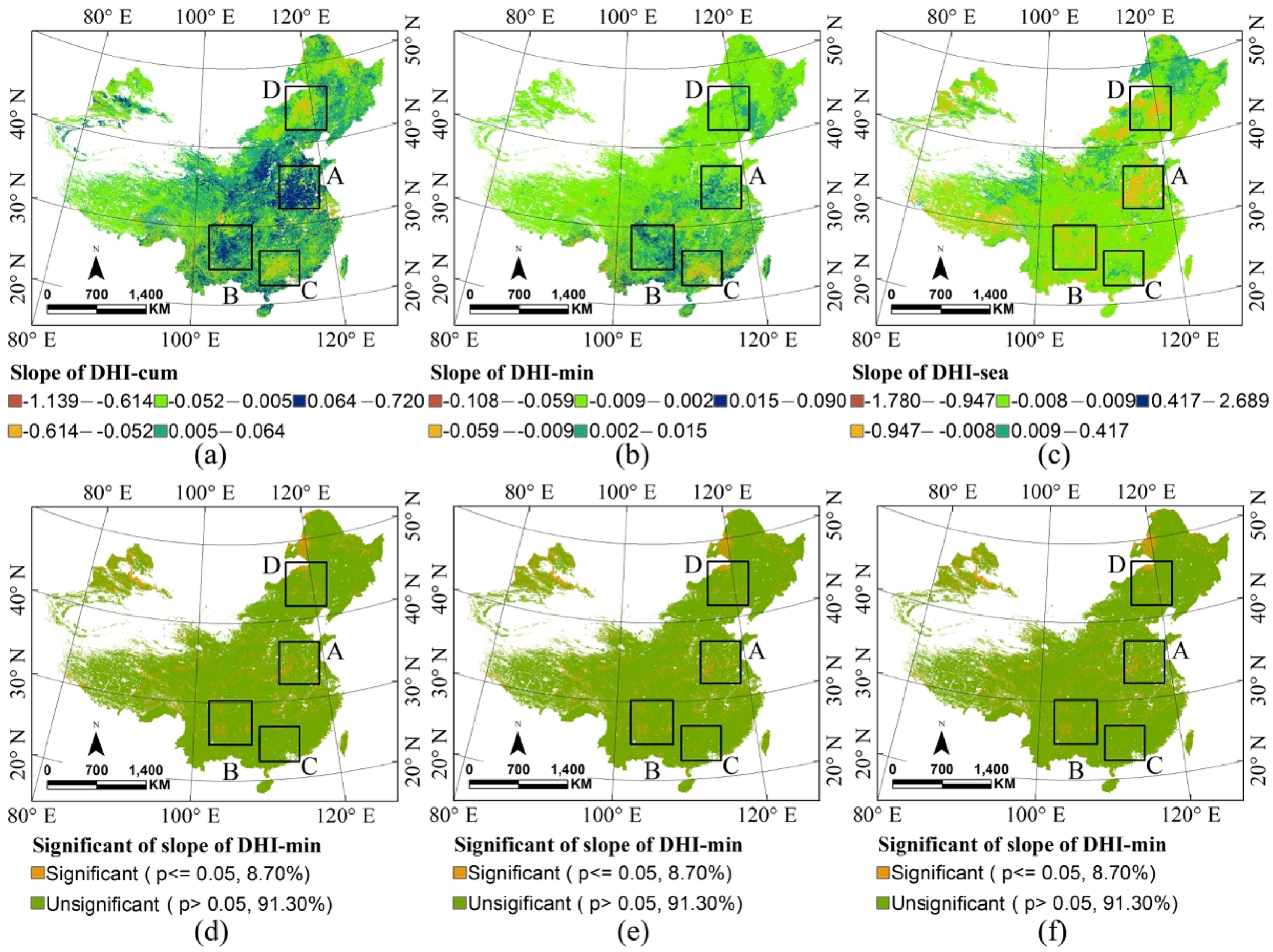
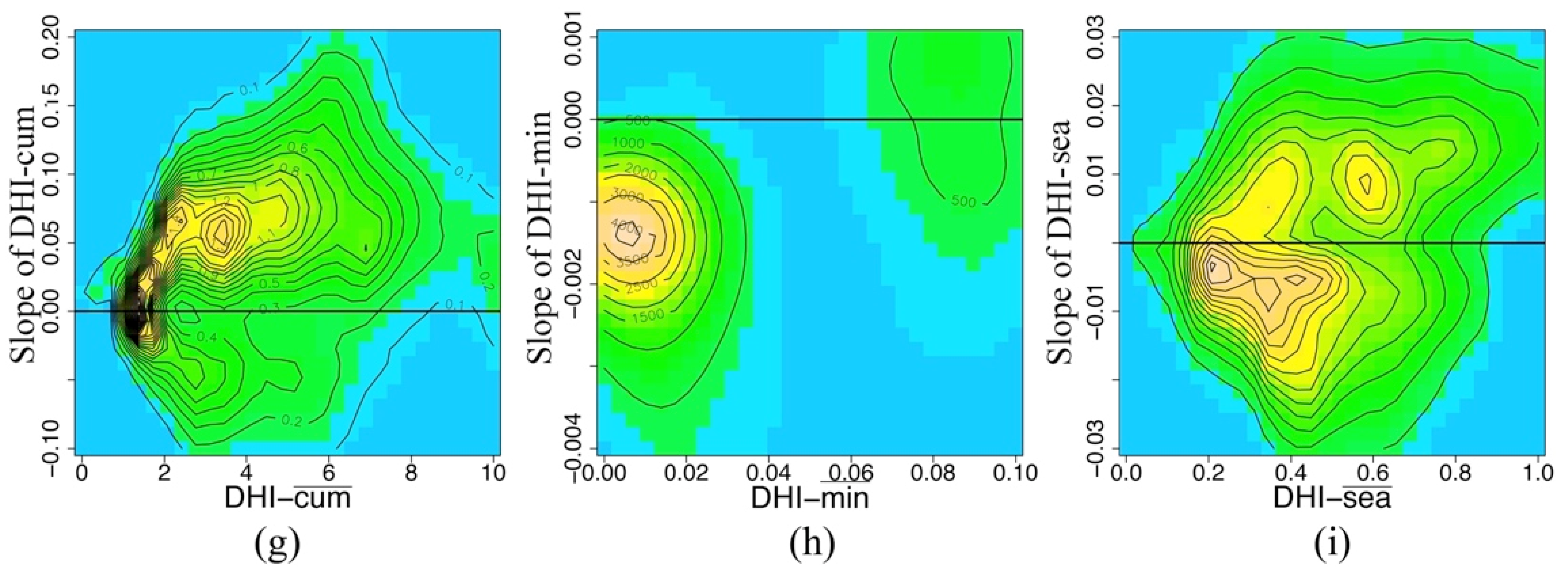
| Significant | Slope of DHI-cum | Slope of DHI-min | Slope of DHI-sea |
|---|---|---|---|
| p ≤ 0.05 | 15.76% | 8.70% | 9.85% |
| p > 0.05 | 84.24% | 91.30% | 90.15% |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Cai, D.; Guo, S.; Guan, Y.; Fraedrich, K.; Nie, Y.; Liu, X.; Bian, X. Spatial-Temporal Dynamics of China’s Terrestrial Biodiversity: A Dynamic Habitat Index Diagnostic. Remote Sens. 2016, 8, 227. https://0-doi-org.brum.beds.ac.uk/10.3390/rs8030227
Zhang C, Cai D, Guo S, Guan Y, Fraedrich K, Nie Y, Liu X, Bian X. Spatial-Temporal Dynamics of China’s Terrestrial Biodiversity: A Dynamic Habitat Index Diagnostic. Remote Sensing. 2016; 8(3):227. https://0-doi-org.brum.beds.ac.uk/10.3390/rs8030227
Chicago/Turabian StyleZhang, Chunyan, Danlu Cai, Shan Guo, Yanning Guan, Klaus Fraedrich, Yueping Nie, Xuying Liu, and Xiaolin Bian. 2016. "Spatial-Temporal Dynamics of China’s Terrestrial Biodiversity: A Dynamic Habitat Index Diagnostic" Remote Sensing 8, no. 3: 227. https://0-doi-org.brum.beds.ac.uk/10.3390/rs8030227