Advanced Methods for Detection of Bacillus cereus and Its Pathogenic Factors



INRAE, AgroParisTech, Micalis Institute, Université Paris-Saclay, 78350 Jouy-en-Josas, France
*
Authors to whom correspondence should be addressed.
Sensors 2020, 20(9), 2667; https://0-doi-org.brum.beds.ac.uk/10.3390/s20092667
Submission received: 21 April 2020
/
Revised: 2 May 2020
/
Accepted: 5 May 2020
/
Published: 7 May 2020
(This article belongs to the Special Issue Advanced Biosensors for Bacterial Detection)
Abstract
:Bacillus cereus is an opportunistic foodborne pathogen causing food intoxication and infectious diseases. Different toxins and pathogenic factors are responsible for diarrheal syndrome, like nonhemolytic enterotoxin Nhe, hemolytic enterotoxin Hbl, enterotoxin FM and cytotoxin K, while emetic syndrome is caused by the depsipeptide cereulide toxin. The traditional method of B. cereus detection is based on the bacterial culturing onto selective agars and cells enumeration. In addition, molecular and chemical methods are proposed for toxin gene profiling, toxin quantification and strain screening for defined virulence factors. Finally, some advanced biosensors such as phage-based, cell-based, immunosensors and DNA biosensors have been elaborated to enable affordable, sensitive, user-friendly and rapid detection of specific B. cereus strains. This review intends to both illustrate the state of the B. cereus diagnostic field and to highlight additional research that is still at the development level.
1. Introduction
Since the beginning of the 20th century, the role of Bacillus cereus in food-poisoning has been clearly demonstrated [1,2,3]. In Europe, it is the second cause (as confirmed and suspected causative agent) of the foodborne outbreak (FBO) after Staphylococcus aureus [4]. In 2017, 44% of the reported cases of FBO in France were caused by B. cereus. Nevertheless, the impact of B. cereus in FBO is most likely underestimated due to the lack of systematic surveillance and is often misdiagnosed as S. aureus or Clostridium perfringens symptom-like infections.
B. cereus can cause emetic or diarrheal-type of foodborne illnesses, which are generally mild and self-limiting. However, severe systemic infections have also been associated with this pathogen. Although rare, these infections may lead to the death of newborns and immunologically-compromised or vulnerable individuals [4,5].
The pathogenicity potential of B. cereus is extremely variable, with strains being harmless and others lethal. This variability makes it difficult to communicate around the dangerousness of B. cereus and often leads to misunderstanding and mismanagement of the associated risks. Therefore, the characterization of B. cereus potential of pathogenicity is a major challenge for the agri–food industries and hospitals. Due to the lack of validated and standardized analytical methods to assess the presence of specific toxins, only the presence of presumptive B. cereus is usually examined. Therefore, during the last decade, the elaboration of new diagnostic tools for toxin gene profiling and for toxin quantification has gained increasing importance. It is thought that the application of such new methods will lead to a significant improvement of B. cereus diagnostics and patient care. In this review, we provide an overview of the traditional methods currently used in the food industry and at hospitals to detect B. cereus. We also present advanced methods developed at a bench-scale that allow a specific, rapid and sensitive detection of B. cereus strains and their pathogenic factors. We recently reviewed methods for the detection of B. cereus spores [6].
2. Pathogenicity of Bacillus cereus
2.1. Bacillus cereus Group
B. cereus are spore-forming, Gram positive, aerobic or facultative anaerobic bacteria. The presence of a peritrichous ciliature allows the bacteria to be motile [7]. Morphologically, B. cereus strains are thin, straight or slightly curved bacilli of 1 × 3–4 µm with square ends, which can form chains (Figure 1).
B. cereus sensus stricto belongs to a larger group of bacteria commonly named the Bacillus cereus group. The group is constituted of eight species: B. mycoides, B. pseudomycoides, B. weihenstephanensis, B. anthracis, B. thuringiensis, B. cereus sensu stricto (usually called B. cereus), B. cytotoxicus and B. toyonensis. Recently, the group was extended with new species, which were all isolated from marine sediments: B. paranthracis, B. pacificus, B. tropicus, B. albus, B. mobilis, B. luti, B. proteolyticus, B. nitratireducens and B. paramycoides [8], while some other strains are under evaluation [6]. Differentiation of species within the B. cereus group is complex due to the genetic proximity between the members of the group [9,10]. Originally, species in the group were classified on the basis of phenotypic differences, distinct virulence trait and the presence of extrachromosomal elements that reflect the specie’s virulence spectrum. Currently, the most widespread classification system of this bacteria group is based on the sequencing of panC housekeeping gene, which encodes for the pantoate-beta-alanine ligase C [11]. Using this classification, seven phylogenetic groups have been determined based principally on their range of growth temperatures. B. cereus can be found in several clusters, among which the groups III and VII comprise of species associated with high cytotoxicity [12].
B. cereus are ubiquitous bacteria. Besides the soil which is their primary reservoir, they can be isolated from vegetation and waters [13,14] and can colonize insects and mammals [15,16,17]. From the environment, they can be transferred into various raw materials used in the food industry. The host is contaminated by spores or vegetative cells present in ingested food, inhaled air, or entering the body through a wound. In addition, these microorganisms can deteriorate the organoleptic qualities of food (in particular eggs and pasteurized milk) with an impact on the market quality of the products, which must then be destroyed. B. cereus, therefore, constitutes a real public health problem and represents a major economic risk for the food industry.
However, currently, no legislation requires systematic screening of food items for this pathogen contamination. Within the EU, the only regulation implying a safety limit for B. cereus in foods concern dried infant formulae with an established maximum limit of 50 CFU/g (Commission regulation (EC) No 1441/2007). In France, the regulatory limit of B. cereus presence in food, especially starch-rich foods have been set to 1 × 105 CFU/g of food (Act DGAL/MUS/N2009-8188, 07 July 2009). According to the Codex Alimentarius Commission of the Food and Agriculture Organization of the United Nations (FAO) and the World Health Organization (WHO) Codex standard for infant formula, the maximum acceptable number of B. cereus is 102 CFU/mL [18]. Most cases of FBO due to B. cereus have been associated with a bacterial concentration above 105 CFU/g of food material but some cases have been linked with bacterial concentration as low as 103 CFU/g [2,19]. Moreover, the determination of a safety limit is difficult, as the pathogenicity does not depend solely on the number of bacterial cells [20,21,22,23].
2.2. B. cereus Gastro-Intestinal Infections
As previously mentioned, there are two types of food poisoning due to B. cereus, which differ in terms of symptoms and origin, known as the emetic and the diarrheal syndromes. The latter is associated with the production of enterotoxins in the small intestine following the ingestion of contaminated food. Bacteria can be ingested as vegetative cells or as spores, which once in the intestine, germinate, multiply, and produce enterotoxins [6,16,24]. Symptoms appear 5 h to 24 h after ingestion, and manifest through abdominal pain and profuse diarrhea [14]. Manifestations are generally mild with spontaneous remission within 24 h without any treatment. However, fatal cases resulting for example from necrotic enteritis have been reported [25].
The emetic syndrome results from an intoxication provoked by the emetic toxin cereulide, a toxin often pre-formed in food before ingestion [26]. This toxin causes nausea and vomiting that occur quickly (between 30 min and 6 h after ingestion) and has been linked to severe clinical forms associated with fatalities, in particular, because of liver failure [27,28,29,30].
2.3. B. cereus Non Gastro-Intestinal Infections
In addition to FBO, B. cereus is responsible for a variety of local and systemic infections, which can lead to the death of patients in approximately 10% of cases [5,7]. These clinical infections have undeniable repercussions on public health, in particular for fragile populations such as premature newborns. In addition, several cases of fulminant infections with B. cereus, similar to anthrax, and affecting healthy persons, have also been reported [31].
However, our recent epidemiological study, together with the alert launched by the Paris hospitals (AP-HP) and the health institutions have shown that the number of cases of serious B. cereus infections is largely underestimated and underdocumented, while the first-line treatment is not always suitable [5]. Many hospitals face deaths of newborns from B. cereus-related bacteremia or meningitis [32,33,34].
Taken together, B. cereus toxigenic bacteria are on the rise and have to be considered as emerging threats, especially for high-risk population, where adequate methods for their detection remain scarce.
3. Detection of B. cereus
3.1. Traditional Methods
The traditional method for B. cereus detection is agar plate-based counting for which the guidelines are provided by the ISO 7932:2004 that had been last revised nearly 15 years ago. Several laborious sub-points are needed, like: sample homogenization, numerous centrifugations and serial dilution preparations, all carried out in aseptic conditions to avoid any contamination [35]. The plating steps have to be carried out in duplicate for each dilution using specific media. Then, plates need to be incubated at 30 °C for 24 h to 48 h. Finally, colonies have to be streaked onto the Brain Heart Infusion (BHI) agar medium. Moreover, an additional confirmation assay is required in order to differentiate bacteria of the B. cereus group. This last step takes another 2 h to 24 h. Traditional methods are, thus, time-consuming, laborious, expensive and require trained staff capable of ensuring the correct procedure. Furthermore, with these methodologies, it is not possible to detect injured or viable but not-culturable (VBNC) cells. The ISO 7932:2004 method specifies a horizontal test for the enumeration of presumptive B. cereus cells. This method consists of a serial dilution of the samples and a subsequent dilution spreading on B. cereus selective Mossel medium agar (MYP). Incubation of the colonies at 30 °C during 18–24 h is needed. On MYP medium, colonies of B. cereus have a pink–purple color, surrounded by a characteristic halo formed of pink precipitation, which permits their identification. Finally, a hemolysis test is performed on bacterial colonies to confirm for B. cereus strains. This method confirms only presumptive B. cereus cells because not all strains of B. cereus are hemolytic.
More recently, alternative methods based on the NF EN ISO 7932 standard for the enumeration of presumptive B. cereus in food have been validated. The AFNOR BKR 23-06-02-10 and AFNOR AES-10/10-07/10 methods use a selective chromogenic medium, COMPASS and BACARA respectively. After hydrolyzation of the chromogenic substrate, B. cereus colonies appear in green (COMPASS) or in orange surrounded by an opaque halo (BACARA). These selective agars inhibit the majority of the background microflora and allow easy identification. Their specificity, selectivity and accuracy are comparable to the reference method. In addition, they are time-saving and do not involve a confirmation step.
3.2. Molecular Methods
Molecular methods used for B. cereus identification provide many advantages compared to the traditional methods such as versatility, lower time and resource consumption, and high specificity [36]. In the past decades, different techniques based on polymerase chain reaction (PCR) have been developed for B. cereus detection, such as Nested PCR [37], Randomly Amplified Polymorphic DNA PCR (RAPD PCR) [38] and Real-Time PCR (RTi-PCR) [39]. The amplification step is coupled to different electrophoresis procedures such as Pulse Field Gel Electrophoresis (PFGE) [40], Temporal Temperature Gradient gel Electrophoresis (TTGE) [41] and Denaturing Gradient Gel Electrophoresis (DGGE) [42]. The coliphage M13 sequence-based polymerase chain reaction (M13-PCR) and DNA amplification fingerprinting were applied to obtain profiles of various B. cereus strains [43]. The resulting discriminative and informative profiles have permitted us to trace the spread of B. cereus contamination in a food plant. Hall et al., [38] were able to characterize the microbiological hazard of products provided by vending machines with the use of PCR and RAPD PCR. Using specific primers that target the gyrB gene encoding DNA gyrase subunit B, they found out that almost 90% of bacteria isolated from hot chocolate powder and hot chocolate drinks of vending machine belonged to B. cereus group. In another study, Fernandez-No et al., [39] were able to simultaneously quantify B. cereus, B. subtilis and B. licheniformis by means of RTi-PCR. By using probes targeting the 16S rRNA gene of each bacterium, the RTi-PCR could detect each specific strains with a limit of detection (LOD) of 165 CFU/g of an artificially contaminated pasteurized foodstuff.
Over the years, new methods have been developed with the leading principle to detect and distinguish B. cereus from other Bacillus group members by a time-saving and in-situ analysis. For example, Manzano et al., [41] compared different molecular methods that use specific probes and primers as recognition elements to distinguish B. cereus from B. thuringiensis from different origins (food, clinical and bio-pesticide). They used three different strategies: a PCR-TTGE technique targeting the gyrB gene, a rep-PCR in which two specific DNA sequences were targeted, and a RAPD-PCR uses M13 primers. They demonstrated that only the rep-PCR was able to cluster strains of the same origin [41]. This work highlights the difficulty to differentiate B. cereus and B. thuringiensis even with molecular methods. Noguera and Ibarra [44] designed universal primers to amplify cry genes. As B. thuringiensis is the only strain within the B. cereus group to produce the cry toxin proteins, this technique has permitted to discriminate B. thuringiensis from other B. cereus strains [44]. Another method used to distinguish B. cereus from other members of the B. cereus group is the genotyping using high-resolution melting analysis [45]. This analysis allows the discrimination of B. cereus by typing an amplified polymorphic 16S-23S intergenic spacer region. This method has been proposed for the control of food safety following adequate sanitizing treatments. Nevertheless, the method cannot dissociate B. cereus from B. thuringiensis.
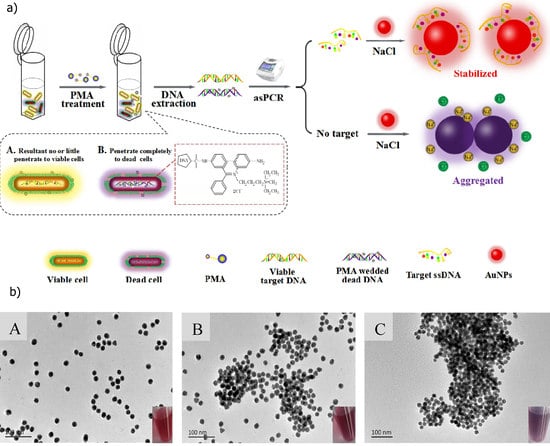
Recently, an innovative approach to increase the rapidity of post-PCR step has been proposed by Li et al., [36]. They replaced the detection of PCR products using the laborious electrophoretic method by a rapid visualization system using gold nanoparticles (AuNPs). They also applied propidium monoazide (PMA) prior to performing the asymmetric PCR (asPCR) amplification in order to only detect the viable B. cereus cells. PMA is a viability-photoreactive DNA binding dye that can penetrate inside the cell when the membrane permeability is altered. Thus, it can only bind to the DNA of dead cells. PMA, upon binding, permanently modifies DNA which in consequence cannot be amplified during the asPCR step. The cereulide synthetase encoding gene (cesB) was used as a specific B. cereus biomarker targeted during the asPCR. The PCR products were mixed with AuNPs and the resulting coloration used as an indicator of B. cereus presence (Figure 2). Indeed, upon salt addition and in the absence of amplicons, AuNPs aggregates and form a violet precipitate. In contrast, salt addition does not induce AuNP aggregation in the presence of the amplicons and the solution remained red. Using this protocol, a LOD of 9.2 × 101 CFU/mL in PBS and 3.4 × 102 CFU/mL in milk were respectively obtained. Besides demonstrating the feasibility of the test, this study pointed out the crucial parameters necessary to obtain a low LOD such as the salt concentration that induces aggregation of AuNPs and the length of the primers.
Despite the advantages offered by the molecular techniques, to overtake the traditional method drawbacks, such as rapid bacterial identification at lower analysis cost, the former exhibits several limitations. Indeed, trained professionals are required to perform this type of assays, which are still laborious (PCR test takes from 2 h to 24 h) and costly. Sometimes amplification errors occur, or false-negative results may be obtained when using complex matrices such as blood or food samples [46,47,48]. Another limitation of PCR is the use of laborious, time-consuming and low-resolution gel electrophoresis methods to visualize PCR products. One exception is microfluidic PCR chips that automatize protocol by combining sample processing, DNA extraction, enzymatic amplification and detection [49,50]. Qiu et al., [51] described an inexpensive PCR module constituted of a thermal cycler combined with a PCR chip that allows amplification and real-time fluorescence monitoring which can be used for the detection of B. cereus genomic DNA. In addition, primers can be conjugated with biotin and dioxigenin to enable lateral flow-based detection of amplicons as an alternative to the optical measurement. Using this module, the negative sample exhibited neither a fluorescent signal nor band. They also optimized the module to allow for the pre-storage of paraffin-encapsulated PCR reagents inside the PCR chamber to achieve the device automation. Thus, reagents can be released during the heating after the melting of paraffin and initiate amplification. This lab-on-a-chip PCR module carried all the necessary steps from sample-introduction to result-visualization.
Nevertheless, molecular methods are often fall short of providing an instant, point-of-care detection, and thus, they are not suitable for safety testing in agri-food chains. Especially, it is difficult to integrate PCR based methods into a point-of-care format because they require a thermal cycler instrument to amplify the target sequence. To overcome this technical obstacle, nucleic acid sequence amplification through isothermal methods have been developed with the use of enzymes for strand separation instead of repeated heating [52,53]. Xia et al., [54] developed a loop-mediated isothermal amplification (LAMP) method for simultaneous amplification of B. cereus and four other foodborne bacterial pathogens Escherichia coli, Salmonella enterica, Vibrio fluvialis and V. parahaemolyticus. In their device, reagents and samples were loaded into the reaction chambers, through inlets and microchannels, to reach the Peltier heater that maintains a temperature of 64 °C during the LAMP reactions. A portable CCD camera is used to capture the fluorescence of formed amplicants. Under optimized conditions, the simultaneous detection of all five pathogens was performed in 60 min and with a success rate of 100%.
3.3. Biosensors
In the last few years, biosensors have emerged as ideal methods for the detection of foodborne pathogens [6,52,53,55,56,57,58,59]. The success of these miniaturized devices is mainly due to their rapidity to provide results, simplicity, and possibility of point-of-care diagnosis together with the low cost and high reliability of analysis [46,60]. In addition, only a small amount of sample is required to perform the analysis [61,62]. So far, several types of biosensors have been used for the detection of B. cereus. Particularly, DNA-based biosensors have had great success, as they give the possibility for selective identification of different B. cereus strains [63].
Mono or poly-clonal antibodies targeting B. cereus cells can also be used as recognition elements in biosensors as an alternative to DNA probes [64]. Different strategies have been carried out for B. cereus detection using specific antibodies. For instance, Setterington and Alocilja [65] developed a biosensor that associates the immune-magnetic separation of bacterial cells with a cyclic voltammetry detection method (Figure 3). First, magnetic/polyaniline core/shell nanoparticles decorated with a polyclonal anti-B. cereus antibody were incubated with the sample containing B. cereus. Second, a magnetic field separation was applied to collect the bacterial cells bound to magnetic particles and separate them from the background matrix (Figure 3). Finally, the collected magnetic particles were transferred to screen-printed carbon electrodes for voltammetric detection. An LOD as low as 40 CFU/mL was obtained with a pure bacterial culture. Unfortunately, this technique showed a limit in the efficiency of the bacterial collection during the immune-magnetic separation step. In response, the authors recommended that any unknown sample should be both diluted and concentrated by several orders of magnitude before the detection step to prevent false-negative signals.
The disadvantage of antibody-based detection is that commercial anti-B. cereus antibodies are costly and have relatively low affinity. Moreover, their stability and binding capacity can be affected by pH or temperature variations. Bacteriophages and bacteriophage-derived proteins become attractive alternatives to antibodies because of their high host specificity, high binding affinity and marked resistance to chemicals, a variation of pH and temperature. Bacteriophages and their proteins (receptors) can be successfully applied as recognition elements in biosensors for bacterial detection [66]. A B. cereus-specific bacteriophage endolysin cell wall-binding domain (CBD) has been used to develop a highly specific and sensitive surface plasmon resonance biosensor for the detection of B. cereus [67]. This biosensor has permitted us to detect B. cereus at concentrations of 105–108 CFU/ml, and showed an LOD of 102 CFU/ml in pure culture. The low LOD was obtained by coupling a subtractive inhibition assay to the biosensor. For the inhibition assay, bacterial cells were pre-incubated with bacteriophage cell wall-binding domain, thus, only non-bound cells were detected by the sensor, which overcomes the limited sensitivity of the technique due to the direct capture of the bacteria. Using this strategy, a LOD of 103 CFU/ml of B. cereus was obtained for the detection of the bacteria in contaminated cooked rice. Park et al., [68] also highlighted the superior affinity of bacteriophage CBD for B. cereus cells compared to commercial antibodies. Based on this idea, they developed an assay that combined the efficient and specific CBD-conjugated magnetic bacterial separation and the ATP bioluminescence bacterial detection. Using this association, they obtained a LOD as low as 101 CFU/mL. In a recent study, Kong et al., [69] combined CBD and antibody to develop a lateral flow test for the cost-effective and efficient detection of B. cereus. The sensitivity of the strip was 104 CFU/ml and the overall assay time was 20 min. Thanks to the high specificity of CBD toward B. cereus, the test showed no significant cross-reactivity.
Recent works employing different detection strategies and recognition elements to detect B. cereus cells are given in Table 1.
4. Detection of B. cereus Toxins
4.1. Detection of Cereulide and the Emetic B. cereus Strains
Emetic B. cereus strains induce the emetic food intoxication form through the production of a cereulide toxin [73]. The toxin is synthesized by a non-ribosomal cereulide synthetase enzyme encoded by the ces gene [22]. The toxic molecule is a cyclic depsipeptide composed of three repeats of “D-Oxy-Leu-Dala-L-Oxy-Val-L-Val” amino acids sequence (Figure 4A,B). Recently, 18 cereulide variants were reported and differences in toxicity amongst them were highlighted [22].
The efficiency of cereulide production varies between emetic strains with the distinction of no/low, medium and high producers producing respectively <1–50 ng, >50–500 ng and >500–1600 ng/mg of bacterial mass [22,74,75]. According to studies carried out on animals, poisonings appeared after ingestion-doses between 8–10 µg/kg of body weight [75,76]. In a recent FBO, concentrations of cereulide ranging from 2 to 6 µg/g of food provoked profuse-vomiting among high-risk population groups [77]. Furthermore, Decleer et al., [78] have demonstrated that subemetic doses of the toxin are sufficient to alter the mitochondrial activity and affect health. However, strains carrying the ces gene represent a minority, generally 1% or less, of isolates from food or the environment.
Cereulide is highly stable and resistant to treatments used by the food industry (pH, enzyme or high-temperature) [79]. In the past, its quantification in foodstuffs was hampered by the absence of suitable standard methods. Antibody-based strategies were explored but, due to the lack of cereulide immunogen properties and the low reproducibility of such tests, they are not commonly used. The cytotoxicity-based methods measure toxin-induced cell damage. Different types of cells were utilized as cereulide target, such as HEp-2 (Human carcinoma of the larynx) [80], CHO (Chinese Hamster Ovary) cells [81] or rat liver cells [82]. A distinctive bioassay based on boar sperm developed by Anderson et al., [83] aimed to detect emetic B. cereus through the sperm motility inhibition caused by the cereulide exposure. It allowed a qualitative detection of emetic B. cereus directly from the primary culture plates. To evaluate the feasibility in real conditions, the test was evaluated with bacteria isolated from rice, vegetarian dishes, cakes and pasta. The detection limit was found to be 0.3 +/− 1 ng per assay. However, this semi-quantitative bioassay could lack specificity because food matrices may contain molecules with lytic properties.
Chemical methods, like LC-MS (liquid chromatography–mass spectrometry) and Matrix-Assisted Laser Desorption Ionization–Time of Flight (MALDI-TOF), provide low cross-reactivity and high specificity for the quantification of cereulide. These measurements need a chemical standard to quantify the molecule [79]. Commercially available valinomycin was used because of its chemical structure similarity to cereulide. However, quantification of the toxin expressed as valinomycin equivalent has been a contentious point because of the differences between the two molecules in biological and chemical assays. To overcome this drawback, Biesta-Peter et al., [84], tested a synthetic cereulide as an alternative to valinomycin. They showed a faster inhibition of the boar semen mobility using the synthetic cereulide compared to valinomycin. They also demonstrated a similar cytotoxic effect against HEp-2 cells of the natural and synthetic cereulide. This new internal standard associated with LC-MS, was further tested for the quantitative detection of cereulide in food. This methodology has enabled the detection of cereulide toxin in spiked food matrices (cooked rice, chinese noodle and french fries) with a LOD of 4.1 µg/kg of sample. The feasibility of LC-MS/MS method to investigate the presence of cereulide in food leftovers and human samples associated with an outbreak was tested and reported by Delbrassinne et al., [77]. In their report, they combined plate agar counting, PCR strategies and LC-MS/MS to detect emetic B. cereus strain and/or its toxin. A similar methodology was previously used to evaluate the safety of dishes collected in various restaurants [85]. The detected levels of cereulide causing the outbreak were between 3.1 and 4.2 µL/g of foodstuff. Alone, LC-MS/MS, quantitatively detected the toxin in the analyzed samples with a LOD of 1 ng/g which indicates that the method was well adapted for food screening.
Zuberovic Muratovic et al., [79] applied UPLC-ESI-MS/MS (ultra-high-performance liquid chromatography–electrospray-tandem mass spectrometry) in a validation study for quantitative cereulide detection in rice and pasta matrices, using 13C6-cereulide as an internal standard together with the synthetic cereulide standard. The use of natural cereulide as standard eliminates the possible difference that may be generated by the synthetic toxin. The method provided a high specificity and sensitivity as it allows a LOD of 0.1 ng/g.
Recently, MALDI-TOF mass spectroscopy was shown to provide an easy and sensitive approach to differentiate emetic and non-emetic B. cereus (Figure 4C) and to quantify cereulide [86]. To develop the methodology, Ducrest et al., [87] screened for emetic toxin and potential other biomarker peaks until the mass to charge ration (m/z) of 20,000. They detected two characteristic peaks related to sodium and potassium alkali adducts of cereulide at 1175 and 1191 respectively. Thus, these findings accentuated the importance of the buffer used for the extent of the alkali adduct recorded since the prevalence of one rather than the other varied in various buffers. An additional peak at m/z 1205 was identified as cereulide variant. Finally, they demonstrated that using a Laser Desorption/Ionization–Time of Flight (LDI-TOF) instead of MALDI-TOF mass spectrometry, it was possible to ameliorate the LOD from 30 ng/mL to 30 pg/mL in milk-rice and ready-to-eat meals. Overall, chemical methods are demonstrated to detect and identify rapidly and with a high sensibility emetic B. cereus.
The exploitation of chemical methods enabled the establishment of a recent EN-ISO 18465 standard method based on stable isotope mass spectrometry (MS)-based dilution and synthetic cereulide as an intern standard. Although the method allows accurate quantification of cereulide in food, it shows many limitations such as being expensive, reliant on sophisticated instrumentations and experienced persons to performed and interpreted the analyzes.
Molecular methods have also been applied to detect the presence of genes from the ces locus that is responsible for the cereulide synthesis. For instance, Yabutani et al., [73] built two sets of primers as recognition elements in order to specifically detect emetic B. cereus in rice products. Applying RTi-PCR, they were able to obtain a LOD of 104 CFU/g. Yu et al., [89] developed a rapid, sensitive, and specific fluorescent molecular method for emetic B. cereus. The method combined hybridization chain reaction (HCR), an isothermal nucleic acid amplification system, and magnetic beads-based flow cytometric bead assay (MB-based FCBA). The use of specific primers and two fluorescent hairpin probes provided high accuracy in discriminating emetic strains. A target ssDNA generated by asymmetric PCR (aPCR) was isolated onto streptavidin-MBs using a biotin-labeled primer that specifically recognized the target DNA. Then, this system was used to initiate a cascade of hybridization processes between the two fluorescent hairpin probes that allowed the formation of a double-stranded DNA polymer carrying numerous fluorophores. The amplified signal produced by this method was visualized by flow cytometry which had the advantage of measuring the fluorescence intensity of a single MB. This method has an LOD of 7.6 and 9.2 × 102 CFU/mL of emetic B. cereus in pure culture and spiked milk, respectively. The same team modified this detection strategy to microplate format in order to simplify the read-out of the reaction [88]. To detect emetic B. cereus cells in milk, they combined PCR, catalytic hairpin assembly (CHA) and fluorescence-quenching property of graphene oxide (GO) (Figure 5). CHA is an enzyme-free amplification technique that achieves amplification by hybridization and strand-exchange reactions. In CHA an initiator strand triggers the formation of a duplex by a hairpin pair, then the initiator strands is released and is free to trigger subsequent CHA reaction. In the study, a classical PCR was first used to amplify the ces fragment. Then, the target ssDNA products were isolated and mixed with a pair of hairpin probes (H1 and H2). H1 is specific to the ces biomarker therefore they can hybridize together. Binding of the H1 probe to the ssDNA products opened up the probe hairpin structure, which allows the binding of H2 onto it. Hybridization of H1 andH2 induced the release of the ssDNA from the complex. The free ssDNA can then be recycled and used for another ssDNA-H1 reaction. Consequently, multiple H1-H2 duplexes were generated. Because the dsDNA had a low affinity with the GO surface, the H1-H2 duplexes do not interact with the GO and a strong fluorescence can be easily recorded. On the contrary, in the absence of target ssDNA, the two hairpin probes did not hybridize together and therefore strongly bound to the surface of the GO, which caused quenching of the fluorescence. Emetic B. cereus cells were detected in spiked milk samples with a LOD of 6.2 × 101 CFU/mL and 5.9 × 102 CFU/mL in pure culture and in spiked milk, respectively. This assay can be adapted for point-of-care detection.
A summary of the methods used for the detection of cereulide is displayed in Table 2.
4.2. Detection of Diarrheal Toxins
In contrast to the highly stable emetic toxin, enterotoxins produced by B. cereus are destroyed during food processing or when the food is reheated because they are heat-labile and susceptible to protease activity. Because of their instability, enterotoxins are usually detected in foods by molecular methods that evidence their genes [92,94,95]. Alternatively, B. cereus strains have to be cultivated and enterotoxins are identified in culture supernatants.
Three enterotoxins have been associated with the diarrheal syndrome, the non-hemolytic enterotoxin (Nhe), the hemolysin BL (Hbl) and the Cytotoxin K (CytK1, CytK2). Nhe complex is composed of three different proteins, A, B and C, that are encoded by the nheA, nheB and nheC genes, respectively [21]. Nhe is recognized as the major diarrheal toxin of B. cereus because of the strong correlation between cytotoxicity and concentration of Nhe in culture filtrates collected from a large collection of isolates [96]. Additionally, Guinebretière et al., showed that strains with a high production of Nhe are more often associated with food implicated in FBO than with food not associated with any illness [97]. Hbl is composed of a binding protein B encoded by the hblA gene and two lytic components L1 and L2 encoded by hblC and hblD genes, respectively. Hbl was the first enterotoxin identified in B. cereus [98]. Hbl is haemolytic, dermonecrotic and cytotoxic towards enterocytes and various other human cells [99,100,101,102,103,104]. CytK is a β-barrel pore-forming toxin. There are two variants of CytK that share 89% of identity: CytK-1, and CytK2. CytK1 was identified in 1998 from a strain of B. cereus that caused a large FBO in France and that conducted to the death of three elderly people [25]. Although CytK2 is five times less toxic than CytK-1, it seems more frequently associated with strains causing FBO [105].
Several PCR techniques such as mRTi-PCR [91], duplex-PCR [106], PMA-mPCR [88] and PCR coupled with electric chip [107] were used for the detection of one or multiple enterotoxin-encoding genes in food matrices [90,95,108]. Several other authors applied PCR based-strategy to detect enterotoxin-encoded genes together with the ces gene [90,92,109]. These methods used one or several high-affinity set of primers that specifically bind to a selected sequence present inside the gene that encodes for the toxin selected as biomarkers of diarrheal strains. So far in all collections tested, all strains carry at least one component of the nhe operon, while, cytK-1 gene prevalence is low and is only present in strains of the species B. cytotoxicus and cytK-2 gene is carried by 42% of the strains causing FBO [110].
Three immunoassay kits can detect Nhe and Hbl complexes in the culture supernatant of B. cereus: Bacillus diarrheal enterotoxin visual immunoassay (BDE VIATM) kit (3M Tecra), B. cereus enterotoxin reversed passive latex agglutination (BCET-RPLA) kit (Oxoid), and the Duopath® Cereus Enterotoxins (Merck). These three kits are based on lateral flow immunological detection, which allows a fast point-of-need analysis. Figure 6 illustrates the Duopath® Cereus Enterotoxins kit, which can detect both Nhe and Hbl enterotoxins. Ceuppens et al., [111] compared the three detection kits using gastrointestinal samples to verify their suitability for B. cereus enterotoxin diagnosis. They highlighted that BCET-RPLA kit requires a dilution step in order to avoid false-positive outcome but present a low LOD. LOD of Hbl in gastrointestinal samples is similar to the limit described by the manufacturer in contrast with Nhe. Furthermore, they emphasized the inability of the kits to detect all the enterotoxin polymorphisms. Two out of these three kits are no more commercially available.
Other strategies were based on an indirect enzyme immunoassays (EIAs) using high-affinity antibodies. Dietrich et al., [112] compared EIA to detect Nhe in culture supernatants with a sandwich enzyme immunoassay. Moravek et al., [96] used the latter-mentioned technique in comparison with EIAs, cytotoxicity tests and PCR for the enterotoxin detection in B. cereus strains isolated from meat, milk, baby food and clinical environments. The results suggested that Nhe production was high in B. cereus strains causing food poisoning.
A chemical approach based on MALDI-TOF/MS was explored in order to detect the different diarrheal toxins produced by B. cereus. Tsilia et al., [113] applied this approach, and used characteristic m/z spectral positions to identify CytK1 and Nhe proteins in supernatants of different B. cereus cultures. Results were obtained with the BDE VIA kit. MALDI-TOF/MS showed a higher specificity, sensitivity and confidence in enterotoxin detection compared to the lateral flow kit. However, the MS method requires labor-intensive preparations, high investment and maintenance.
A different detection strategy was explored by Gabig-Ciminska et al., [107] who developed an electric chip for the screening of haemolysin producing B. cereus. The chip-recognized hblC and hblA genes. The first step of the test consisted of a sandwich formation between the DNA target, the capturing probes immobilized onto magnetic beads by amino-link, and the specific detection probes labeled with biotin. The formed sandwich was detected by a silicon chip-based potentiometric cell, using extravidin-alkaline phosphatase complex as an enzymatic label. The addition of the enzymatic substrate pAPP enabled for amperometric detection of the reaction product. The biosensor detected specific signals from 20 amol of bacterial DNA in less than 4 h. This low LOD allowed for B. cereus enterotoxin-encoding genes detection in crude cell extracts without preceding nucleic acid amplification [107].
Recently, other electrochemical biosensors were developed to detect diarrheal B. cereus strains. For instance, Izadi et al., [60] developed a gold nanoparticle modified pencil graphene electrode (AuNPs-PGE) for screening diarrheal B. cereus strains in milk and infant formula using electrochemical impedance spectroscopy (EIS). A ssDNA probe targeting nheA gene was selected as a recognition element and immobilized on the electrode surface. For this purpose, the probe was modified by a thiol group (HS-(CH2)6 at the 5′ end to enable its self-assembling onto the gold nanoparticles and electro-deposited over a pencil graphite electrode, as illustrated in Figure 7. Hybridization of the target DNA against the immobilized ss-DNA increased the charge transfer resistance of the biosensor enabling the detection of B. cereus. A LOD of 100 CFU/mL was achieved using genomic DNA extracted from bacteria from spiked infant formula.
4.3. Detection of Other B. cereus Pathogenic Factors
The detection of Nhe and Hbl enterotoxins provides an indication of the toxicity of a strain but are not sufficient to totally discriminate between pathogenic and non-pathogenic strain potentials [97]. Indeed, several studies have shown that the Nhe production by hazardous strains is variable and that non-pathogenic strains can also produce it in large quantities [90,114].
In addition to Nhe and Hbl toxins, B. cereus produces several compounds (degradation enzymes, cytotoxic factors, hemolysins, proteases, cell-surface proteins) that might also contribute to virulence [105,115]. For instance, InhA1 and NprA are secreted metalloproteases, which interfere with the host immune defenses and play a role in the virulence of B. cereus [116,117,118,119]. Other enterotoxins, such as enterotoxin T (BcET) and enterotoxin FM (EntFM) were reported to be involved in foodborne illness [120,121]. The pore-forming Hemolysin II (HlyII) toxin induces macrophage apoptosis and allows B. cereus to bypass the host immune defenses [121,122]. The importance of HlyII in the pathogenicity of B. cereus [122] has been demonstrated in several models, invertebrates and mammals [123].
Although the role of these proteins are not fully established, several authors applied molecular techniques to detect their genes together with the other toxin-encoded genes in FBO or clinical strains in a multiplex detection format [90,92,109,124]. For instance, [95] characterized B. cereus toxigenic strains by means of PCR technique in several food samples from local restaurants and supermarkets, using cytK, bceT and genes encoring Hbl and Nhe complexes as biomarkers. They showed that almost 70% of the isolates held the bceT gene. The high presence of the enterotoxin encoded by the bceT in the foodstuffs suggested its importance in B. cereus virulence. The search for the hlyII gene in a large B. cereus collection including FBO-related strains and human clinical samples, revealed that the hlyII gene is present in approximately a quarter of B. cereus strains [125,126] and those which carry this gene are described as pathogens [126].
A short list of the main strategies explored to detect B. cereus toxins is displayed in Table 3.
However, to the best of our knowledge, common markers to all pathogenic strains have not been described. For example, some genes considered as virulence factors such as nhe, pi-plc and entFM are present in all B. cereus strains, pathogenic or not [23,90,97,131,132], while well-known virulence factors such as hlyII, cytk-1 and ces are not characteristic of all pathogenic strains. It is even apparent that many strains causing infections do not possess any of them [23,126].
Taken together, the virulence potential of B. cereus is likely multifactorial with a combination of known and maybe unknown factors been responsible for the various pathologies associated with B. cereus infections. This implies that only multiplex detection of B. cereus genes should be applied to determine the pathogenic potential of a particular strain. Currently, there are no advanced detection methods developed to detect these potential factors at the protein level. In addition, these factors are not yet searched in foodstuff or in clinical sample on a routine basis. Thereafter, innovative multiplexed technologies need to be developed for the detection of pathogenic toxins of B. cereus, in order to provide a diagnostic tool to improve both clinical diagnosis and food safety.
5. Conclusions
Bacillus cereus is an important cause of foodborne pathogenesis because it contaminates numerous foods including infant formula, infant rice cereal, milk and dried milk products, rice (including cooked rice), eggs, meat, and spices. Pathogenic B. cereus strains cause food poisoning or even severe human infectious diseases with an infective dose as low as 103 CFU/g of food. Although B. cereus is currently searched only in infant formula, it is recommended as a suitable microbiological safety indicator for food products. The traditional detection methods that enumerate B. cereus colonies on selective agars are labor, cost-intensive and time-consuming (from 2 to 7 days for confirmation). Furthermore, they cannot detect injured or VBNC cells. Subsequently, they are not suitable for the needs of the food industry. Molecular methods, immunoassays and chemical methods have been developed to identify B. cereus strains that produce toxins such as cereulide, cytotoxin K, Hbl and Nhe. To reveal the prevalence and characteristics of B. cereus in various foods PCR-based methods targeting one or several toxin genes like hblA, hblC, hblD, nheA, nheB, nheC, cytK, entFM, bceT, hlyII, and ces are usually applied. In the past decade, electrochemical, optical and colorimetric biosensors exhibiting a rapid, low cost and high sensitivity have been developed for specific detection of B. cereus in food products. Considering that biosensors may exhibit high reproducibility and stability and can be miniaturized to a portable multiplex format, they can constitute a suitable tool to reduce risk of B. cereus contamination. Regular screening of foodstuffs for B. cereus will enhance the perception about contamination and provide a theoretical basis for developing effective measures to reduce product recalls and substantial economic loss. At the hospital level, an accurate diagnosis will help improving patient care and health.
Presented biosensors based on various recognition elements such as DNA probe, antibodies, aptamers, or phage possess advantages and disadvantages, often trading efficiency and robustness in exchange for specificity, affordability or limit of detection. For instance, antibodies are used in biosensors and assays for the detection of B. cereus cells or their toxins. Although it is usually easy to perform such tests because they may be performed without biomarker purification or special sample pretreatment, antibodies may show cross-reactivity, which potentially reduces their specificity. Peptides and aptamers represent attractive alternatives to expensive antibodies for a specific bacterial targeting since they offer not only good sensitivity and specificity but also high stability and chemical resistance. Especially aptamers selected by the cell-SELEX technology are very promising tools for bacterial cells recognition [133,134]. However, up to now, aptamer-based biosensors have been developed only for B. cereus spores detection [6]. Finally, DNA based biosensors make possible to differentiate specific B. cereus strains or to detect their toxin-producing genes. To enable efficient detection, DNA probes should be designed carefully to combine maximal specificity and minimal non-specific interaction, and to avoid any formation of the secondary structure of the probe, which will prevent its access to targets. Although biosensors offer great improvements compared to classical culture-based methods, there are many challenges to overcome, especially those involving their stability. We strongly believe that an interdisciplinary approach involving analytical chemistry, microbiology, materials science, biochemistry, and molecular biology will allow for the development of proposed systems at the proof-of-concept stage to reach the market in the future.
Author Contributions
All authors contributed equally. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by the European Union’s Horizon 2020 research and innovation the programme under the Marie Skłodowska–Curie grant agreement No 872662 (IPANEMA), the European Union’s Horizon 2020 research and innovation programme under the European EJP grant agreement No 773830 (Toxdetect), and by the University Paris-Saclay through the Poc in labs 2019 grant agreement No 00003469 (OSCAR).
Acknowledgments
Figure 1. We thank Thierry Meylheuc for its expertise and the MIMA2 platform for access to electron microscopy equipment (MIMA2, INRAE, 2018. Microscopy and Imaging Facility for Microbes, Animals and Foods, https://0-doi-org.brum.beds.ac.uk/10.15454/1.5572348210007727E12). Figure 2. Reprinted by permission from ELSEVIER license 4792010480337, Asymmetric polymerase chain assay combined with propidium monoazide treatment and unmodified gold nanoparticles for colorimetric detection of viable emetic Bacillus cereus in milk, Sens. Actuat. B: Chem., Fan Li, Fulai Li, Guotai Yang, Zoraida P. Aguilar, Weihua Lai, Hengyi Xu, 2018; Figure 4C Reprinted by permission from ELSEVIER, Identification of cereulide producing Bacillus cereus by MALDI-TOF MS, Food Microbiol., Sebastian Ulrich, Christoph Gottschalk, Richard Dietrich, Erwin Märtlbauer, Manfred Gareis, 2019, License Number 4804080940109. Figure 5: Adapted with permission from Catalytic hairpin assembly combined with graphene oxide for the detection of emetic Bacillus cereus in milk, Zhongxu Zhan, Bei Yu, Hui Li, Leina Yan, Zoraida P. Aguilar, Hengyi Xu. Journal of Dairy Science. 2019 ELSEVIER License Number 4804081321892. Figure 7: Reprinted by permission from ELSEVIER license 4796940392012. Fabrication of an electrochemical DNA-based biosensor for Bacillus cereus detection in milk and infant formula. Zahra Izadi, Mahmoud Sheikh-Zeinoddin, Ali A. Ensafi, Sabihe Soleimanian-Zad. Biosens. Bioelectron., 2016.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bacillus Cereus Factsheet 2016 FINAL ACCESSIBLE. Available online: https://www.fsai.ie/WorkArea/DownloadAsset.aspx?id=10919 (accessed on 7 May 2020).
- EFSA Panel on Biological Hazards (BIOHAZ). Risks for public health related to the presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in foodstuffs. EFSA J. 2016, 14, e04524. [Google Scholar]
- Tewari, A.; Abdullah, S. Bacillus cereus food poisoning: International and Indian perspective. J. Food Sci. Technol. 2015, 52, 2500–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.-L.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.-A.; Ramarao, N. Bacillus cereus-induced food-borne outbreaks in France, 2007 to 2014: Epidemiology and genetic characterisation. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Glasset, B.; Herbin, S.; Granier, S.A.; Cavalié, L.; Lafeuille, E.; Guérin, C.; Ruimy, R.; Casagrande-Magne, F.; Levast, M.; Chautemps, N. Bacillus cereus, a serious cause of nosocomial infections: Epidemiologic and genetic survey. PLoS ONE 2018, 13, e0194346. [Google Scholar] [CrossRef] [PubMed]
- Vidic, J.; Chaix, C.; Manzano, M.; Heyndrickx, M. Food Sensing: Detection of Bacillus cereus Spores in Dairy Products. Biosensors 2020, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Du, J.; Lai, Q.; Zeng, R.; Ye, D.; Xu, J.; Shao, Z. Proposal of nine novel species of the Bacillus cereus group. Int. J. Syst. Evol. Microbiol. 2017, 67, 2499–2508. [Google Scholar] [CrossRef]
- Liu, Y.; Lai, Q.; Göker, M.; Meier-Kolthoff, J.P.; Wang, M.; Sun, Y.; Wang, L.; Shao, Z. Genomic insights into the taxonomic status of the Bacillus cereus group. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Okinaka, R.T.; Keim, P. The phylogeny of Bacillus cereus sensu lato. In The Bacterial Spore: From Molecules to Systems; ASM: Washington, DC, USA, 2016; pp. 237–251. [Google Scholar]
- Guinebretière, M.H.; Thompson, F.L.; Sorokin, A.; Normand, P.; Dawyndt, P.; Ehling-Schulz, M.; Svensson, B.; Sanchis, V.; Nguyen-The, C.; Heyndrickx, M. Ecological diversification in the Bacillus cereus group. Environ. Microbiol. 2008, 10, 851–865. [Google Scholar] [CrossRef]
- Guinebretičre, M.-H.; Velge, P.; Couvert, O.; Carlin, F.; Debuyser, M.-L. Ability of Bacillus cereus group strains to cause food poisoning varies according to phylogenetic affiliation (groups I to VII) rather than species affiliation. J. Clin. Microbiol. 2010, 48, 3388–3391. [Google Scholar] [CrossRef] [Green Version]
- Altayar, M.; Sutherland, A. Bacillus cereus is common in the environment but emetic toxin producing isolates are rare. J. Appl. Microbiol. 2006, 100, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus group: Bacillus species with pathogenic potential. In Gram-Positive Pathogens; ASM: Washington, DC, USA, 2019; pp. 875–902. [Google Scholar]
- Jensen, G.; Hansen, B.; Eilenberg, J.; Mahillon, J. The hidden lifestyles of Bacillus cereus and relatives. Environ. Microbiol. 2003, 5, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, S.; Boon, N.; Uyttendaele, M. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS Microbiol. Ecol. 2013, 84, 433–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [Green Version]
- Koletzko, B.; Baker, S.; Cleghorn, G.; Neto, U.F.; Gopalan, S.; Hernell, O.; Hock, Q.S.; Jirapinyo, P.; Lonnerdal, B.; Pencharz, P. Global standard for the composition of infant formula: Recommendations of an ESPGHAN coordinated international expert group. J. Pediatr. Gastr. Nutr. 2005, 41, 584–599. [Google Scholar] [CrossRef] [Green Version]
- Six, S.; De Buyser, M.; Vignaud, M.; Dao, T.; Messio, S.; Pairaud, S.; Hennekinne, J.; Pihier, N. Bacillus cereus food poisoning outbreaks: Strain characterization results, 2006–2010. BEH—Bulletin Épidémiologique Hebdomadaire 2012, 2012, 45–49. [Google Scholar]
- Riol, C.D.; Dietrich, R.; Märtlbauer, E.; Jessberger, N. Consumed foodstuffs have a crucial impact on the toxic activity of enteropathogenic Bacillus cereus. Front. Microbiol. 2018, 9, 1946. [Google Scholar] [CrossRef] [Green Version]
- Jeßberger, N.; Krey, V.M.; Rademacher, C.; Böhm, M.-E.; Mohr, A.-K.; Ehling-Schulz, M.; Scherer, S.; Märtlbauer, E. From genome to toxicity: A combinatory approach highlights the complexity of enterotoxin production in Bacillus cereus. Front. Microbiol. 2015, 6, 560. [Google Scholar]
- Marxen, S.; Stark, T.D.; Frenzel, E.; Rütschle, A.; Lücking, G.; Pürstinger, G.; Pohl, E.E.; Scherer, S.; Ehling-Schulz, M.; Hofmann, T. Chemodiversity of cereulide, the emetic toxin of Bacillus cereus. Anal. Bioanal. Chem. 2015, 407, 2439–2453. [Google Scholar] [CrossRef]
- Kim, J.B.; Kim, J.M.; Cho, S.H.; Oh, H.S.; Choi, N.J.; Oh, D.H. Toxin genes profiles and toxin production ability of Bacillus cereus isolated from clinical and food samples. J. Food Sci. 2011, 76, T25–T29. [Google Scholar] [CrossRef]
- Berthold-Pluta, A.; Pluta, A.; Garbowska, M. The effect of selected factors on the survival of Bacillus cereus in the human gastrointestinal tract. Microb. Pathog. 2015, 82, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.; De Buyser, M.L.; Granum, P.E. A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Agata, N.; Mori, M.; Ohta, M.; Suwan, S.; Ohtani, I.; Isobe, M. A novel dodecadepsipeptide, cereulide, isolated from Bacillus cereus causes vacuole formation in HEp-2 cells. FEMS Microbiol. Lett. 1994, 121, 31–34. [Google Scholar] [PubMed] [Green Version]
- Mahler, H.; Pasi, A.; Kramer, J.M.; Schulte, P.; Scoging, A.C.; Bär, W.; Krähenbühl, S. Fulminant liver failure in association with the emetic toxin of Bacillus cereus. N. Engl. J. Med. 1997, 336, 1142–1148. [Google Scholar] [CrossRef]
- Dierick, K.; Van Coillie, E.; Swiecicka, I.; Meyfroidt, G.; Devlieger, H.; Meulemans, A.; Hoedemaekers, G.; Fourie, L.; Heyndrickx, M.; Mahillon, J. Fatal family outbreak of Bacillus cereus-associated food poisoning. J. Clin. Microbiol. 2005, 43, 4277–4279. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, M.; Denayer, S.; Botteldoorn, N.; Delbrassinne, L.; Veys, J.; Waegenaere, J.; Sirtaine, N.; Driesen, R.B.; Sipido, K.R.; Mahillon, J. Sudden death of a young adult associated with Bacillus cereus food poisoning. J. Clin. Microbiol. 2011, 49, 4379–4381. [Google Scholar] [CrossRef] [Green Version]
- Tschiedel, E.; Rath, P.M.; Steinmann, J.; Becker, H.; Dietrich, R.; Paul, A.; Felderhoff-Müser, U.; Dohna-Schwake, C. Lifesaving liver transplantation for multi-organ failure caused by Bacillus cereus food poisoning. Pediatr. Trans. 2015, 19, E11–E14. [Google Scholar] [CrossRef]
- Hoffmaster, A.R.; Hill, K.K.; Gee, J.E.; Marston, C.K.; De, B.K.; Popovic, T.; Sue, D.; Wilkins, P.P.; Avashia, S.B.; Drumgoole, R. Characterization of Bacillus cereus isolates associated with fatal pneumonias: Strains are closely related to Bacillus anthracis and harbor B. anthracis virulence genes. J. Clin. Microbiol. 2006, 44, 3352–3360. [Google Scholar] [CrossRef] [Green Version]
- Ramarao, N.; Belotti, L.; Deboscker, S.; Ennahar-Vuillemin, M.; De Launay, J.; Lavigne, T.; Koebel, C.; Escande, B.; Guinebretiere, M.H. Two unrelated episodes of Bacillus cereus bacteremia in a neonatal intensive care unit. Am. J. Infect. Control 2014, 42, 694–695. [Google Scholar] [CrossRef]
- Decousser, J.; Ramarao, N.; Duport, C.; Dorval, M.; Bourgeois-Nicolaos, N.; Guinebretiere, M.H.; Razafimahefa, H.; Doucet-Populaire, F. Bacillus cereus and severe intestinal infections in preterm neonates: Putative role of the pooled breast milk. Am. J. Infect. Control 2013, 41, 918–921. [Google Scholar] [CrossRef]
- Lotte, R.; Herisse, A.L.; Berrouane, Y.; Lotte, L.; Casagrande, F.; Landraud, L.; Herbin, S.; Ramarao, N.; Boyer, L.; Ruimy, R. Virulence Analysis of Bacillus cereus Isolated after Death of Preterm Neonates, Nice, France, 2013. Emerg. Infect. Dis. 2017, 23, 845–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacteriological Analytical Manual; Association of Official Analytical Chemists: Washington, DC, USA, 1978; Volume 1.
- Li, F.; Li, F.; Yang, G.; Aguilar, Z.P.; Lai, W.; Xu, H. Asymmetric polymerase chain assay combined with propidium monoazide treatment and unmodified gold nanoparticles for colorimetric detection of viable emetic Bacillus cereus in milk. Sens. Actuat. B Chem. 2018, 255, 1455–1461. [Google Scholar] [CrossRef]
- Sacchi, C.T.; Whitney, A.M.; Mayer, L.W.; Morey, R.; Steigerwalt, A.; Boras, A.; Weyant, R.S.; Popovic, T. Sequencing of 16S rRNA gene: A rapid tool for identification of Bacillus anthracis. Emerg. Infect. Dis. 2002, 8, 1117. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.; Griffiths, H.; Saltmarsh, M.; Peters, A.; Fielding, L. Profiling Bacillus cereus populations in a traditional style, hot-drinks vending machine and vended hot chocolate drink using polymerase chain reaction (PCR) and random amplified polymorphic DNA (RAPD) techniques. Food Control 2012, 27, 127–131. [Google Scholar] [CrossRef]
- Fernández-No, I.; Guarddon, M.; Böhme, K.; Cepeda, A.; Calo-Mata, P.; Barros-Velázquez, J. Detection and quantification of spoilage and pathogenic Bacillus cereus, Bacillus subtilis and Bacillus licheniformis by real-time PCR. Food Microbiol. 2011, 28, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Shou, Y.; Yoshida, T.M.; Marrone, B.L. Differentiation of Bacillus anthracis, B. cereus, and B. thuringiensis by using pulsed-field gel electrophoresis. Appl. Environ. Microbiol. 2007, 73, 3446–3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, M.; Giusto, C.; Iacumin, L.; Cantoni, C.; Comi, G. Molecular methods to evaluate biodiversity in Bacillus cereus and Bacillus thuringiensis strains from different origins. Food Microbiol. 2009, 26, 259–264. [Google Scholar] [CrossRef]
- Garbeva, P.; Van Veen, J.; Van Elsas, J. Predominant Bacillus spp. in agricultural soil under different management regimes detected via PCR-DGGE. Microbial Ecol. 2003, 45, 302–316. [Google Scholar] [CrossRef]
- Guinebretiere, M.-H.; Nguyen-The, C. Sources of Bacillus cereus contamination in a pasteurized zucchini puree processing line, differentiated by two PCR-based methods. FEMS Microbiol. Ecol. 2003, 43, 207–215. [Google Scholar] [CrossRef]
- Noguera, P.A.; Ibarra, J.E. Detection of new cry genes of Bacillus thuringiensis by use of a novel PCR primer system. Appl. Environ. Microbiol. 2010, 76, 6150–6155. [Google Scholar] [CrossRef] [Green Version]
- Antolinos, V.; Fernández, P.S.; Ros-Chumillas, M.; Periago, P.M.; Weiss, J. Development of a high-resolution melting–based approach for efficient differentiation among Bacillus cereus group isolates. Foodborne Pathog. Dis. 2012, 9, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Qu, Q.; Li, L.; Ran, X.; Gui, J.; Wang, Q.; Cui, X.; Jiang, C. Electrochemical DNA Biosensor Based on Magnetite/Multiwalled Carbon Nanotubes/Chitosan Nanocomposite for Bacillus cereus Detection of Potential Marker for Gold Prospecting. Electroanalysis 2018, 30, 910–920. [Google Scholar] [CrossRef]
- Kang, X.; Pang, G.; Chen, Q.; Liang, X. Fabrication of Bacillus cereus electrochemical immunosensor based on double-layer gold nanoparticles and chitosan. Sens. Actuat. B Chem. 2013, 177, 1010–1016. [Google Scholar] [CrossRef]
- Nassi, A.; Guillon, F.-X.; Amar, A.; Hainque, B.; Amriche, S.; Maugé, D.; Markova, E.; Tsé, C.; Bigey, P.; Lazerges, M. Electrochemical DNA-biosensors based on long-range electron transfer: Optimization of the amperometric detection in the femtomolar range using two-electrode setup and ultramicroelectrode. Electrochim. Acta 2016, 209, 269–277. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, J.; Ma, W.; Zheng, W. PCR microfluidic devices for DNA amplification. Biotechnol. Adv. 2006, 24, 243–284. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Z.; Corstjens, P.L.; Mauk, M.G.; Bau, H.H. A disposable microfluidic cassette for DNA amplification and detection. Lab Chip 2006, 6, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Mauk, M.G.; Chen, D.; Liu, C.; Bau, H.H. A large volume, portable, real-time PCR reactor. Lab Chip 2010, 10, 3170–3177. [Google Scholar] [CrossRef]
- Vidic, J.; Manzano, M.; Chang, C.-M.; Jaffrezic-Renault, N. Advanced biosensors for detection of pathogens related to livestock and poultry. Vet. Res. 2017, 48, 11. [Google Scholar] [CrossRef] [Green Version]
- Vidic, J.; Vizzini, P.; Manzano, M.; Kavanaugh, D.; Ramarao, N.; Zivkovic, M.; Radonic, V.; Knezevic, N.; Giouroudi, I.; Gadjanski, I. Point-of-need DNA testing for detection of foodborne pathogenic bacteria. Sensors 2019, 19, 1100. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Liu, Z.; Yan, S.; Yin, F.; Feng, X.; Liu, B.-F. Identifying multiple bacterial pathogens by loop-mediated isothermal amplification on a rotate & react slipchip. Sens. Actuat. B Chem. 2016, 228, 491–499. [Google Scholar]
- Vizzini, P.; Braidot, M.; Vidic, J.; Manzano, M. Electrochemical and Optical Biosensors for the Detection of Campylobacter and Listeria: An Update Look. Micromachines 2019, 10, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, M.; Viezzi, S.; Mazerat, S.; Marks, R.S.; Vidic, J. Rapid and label-free electrochemical DNA biosensor for detecting hepatitis A virus. Biosens. Bioelectron. 2018, 100, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Kubičárová, T.; Fojta, M.; Vidic, J.; Tomschik, M.; Suznjevic, D.; Paleček, E. Voltammetric and chronopotentiometric measurements with nucleic acid-modified mercury film on a glassy carbon electrode. Electroanalysis 2000, 12, 1390–1396. [Google Scholar] [CrossRef]
- Miodek, A.; Sauriat-Dorizon, H.; Chevalier, C.; Delmas, B.; Vidic, J.; Korri-Youssoufi, H. Direct electrochemical detection of PB1-F2 protein of influenza A virus in infected cells. Biosens. Bioelectron. 2014, 59, 6–13. [Google Scholar] [CrossRef]
- Miodek, A.; Vidic, J.; Sauriat-Dorizon, H.; Richard, C.-A.; Le Goffic, R.; Korri-Youssoufi, H.; Chevalier, C. Electrochemical detection of the oligomerization of PB1-F2 influenza A virus protein in infected cells. Anal. Chem. 2014, 86, 9098–9105. [Google Scholar] [CrossRef]
- Izadi, Z.; Sheikh-Zeinoddin, M.; Ensafi, A.A.; Soleimanian-Zad, S. Fabrication of an electrochemical DNA-based biosensor for Bacillus cereus detection in milk and infant formula. Biosens. Bioelectron. 2016, 80, 582–589. [Google Scholar] [CrossRef]
- Pal, S.; Ying, W.; Alocilja, E.C.; Downes, F.P. Sensitivity and specificity performance of a direct-charge transfer biosensor for detecting Bacillus cereus in selected food matrices. Biosyst. Eng. 2008, 99, 461–468. [Google Scholar] [CrossRef]
- Kotsiri, Z.; Vantarakis, A.; Rizzotto, F.; Kavanaugh, D.; Ramarao, N.; Vidic, J. Sensitive Detection of E. coli in Artificial Seawater by Aptamer-Coated Magnetic Beads and Direct PCR. Appl. Sci. 2019, 9, 5392. [Google Scholar] [CrossRef] [Green Version]
- Velusamy, V.; Arshak, K.; Korostynska, O.; Oliwa, K.; Adley, C. Conducting polymer based DNA biosensor for the detection of the Bacillus cereus group species. Proc. SPIE 2009. [Google Scholar] [CrossRef]
- Skottrup, P.D.; Nicolaisen, M.; Justesen, A.F. Towards on-site pathogen detection using antibody-based sensors. Biosens. Bioelectron. 2008, 24, 339–348. [Google Scholar] [CrossRef]
- Setterington, E.B.; Alocilja, E.C. Electrochemical biosensor for rapid and sensitive detection of magnetically extracted bacterial pathogens. Biosensors 2012, 2, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tawil, N.; Sacher, E.; Mandeville, R.; Meunier, M. Bacteriophages: Biosensing tools for multi-drug resistant pathogens. Analyst 2014, 139, 1224–1236. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Sim, J.; Kang, T.; Nguyen, H.H.; Park, H.K.; Chung, B.H.; Ryu, S. A novel and highly specific phage endolysin cell wall binding domain for detection of Bacillus cereus. Eur. Biophys. J. 2015, 44, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Kong, M.; Lee, J.-H.; Ryu, S.; Park, S. Detection of Bacillus Cereus Using Bioluminescence Assay with Cell Wall-Binding Domain Conjugated Magnetic Nanoparticles. BioChip J. 2018, 12, 287–293. [Google Scholar] [CrossRef]
- Kong, M.; Shin, J.H.; Heu, S.; Park, J.-K.; Ryu, S. Lateral flow assay-based bacterial detection using engineered cell wall binding domains of a phage endolysin. Biosens. Bioelectron. 2017, 96, 173–177. [Google Scholar] [CrossRef]
- Pal, S.; Alocilja, E.C.; Downes, F.P. Nanowire labeled direct-charge transfer biosensor for detecting Bacillus species. Biosens. Bioelectron. 2007, 22, 2329–2336. [Google Scholar] [CrossRef]
- Chu, P.-T.; Hsieh, M.-F.; Yin, S.-Y.; Wen, H.-W. Development of a rapid and sensitive immunomagnetic-bead based assay for detecting Bacillus cereus in milk. Eur. Food Res. Technol. 2009, 229, 73. [Google Scholar] [CrossRef] [Green Version]
- Spieker, E.; Lieberzeit, P.A. Molecular imprinting studies for developing qcm-sensors for Bacillus cereus. Proc. Eng. 2016, 168, 561–564. [Google Scholar] [CrossRef]
- Yabutani, M.; Agata, N.; Ohta, M. A new rapid and sensitive detection method for cereulide-producing Bacillus cereus using a cycleave real-time PCR. Lett. Appl. Microbiol. 2009, 48, 698–704. [Google Scholar]
- Stark, T.; Marxen, S.; Rütschle, A.; Lücking, G.; Scherer, S.; Ehling-Schulz, M.; Hofmann, T. Mass spectrometric profiling of Bacillus cereus strains and quantitation of the emetic toxin cereulide by means of stable isotope dilution analysis and HEp-2 bioassay. Anal. Bioanal. Chem. 2013, 405, 191–201. [Google Scholar] [CrossRef]
- Rouzeau-Szynalski, K.; Stollewerk, K.; Messelhaeusser, U.; Ehling-Schulz, M. Why be serious about emetic Bacillus cereus: Cereulide production and industrial challenges. Food Microbiol. 2020, 85, 103279. [Google Scholar] [CrossRef] [PubMed]
- Marxen, S.; Stark, T.D.; Rütschle, A.; Lücking, G.; Frenzel, E.; Scherer, S.; Ehling-Schulz, M.; Hofmann, T. Multiparametric Quantitation of the Bacillus cereus Toxins Cereulide and Isocereulides A–G in Foods. J. Agric. Food Chem. 2015, 63, 8307–8313. [Google Scholar] [CrossRef] [PubMed]
- Delbrassinne, L.; Botteldoorn, N.; Andjelkovic, M.; Dierick, K.; Denayer, S. An emetic Bacillus cereus outbreak in a kindergarten: Detection and quantification of critical levels of cereulide toxin. Foodborne Pathog. Dis. 2015, 12, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Decleer, M.; Jovanovic, J.; Vakula, A.; Udovicki, B.; Agoua, R.-S.E.; Madder, A.; De Saeger, S.; Rajkovic, A. Oxygen consumption rate analysis of mitochondrial dysfunction caused by Bacillus cereus cereulide in Caco-2 and HepG2 cells. Toxins 2018, 10, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuberovic Muratovic, A.; Tröger, R.; Granelli, K.; Hellenäs, K.-E. Quantitative analysis of cereulide toxin from Bacillus cereus in rice and pasta using synthetic cereulide standard and 13C6-cereulide standard—A short validation study. Toxins 2014, 6, 3326–3335. [Google Scholar] [CrossRef]
- Hughes, S.; Bartholomew, B.; Hardy, J.; Kramer, J. Potential application of a HEp-2 cell assay in the investigation of Bacillus cereus emetic-syndrome food poisoning. FEMS Microbiol. Lett. 1988, 52, 7–11. [Google Scholar] [CrossRef]
- Beattie, S.A.; Williams, A. Detection of toxigenic strains of Bacillus cereus and other Bacillus spp. with an improved cytotoxicity assay. Lett. Appl. Microbiol. 1999, 28, 221–225. [Google Scholar] [CrossRef]
- Kawamura-Sato, K.; Hirama, Y.; Agata, N.; Ito, H.; Torii, K.; Takeno, A.; Hasegawa, T.; Shimomura, Y.; Ohta, M. Quantitative analysis of cereulide, an emetic toxin of Bacillus cereus, by using rat liver mitochondria. Microbiol. Immunol. 2005, 49, 25–30. [Google Scholar] [CrossRef]
- Andersson, M.A.; Jääskeläinen, E.L.; Shaheen, R.; Pirhonen, T.; Wijnands, L.M.; Salkinoja-Salonen, M.S. Sperm bioassay for rapid detection of cereulide-producing Bacillus cereus in food and related environments. Int. J. Food Microbiol. 2004, 94, 175–183. [Google Scholar] [CrossRef]
- Biesta-Peters, E.G.; Reij, M.W.; Blaauw, R.H.; Rajkovic, A.; Ehling-Schulz, M.; Abee, T. Quantification of the emetic toxin cereulide in food products by liquid chromatography-mass spectrometry using synthetic cereulide as a standard. Appl. Environ. Microbiol. 2010, 76, 7466–7472. [Google Scholar] [CrossRef] [Green Version]
- Delbrassinne, L.; Andjelkovic, M.; Dierick, K.; Denayer, S.; Mahillon, J.; Van Loco, J. Prevalence and levels of Bacillus cereus emetic toxin in rice dishes randomly collected from restaurants and comparison with the levels measured in a recent foodborne outbreak. Foodborne Pathog. Dis. 2012, 9, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, S.; Gottschalk, C.; Dietrich, R.; Märtlbauer, E.; Gareis, M. Identification of cereulide producing Bacillus cereus by MALDI-TOF MS. Food Microbiol. 2019, 82, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Ducrest, P.J.; Pfammatter, S.; Stephan, D.; Vogel, G.; Thibault, P.; Schnyder, B. Rapid detection of Bacillus ionophore cereulide in food products. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Feng, L.; Xu, H.; Liu, C.; Shah, N.P.; Wei, H. Detection of viable enterotoxin-producing Bacillus cereus and analysis of toxigenicity from ready-to-eat foods and infant formula milk powder by multiplex PCR. J. Dairy Sci. 2016, 99, 1047–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Li, F.; Zhao, T.; Li, F.; Zhou, B.; Xu, H. Hybridization chain reaction-based flow cytometric bead sensor for the detection of emetic Bacillus cereus in milk. Sens. Actuat. B Chem. 2018, 256, 624–631. [Google Scholar] [CrossRef]
- Wehrle, E.; Moravek, M.; Dietrich, R.; Bürk, C.; Didier, A.; Märtlbauer, E. Comparison of multiplex PCR, enzyme immunoassay and cell culture methods for the detection of enterotoxinogenic Bacillus cereus. J. Microbiol. Methods 2009, 78, 265–270. [Google Scholar] [CrossRef]
- Wehrle, E.; Didier, A.; Moravek, M.; Dietrich, R.; Märtlbauer, E. Detection of Bacillus cereus with enteropathogenic potential by multiplex real-time PCR based on SYBR green I. Mol. Cell. Probes 2010, 24, 124–130. [Google Scholar] [CrossRef]
- Forghani, F.; Langaee, T.; Eskandari, M.; Seo, K.-H.; Chung, M.-J.; Oh, D.-H. Rapid detection of viable Bacillus cereus emetic and enterotoxic strains in food by coupling propidium monoazide and multiplex PCR (PMA-mPCR). Food Control 2015, 55, 151–157. [Google Scholar] [CrossRef]
- In’t Veld, P.; Van der Laak, L.; Van Zon, M.; Biesta-Peters, E. Elaboration and validation of the method for the quantification of the emetic toxin of Bacillus cereus as described in EN-ISO 18465-Microbiology of the food chain–Quantitative determination of emetic toxin (cereulide) using LC-MS/MS. Int. J. Food Microbiol. 2019, 288, 91–96. [Google Scholar] [CrossRef]
- Tallent, S.M.; Hait, J.M.; Knolhoff, A.M.; Bennett, R.W.; Hammack, T.S.; Croley, T.R. Rapid Testing of Food Matrices for Bacillus cereus Enterotoxins. J. Food Saf. 2017, 37. [Google Scholar] [CrossRef]
- Batchoun, R.; Al-Sha’er, A.I.; Khabour, O.F. Molecular characterization of Bacillus cereus toxigenic strains isolated from different food matrices in Jordan. Foodborne Pathog. Dis. 2011, 8, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Moravek, M.; Dietrich, R.; Buerk, C.; Broussolle, V.; Guinebretière, M.-H.; Granum, P.E.; Nguyen-the, C.; Märtlbauer, E. Determination of the toxic potential of Bacillus cereus isolates by quantitative enterotoxin analyses. FEMS Microbiol. Lett. 2006, 257, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinebretière, M.-H.; Broussolle, V. Enterotoxigenic profiles of food-poisoning and food-borne Bacillus cereus strains. J. Clin. Microbiol. 2002, 40, 3053–3056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beecher, D.J.; Wong, A. Identification of hemolysin BL-producing Bacillus cereus isolates by a discontinuous hemolytic pattern in blood agar. Appl. Environ. Microbiol. 1994, 60, 1646–1651. [Google Scholar] [CrossRef] [Green Version]
- Thompson, N.; Ketterhagen, M.; Bergdoll, M.; Schantz, E. Isolation and some properties of an enterotoxin produced by Bacillus cereus. Infect. Immun. 1984, 43, 887–894. [Google Scholar] [CrossRef] [Green Version]
- Beecher, D.J.; MacMillan, J.D. A novel bicomponent hemolysin from Bacillus cereus. Infect. Immun. 1990, 58, 2220–2227. [Google Scholar] [CrossRef] [Green Version]
- Beecher, D.J.; Macmillan, J. Characterization of the components of hemolysin BL from Bacillus cereus. Infect. Immun. 1991, 59, 1778–1784. [Google Scholar] [CrossRef] [Green Version]
- Beecher, D.J.; Wong, A. Improved purification and characterization of hemolysin BL, a hemolytic dermonecrotic vascular permeability factor from Bacillus cereus. Infect. Immun. 1994, 62, 980–986. [Google Scholar] [CrossRef] [Green Version]
- Beecher, D.J.; Schoeni, J.L.; Wong, A. Enterotoxic activity of hemolysin BL from Bacillus cereus. Infect. Immun. 1995, 63, 4423–4428. [Google Scholar] [CrossRef] [Green Version]
- Beecher, D.J.; Wong, A.C. Tripartite haemolysin BL: Isolation and characterization of two distinct homologous sets of components from a single Bacillus cereus isolate. Microbiology 2000, 146, 1371–1380. [Google Scholar] [CrossRef] [Green Version]
- Ramarao, N.; Sanchis, V. The pore-forming haemolysins of Bacillus cereus: A review. Toxins 2013, 5, 1119–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otlewska, A.; Oltuszak-Walczak, E.; Walczak, P. Differentiation of strains from the Bacillus cereus group by RFLP-PFGE genomic fingerprinting. Electrophoresis 2013, 34, 3023–3028. [Google Scholar] [PubMed]
- Gabig-Ciminska, M.; Andresen, H.; Albers, J.; Hintsche, R.; Enfors, S.-O. Identification of pathogenic microbial cells and spores by electrochemical detection on a biochip. Microb. Cell Factories 2004, 3, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh-Lakha, S.; Leon-Velarde, C.G.; Chen, S.; Lee, S.; Shannon, K.; Fabri, M.; Downing, G.; Keown, B. A study to assess the numbers and prevalence of Bacillus cereus and its toxins in pasteurized fluid milk. J. Food Prot. 2017, 80, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Tallent, S.; Hait, J.; Bennett, R. Analysis of Bacillus cereus toxicity using PCR, ELISA and a lateral flow device. J. Appl. Microbiol. 2015, 118, 1068–1075. [Google Scholar] [CrossRef]
- Guinebretiere, M.H.; Auger, S.; Galleron, N.; Contzen, M.; De Sarrau, B.; De Buyser, M.L.; Lamberet, G.; Fagerlund, A.; Granum, P.E.; Lereclus, D. Bacillus cytotoxicus sp. nov. is a new thermotolerant species of the Bacillus cereus group occasionally associated with food poisoning. Int. J. Syst. Evol. Microbiol. 2012. [Google Scholar] [CrossRef]
- Ceuppens, S.; Rajkovic, A.; Hamelink, S.; Van de Wiele, T.; Boon, N.; Uyttendaele, M. Enterotoxin production by Bacillus cereus under gastrointestinal conditions and their immunological detection by commercially available kits. Foodborne Pathog. Dis. 2012, 9, 1130–1136. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, R.; Moravek, M.; Bürk, C.; Granum, P.E.; Märtlbauer, E. Production and characterization of antibodies against each of the three subunits of the Bacillus cereus nonhemolytic enterotoxin complex. Appl. Environ. Microbiol. 2005, 71, 8214–8220. [Google Scholar] [CrossRef] [Green Version]
- Tsilia, V.; Devreese, B.; De Baenst, I.; Mesuere, B.; Rajkovic, A.; Uyttendaele, M.; Van de Wiele, T.; Heyndrickx, M. Application of MALDI-TOF mass spectrometry for the detection of enterotoxins produced by pathogenic strains of the Bacillus cereus group. Anal. Bioanal. Chem. 2012, 404, 1691–1702. [Google Scholar] [CrossRef]
- Martínez-Blanch, J.; Sánchez, G.; Garay, E.; Aznar, R. Development of a real-time PCR assay for detection and quantification of enterotoxigenic members of Bacillus cereus group in food samples. Int. J. Food Microbiol. 2009, 135, 15–21. [Google Scholar] [CrossRef]
- Guillemet, E.; Lerééc, A.; Royer, C.; Tran, S.; Barbosa, I.; Sansonetti, P.; Lereclus, D.; Ramarao, N. The bacterial repair protein Mfd confers resistance to the host nitric-oxide response. Sci. Rep. 2016, 6, 29349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlton, S.; Moir, A.; Baillie, L.; Moir, A. Characterization of the exosporium of Bacillus cereus. J. Appl. Microbiol. 1999, 87, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Ramarao, N.; Lereclus, D. The InhA1 metalloprotease allows spores of the B. cereus group to escape macrophages. Cell. Microbiol. 2005, 7, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Haydar, A.; Tran, S.-L.; Guillemet, E.; Darrigo, C.; Perchat, S.; Lereclus, D.; Coquet, L.; Jouenne, T.; Ramarao, N. InhA1-mediated cleavage of the metalloprotease NprA allows Bacillus cereus to escape from macrophages. Front. Microbiol. 2018, 9, 1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemet, E.; Cadot, C.; Tran, S.L.; Guinebretiere, M.H.; Lereclus, D.; Ramarao, N. The InhA metalloproteases of Bacillus cereus contribute concomitantly to virulence. J. Bacteriol. 2010, 192, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinagawa, K. Analytical methods for Bacillus cereus and other Bacillus species. Int. J. Food Microbiol. 1990, 10, 125–141. [Google Scholar] [CrossRef]
- Tran, S.-L.; Guillemet, E.; Gohar, M.; Lereclus, D.; Ramarao, N. CwpFM (EntFM) is a Bacillus cereus potential cell wall peptidase implicated in adhesion, biofilm formation, and virulence. J. Bacteriol. 2010, 192, 2638–2642. [Google Scholar] [CrossRef] [Green Version]
- Tran, S.L.; Guillemet, E.; Ngo-Camus, M.; Clybouw, C.; Puhar, A.; Moris, A.; Gohar, M.; Lereclus, D.; Ramarao, N. Haemolysin II is a Bacillus cereus virulence factor that induces apoptosis of macrophages. Cell. Microbiol. 2011, 13, 92–108. [Google Scholar] [CrossRef]
- Tran, S.-L.; Ramarao, N. Bacillus cereus immune escape: A journey within macrophages. FEMS Microbiol. Lett. 2013, 347, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chon, J.-W.; Yim, J.-H.; Kim, H.-S.; Kim, D.-H.; Kim, H.; Oh, D.-H.; Kim, S.-K.; Seo, K.-H. Quantitative prevalence and toxin gene profile of Bacillus cereus from ready-to-eat vegetables in South Korea. Foodborne Pathog. Dis. 2015, 12, 795–799. [Google Scholar] [CrossRef]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the cytK family of genes in Bacillus cereus. Microbiology 2004, 150, 2689–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadot, C.; Tran, S.-L.; Vignaud, M.-L.; De Buyser, M.-L.; Kolstø, A.-B.; Brisabois, A.; Lereclus, D.; Guinebretière, M.-H.; Ramarao, N. InhA1, NprA, and HlyII as candidates for markers to differentiate pathogenic from nonpathogenic Bacillus cereus strains. J. Clin. Microbiol. 2010, 48, 1358–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, B.M.; Hendriksen, N.B. Detection of Enterotoxic Bacillus cereus and Bacillus thuringiensis Strains by PCR Analysis. Appl. Environ. Microbiol. 2001, 67, 185–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabig-Ciminska, M.; Liu, Y.; Enfors, S.-O. Gene-based identification of bacterial colonies with an electric chip. Anal. Biochem. 2005, 345, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Elsholz, B.; Enfors, S.-O.; Gabig-Ciminska, M. Confirmative electric DNA array-based test for food poisoning Bacillus cereus. J. Microbiol. Methods 2007, 70, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Krause, N.; Moravek, M.; Dietrich, R.; Wehrle, E.; Slaghuis, J.; Märtlbauer, E. Performance characteristics of the Duopath® Cereus Enterotoxins assay for rapid detection of enterotoxinogenic Bacillus cereus strains. Int. J. Food Microbiol. 2010, 144, 322–326. [Google Scholar] [CrossRef]
- Martinez-Blanch, J.F.; Sanchez, G.; Garay, E.; Aznar, R. Evaluation of a real-time PCR assay for the detection and quantification of Bacillus cereus group spores in food. J. Food Prot. 2010, 73, 1480–1485. [Google Scholar] [CrossRef]
- Zhao, S.; Chen, J.; Fei, P.; Feng, H.; Wang, Y.; Ali, M.A.; Li, S.; Jing, H.; Yang, W. Prevalence, molecular characterization, and antibiotic susceptibility of Bacillus cereus isolated from dairy products in China. J. Dairy Sci. 2020, 103, 3994–4001. [Google Scholar] [CrossRef]
- Kaur, H.; Bruno, J.G.; Kumar, A.; Sharma, T.K. Aptamers in the therapeutics and diagnostics pipelines. Theranostics 2018, 8, 4016. [Google Scholar] [CrossRef]
- Kaur, H. Recent developments in cell-SELEX technology for aptamer selection. Biochim. Biophys. Acta Gener. Subj. 2018, 1862, 2323–2329. [Google Scholar] [CrossRef]
Figure 1.
Scanning electron microscopy image of B. cereus cells.

Figure 2.
Schematic illustration of the colorimetric method based on PMA-asPCR (a) and visualization of gold nanoparticles for the detection of viable emetic B. cereus. Transmission electron microscope images correspond to (b) AuNPs alone, (c) AuNPs treated with 1 pmol ssDNA and 25 mM NaCl, and (d) AuNPs upon the addition of 25 mM NaCl without ssDNA. Insets show the respective images. (Reprinted with Permission from [36]).
Figure 2.
Schematic illustration of the colorimetric method based on PMA-asPCR (a) and visualization of gold nanoparticles for the detection of viable emetic B. cereus. Transmission electron microscope images correspond to (b) AuNPs alone, (c) AuNPs treated with 1 pmol ssDNA and 25 mM NaCl, and (d) AuNPs upon the addition of 25 mM NaCl without ssDNA. Insets show the respective images. (Reprinted with Permission from [36]).

Figure 3.
Immunomagnetic separation from sample matrix, magnetic alignment on SPCE surface and electrochemical detection of B. cereus cells (from [65]).
Figure 3.
Immunomagnetic separation from sample matrix, magnetic alignment on SPCE surface and electrochemical detection of B. cereus cells (from [65]).

Figure 4.
The 2D (A) and 3D (B) chemical structure of cereulide from ChemSpider (Royal Chemical Society). (C) Mass spectra of B. cereus strains/isolates, emetic strain in red, non-emetic strains in green (from [86]).
Figure 4.
The 2D (A) and 3D (B) chemical structure of cereulide from ChemSpider (Royal Chemical Society). (C) Mass spectra of B. cereus strains/isolates, emetic strain in red, non-emetic strains in green (from [86]).

Figure 5.
The principle of catalytic hairpin assembly-graphene oxide assay to detect emetic B. cereus in the presence of the target ssDNA (A) and in the negative control (B), (Adapted with permission from [88]).
Figure 5.
The principle of catalytic hairpin assembly-graphene oxide assay to detect emetic B. cereus in the presence of the target ssDNA (A) and in the negative control (B), (Adapted with permission from [88]).

Figure 6.
Example of Duopath® lateral flow kit that detected the absence of enterotoxin (A), the presence of Nhe (B) and the presence of both Nhe and Hbl enterotoxins (C).
Figure 6.
Example of Duopath® lateral flow kit that detected the absence of enterotoxin (A), the presence of Nhe (B) and the presence of both Nhe and Hbl enterotoxins (C).

Figure 7.
Schematic presentation of the immobilization of the ssDNA on a pencil graphite electrode modified with nano-gold (A) and its hybridization with B. cereus genomic DNA (B) (Reprinted with Permission from [60]).
Figure 7.
Schematic presentation of the immobilization of the ssDNA on a pencil graphite electrode modified with nano-gold (A) and its hybridization with B. cereus genomic DNA (B) (Reprinted with Permission from [60]).


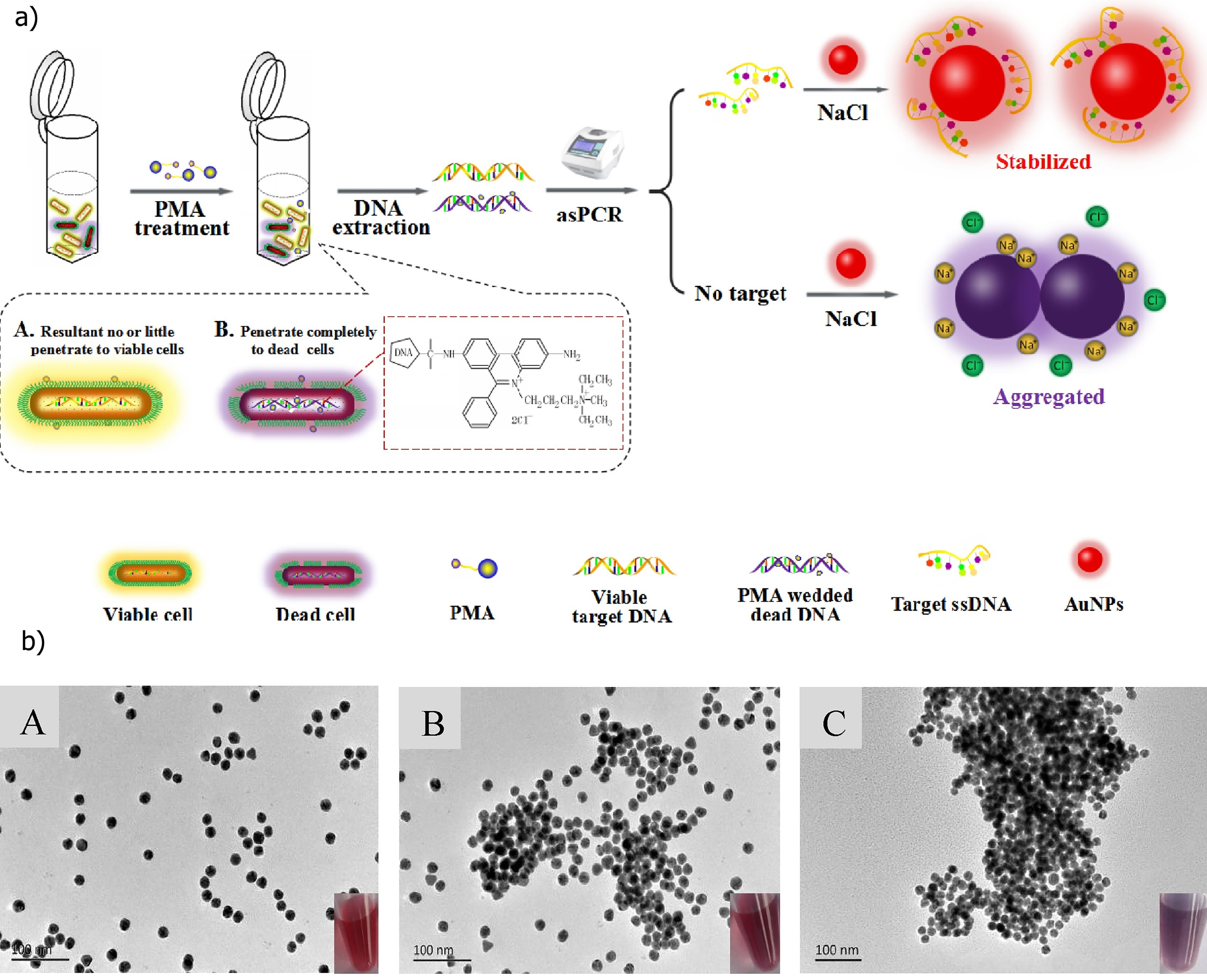{kind=link}
{kind=link}
{kind=link}
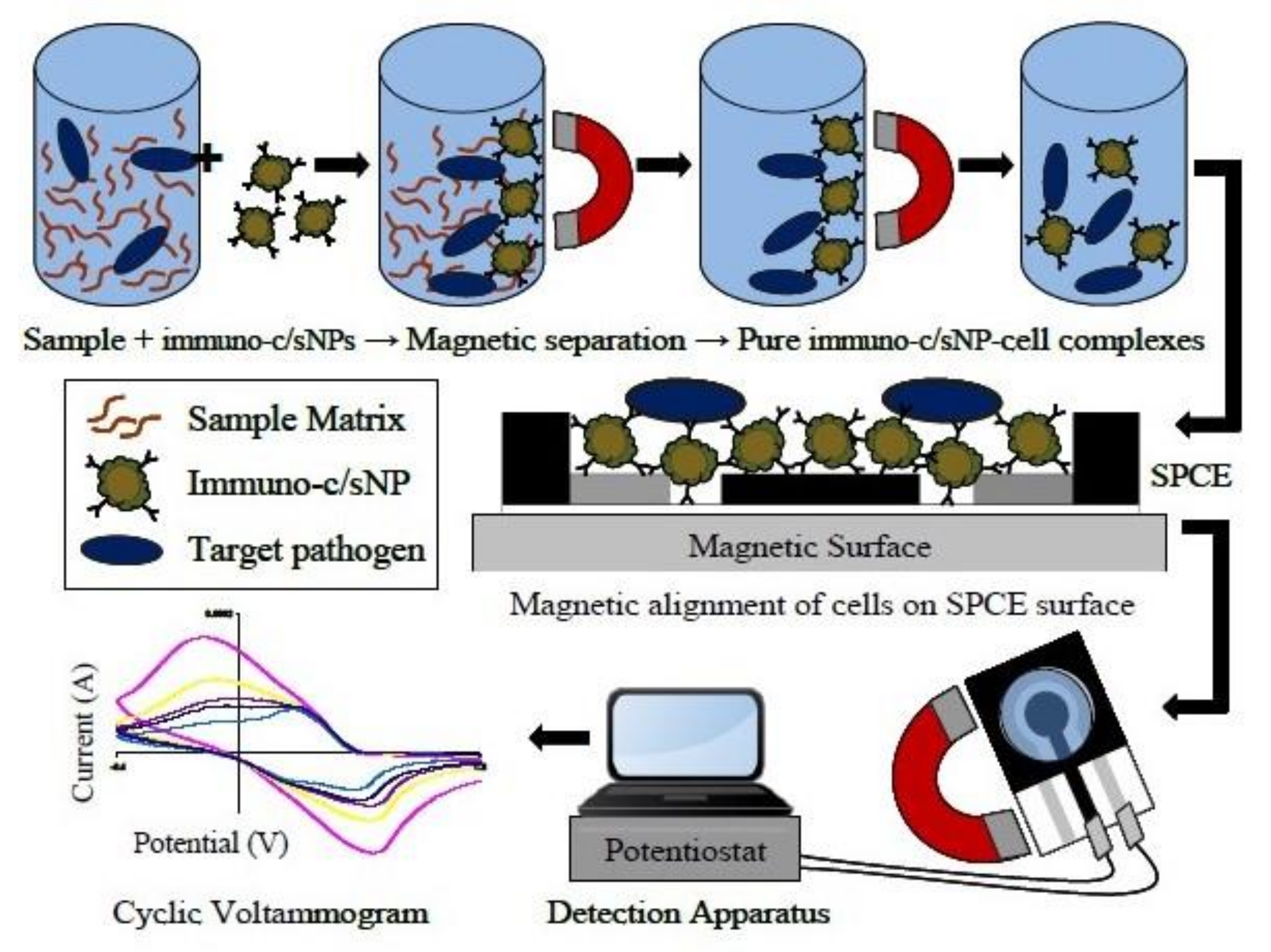{kind=link}
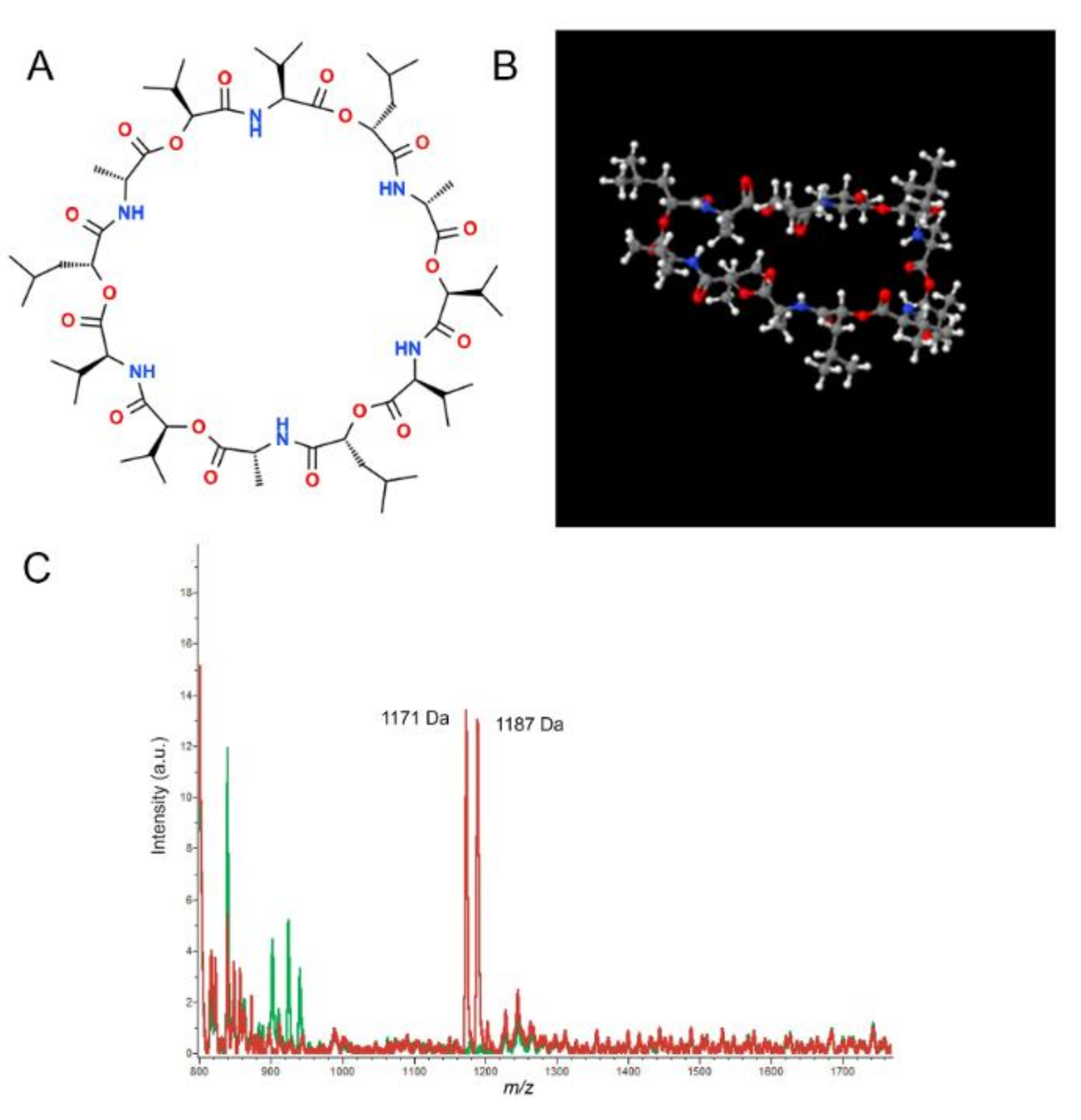{kind=link}
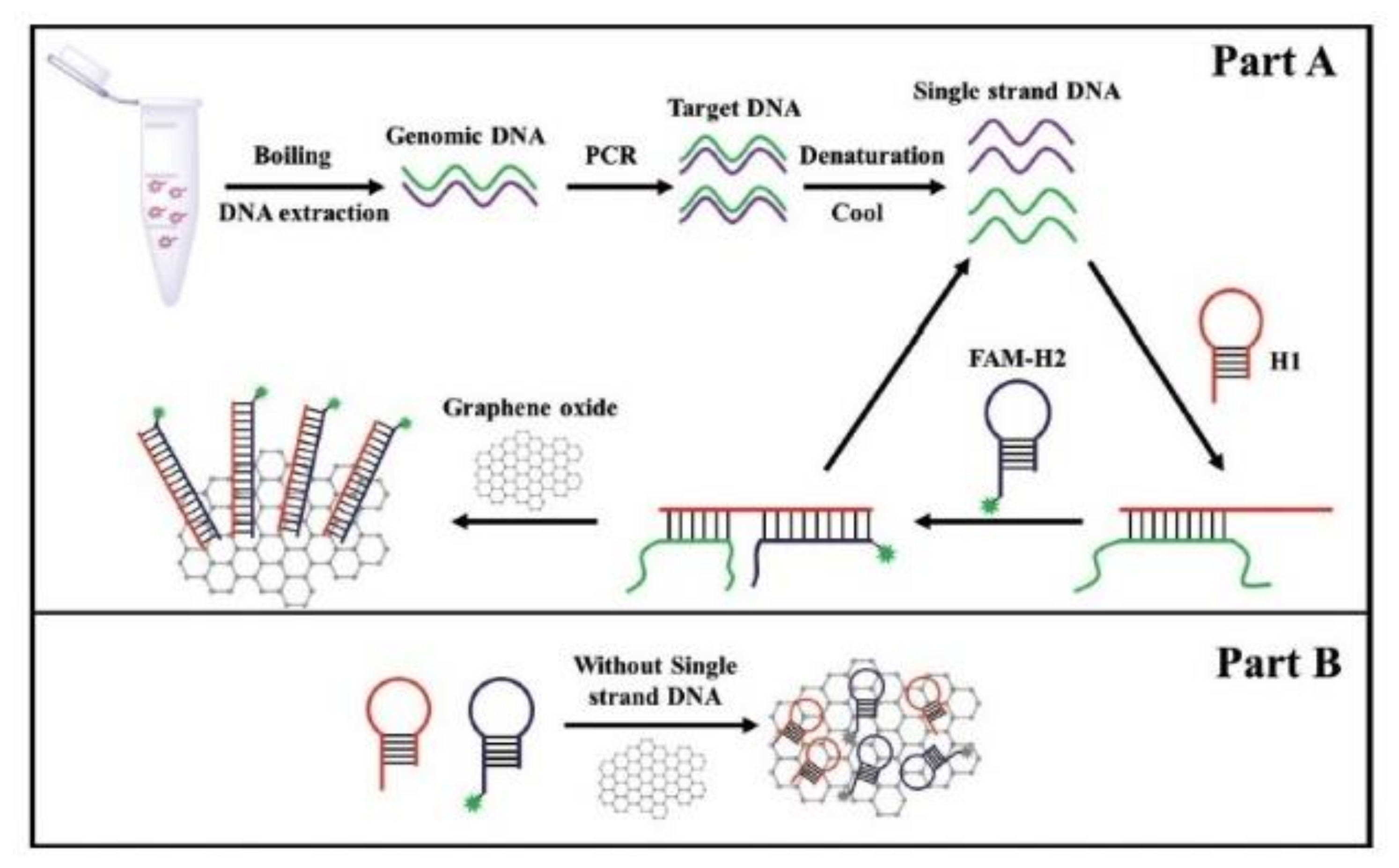{kind=link}
{kind=link}
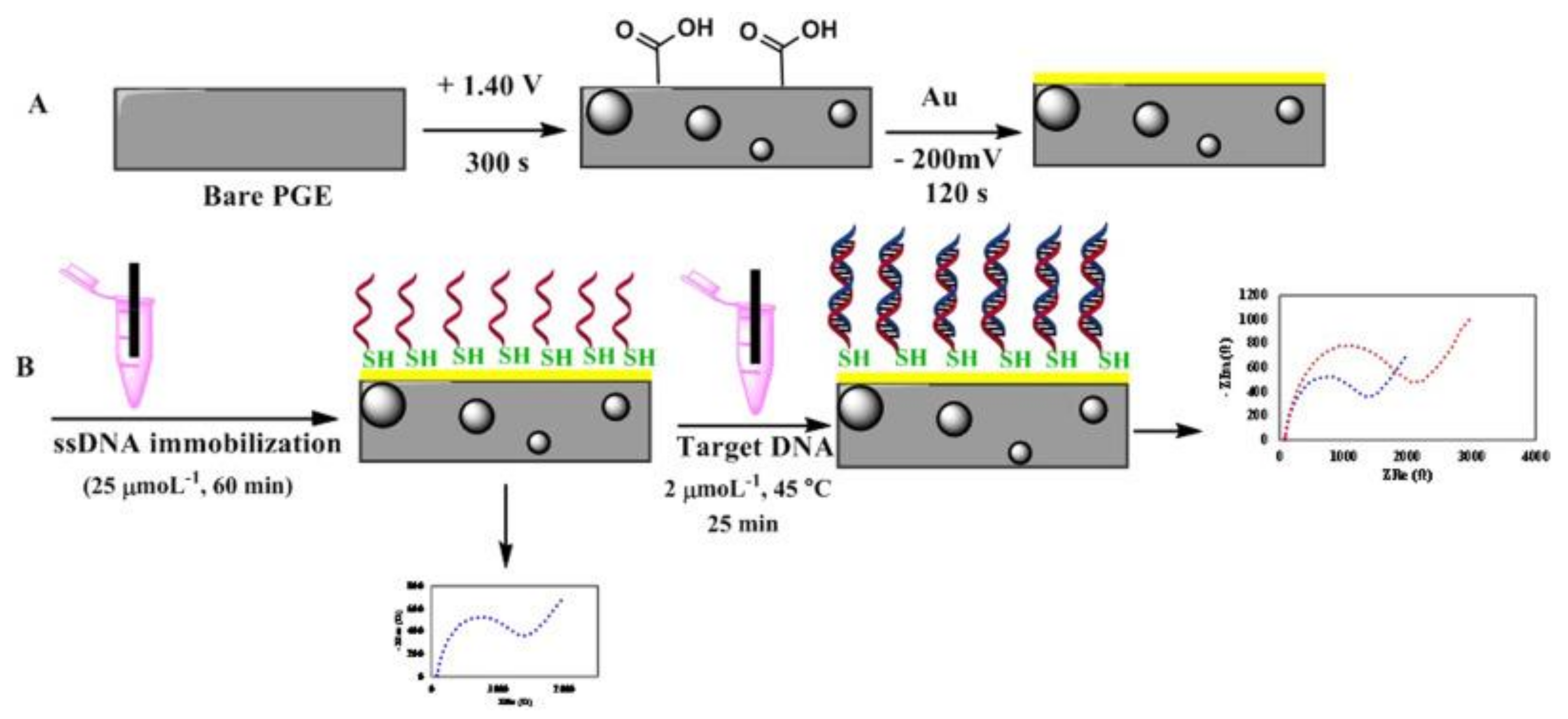{kind=link}
Table 1.
Detection methods for B. cereus vegetative cells.
| Recognition Element | Biomarker | Method | Limit of Detection (LOD) | Matrice | Reference |
|---|---|---|---|---|---|
| Polyclonal antibodies | Whole-cell | Direct-charge transfer biosensor | 101 to 102 CFU/mL | Pure culture | [70] |
| Polyclonal antibodies | Whole-cell | Direct-charge transfer biosensor | 35–88 CFU/mL | Inoculated sprout, strawberries, lettuce, tomatoes, fried rice and corn. | [61] |
| DNA probe | DNA | PCR-TTGE; RAPD-PCR; rep-PCR | Food, patients and pesticides | [41] | |
| DNA probe | motB gene | Electrochemical biosensor | Pure culture | [63] | |
| Antibodies | Whole-cell | IMLN* Magnetic sensor | 10 CFU/mL | Milk | [71] |
| Primer-probe | 16S rRNA | RTi-PCR | 16.5 CFU/mL | Pasteurized food | [39] |
| Primer | gyrB gene | PCR and RAPD PCR | Vended hot chocolate powder and hot-drinks vending machine | [38] | |
| Polyclonal antibodies | Whole-cell | Cyclic voltammetry with IMS* | 40 CFU/mL | Pure culture | [65] |
| Phage endolysin CBD | Whole-cell | Surface Plasmon Resonance (SPR) | 102 CFU/mL | Pure culture | [67] |
| Primer | Cereulide sinthetase gene (cesB) | PMA-asPCR-AuNPs colorimetric assay | 9.2 × 101 CFU/mL 3.4 × 102 CFU/mL | Pure culture Milk | [36] |
| Antibody or Bacteriophage CBD | Wholecell | ATP bioluminescence assay | 101 CFU /mL | Pure culture | [68] |
| Probe | DNA | Electrochemical DNA biosensor | 2.0 × 10−15 M | Pure culture | [46] |
| Molecularly Imprinted Polymer | Whole-cell | Quartz crystal microbalance | 107 CFU/mL | Water | [72] |
* IMLN = Immunomagnetic beads-immunopolisomal nanovesicles; IMS = Immuno magnetic separation.
Table 2.
Detection methods for cereulide and emetic B. cereus vegetative cells.
| Recognition Element | Biomarker | Method | Matrice | Reference |
|---|---|---|---|---|
| Primer | ces gene | mPCR | Pure culture | [90] |
| Primer and probe | ces gene | RTi-PCR | Pure culture | [73] |
| Cereulide; valinomycin | m/z 1170.7 m/z 1128.5 | LC-MS | Cooked rice; Chinese noodle dish | [84] |
| Primer | ces gene | mRTi-PCR | Spiked baby food | [91] |
| Cereulide; cereulide-C13 | m/z 1170.7 m/z 1176.7 | UPLC-ESI*-MS/MS | Rice; pasta | [79] |
| Primer | ces gene | PMA*-mPCR | Spiked baby cereal; pasteurized milk rice | [92] |
| Mass/Charge | Cereulide | LC-MS/MS | Spiked rice; cream pastry; mini pancakes; infant formula | [93] |
| Mass/Charge | Cereulide; cereulide variant | *MALDI-TOF/MS | Rice; milk; ready-to-eat meals | [87] |
| Mass/Charge | Cereulide | *MALDI-TOF/MS | Pure culture | [86] |
| Primer and hairpin probe | ces gene | Fluorescence assay combined with PCR, CHA* and GO* | Pure culture Milk | [88] |
Note: * ESI= Electrospray; PMA = Propidium monoazide; MALDI-TOF = Matrix-assisted laser desorption/ionization-time of flight; CHA = catalytic hairpin assembly; GO = graphene oxide.
Table 3.
Detection method for B. cereus enterotoxins and other pathogenic factors.
| Recognition Element | Biomarker | Method | Matrice | Reference |
|---|---|---|---|---|
| Primer | hblA,hblD,hblC/nheA, nheB,nheC/bceT | PCR | Pure culture | [127] |
| Antibodies | nheA,nheB,nheC | EIAs | Infant formula and dried milk products | [112] |
| Primer and probe | hblC | Electrochemical biosensor | Pure culture | [128] |
| Antibodies, primer | HBL-L2/nheA, nheB,nheC | EIAs, sandwich enzyme immunoassay, cytotoxicity test, PCR | Remnant connected to food-borne outbreak | [96] |
| Primer and probe | hblA,hblD,hblC/nheA, nheB,nheC/cytK/ces | Electric DNA array | Pure culture | [129] |
| DNA probe | Pc-plc gene | RTi-PCR | Artificially contaminated liquid eggs and infant formula | [114] |
| Antibodies, primer | hblA,hblD,hblC/nheA, nheB,nheC/cytK1/ces | Immunoassay, mPCR, cytotoxicity test | Pure culture | [90] |
| Monoclonal antibodies | HBL-L2/nheB | Duopath® kit | Artificially contaminated baby food, rice | [130] |
| Primer | hblA,hblD,hblC/nheA, nheB,nheC/bceT/ cytK1/ces | PCR, BCET-RPLA | Rice, yogurt, pasta, cake | [95] |
| Antibodies | NheA, NheB/Hbl-L2 | BDE VIATM; BCET-RPLA; Duopath® kit | Lasagna, human faeces, potatoes | [111] |
| Mass/charge | Cytk1 and NheA | MALDI-TOF/MS | Pure culture | [113] |
| Primer | hblD/nheA/entFM/cytK/ces | PMA-mPCR | Pure culture | [92] |
| Probe | nheA | Electrochemical DNA-based biosensor | Milk and infant formula | [60] |
| Antibodies, primer | nheA/hblC/entFM/cytK | TECRA, mPCR | Pasteurized milk | [108] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ramarao, N.; Tran, S.-L.; Marin, M.; Vidic, J. Advanced Methods for Detection of Bacillus cereus and Its Pathogenic Factors. Sensors 2020, 20, 2667. https://0-doi-org.brum.beds.ac.uk/10.3390/s20092667
AMA Style
Ramarao N, Tran S-L, Marin M, Vidic J. Advanced Methods for Detection of Bacillus cereus and Its Pathogenic Factors. Sensors. 2020; 20(9):2667. https://0-doi-org.brum.beds.ac.uk/10.3390/s20092667
Chicago/Turabian StyleRamarao, Nalini, Seav-Ly Tran, Marco Marin, and Jasmina Vidic. 2020. "Advanced Methods for Detection of Bacillus cereus and Its Pathogenic Factors" Sensors 20, no. 9: 2667. https://0-doi-org.brum.beds.ac.uk/10.3390/s20092667
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.