Rational Design of a User-Friendly Aptamer/Peptide-Based Device for the Detection of Staphylococcus aureus

 , , ,
, , ,  , , , , , ,
, , , , , ,  , , , ,
, , , , 
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biorecognition Element | Detection Method | Detection Limit | Ref. |
|---|---|---|---|
| PEI-GA modified antibody | amperometric | 10 CFU/mL | [20] |
| lytic phage | surface plasmon resonance-based sensor | 104 CFU/mL | [21] |
| monoclonal antibody | optical fiber | 104 CFU/mL | [22] |
| hybridizing S. aureus DNA | electrochemical (multiwalled carbon nanotubes-chitosan-bismuth) | 3.17 × 10−14 M | [24] |
| hybridizing S. aureus ssDNA | chitosan–Co3O4 nanorod–graphene | 4.3 × 10−13 M | [58] |
| DNA aptamer | potentiometric (single-walled carbon nanotubes) | 8 × 102 CFU/mL | [10] |
| DNA aptamer | graphene interdigitated gold electrode | 41 CFU/mL | [30] |
| biotynilated DNA aptamers | electrochemical (silver nanoparticles) | 1.0 CFU/mL | [31] |
| DNA aptamer | fluorescence (labeled aptamer) | 102 CFU/mL | [32] |
| aptamer-conjugated gold nanoparticles | resonance light-scattering–detection system | single cells | [36] |
2. Materials and Methods
2.1. In Silico Design
2.2. Molecular Dynamics
2.3. Peptide Synthesis and Labeling
2.4. Circular Dichroism
2.5. Fluorescence Analysis
2.6. Nanostructured Zirconia Substrates
2.7. Aptamers Microarrays
2.8. Fluorescence Confocal Microscopy
2.9. Bacterial Culture
2.10. Detection of Bacteria-Aptamer Interaction
2.11. Bacterial Displacement
3. Results and Discussion
3.1. Aptamer Selection for In Silico Design of Interacting Peptide
3.2. Peptide Scaffold Selection
3.3. In Silico Design and Synthesis of λ-Cro Mutant Specific for Aptamer Binding
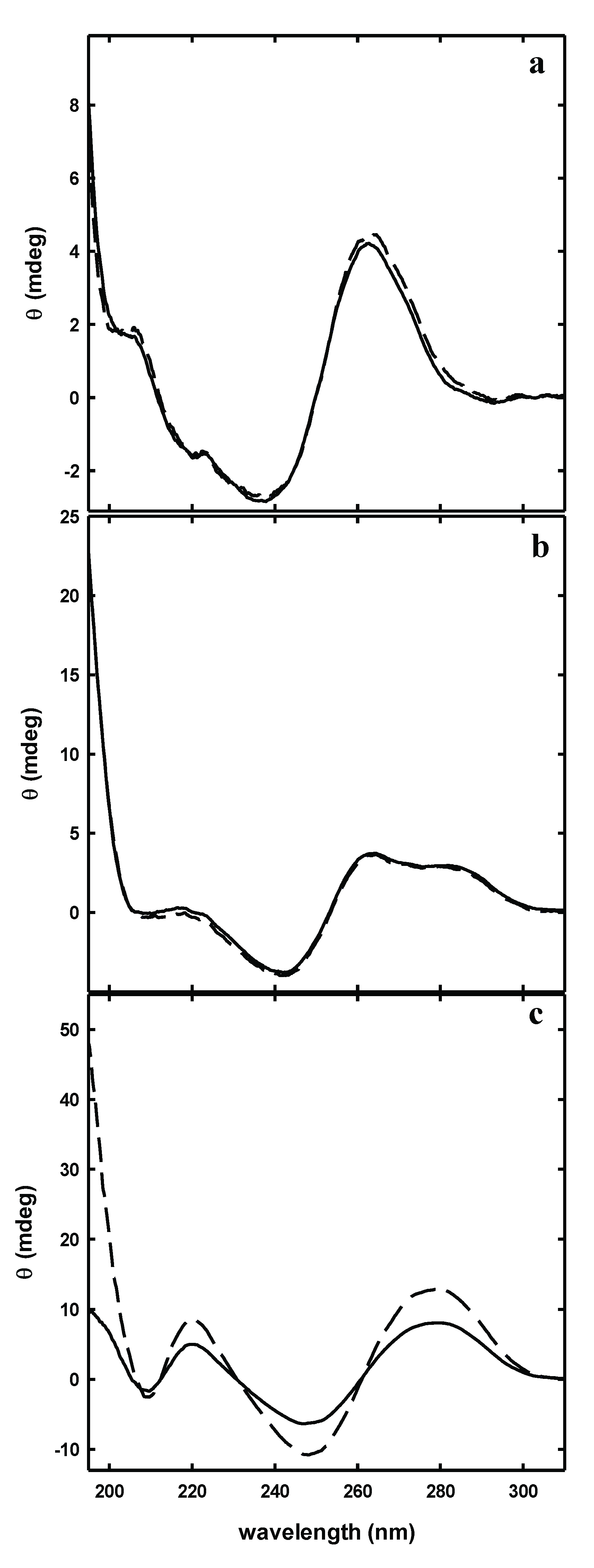
3.4. λ-Cro Peptide Mutants Characterization by Far-UV Circular Dichroism
3.5. Circular Dichroism Studies on Peptide-Aptamer Interaction
3.6. Fluorescence Characterization of Peptide-Aptamer Interactions
3.7. Molecular Dynamics Simulation
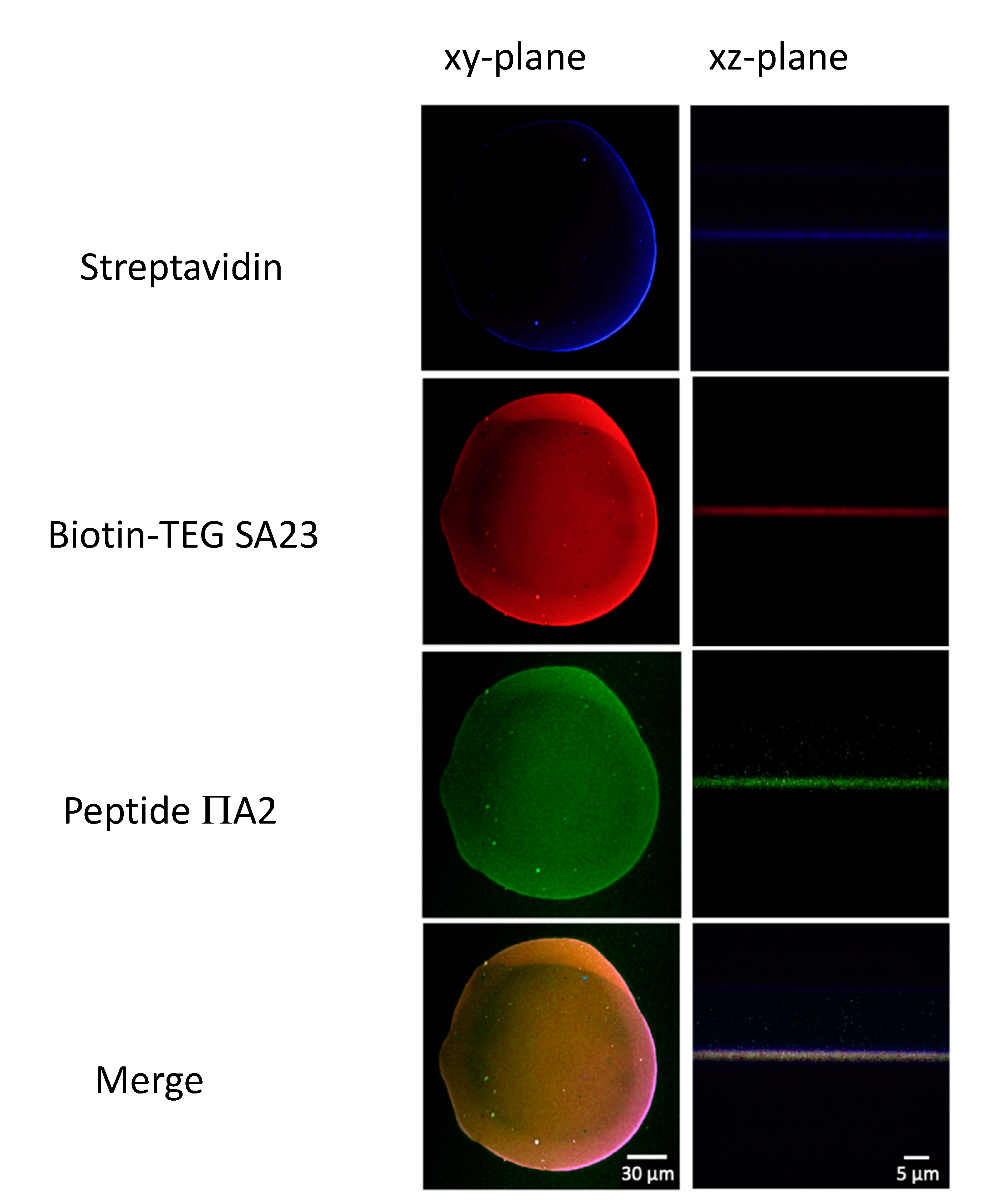
3.8. Aptamer Immobilization on Ns-ZrO2
3.9. Aptamers Hybridization with Peptides
3.10. Interaction of S. aureus with SA23 Aptamer
3.11. Aptamer:Fluorescent Peptide Complex Displacement by S. aureus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PCR | polymerase chain reaction |
| PNA | peptide nucleic acid |
| MALDI-TOF | matrix assisted laser desorption ionization time-of-flight |
| MS | mass spectrometry |
| SELEX | systematic evolution of ligands by exponential enrichment |
| CNT | carbon nanotubes |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2 |
| PEI-GA | glutaraldehyde pre-coated with polyethyleneimine |
| CFU | colony forming unit |
| PDB | protein data bank |
| MD | molecular dynamics |
| CD | circular dirchoism |
| ns-ZrOx | nanostructured zirconia |
| PMCS | pulsed microplasma cluster source |
| AFM | atomic force microscopy |
| PSIM | protein surface interaction microarray |
| TEG | triethylene glycol |
| PBS | phosphate buffed saline |
| RT | room temperature |
| RMSD | root mean square deviation |
References
- Ferreira, J.S.; Costa, W.L.R.; Cerqueira, E.S.; Carvalho, J.S.; Oliveira, L.C.; Almeida, R.C.C. Food handler-associated methicillin-resistant Staphylococcus aureus in public hospitals in Salvador, Brazil. Food Control 2014, 37, 395–400. [Google Scholar] [CrossRef]
- Török, E.; Day, N. Staphylococcal and streptococcal infections. Medicine (Baltimore) 2014, 33, 97–100. [Google Scholar] [CrossRef]
- Soriano, J.M.; Font, G.; Moltó, J.C.; Mañes, J. Enterotoxigenic staphylococci and their toxins in restaurant foods. Trends Food Sci. Technol. 2002, 13, 60–67. [Google Scholar] [CrossRef]
- David, M.Z.; Daum, R.S. Community-associated methicillin-resistant Staphylococcus aureus: Epidemiology and clinical consequences of an emerging epidemic. Clin. Microbiol. Rev. 2010, 23, 616–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannerman, T.L. Staphylococcus, Micrococcus, and other catalase-positive cocci that grow aerobically. In Manual of Clinical Microbiology, 8th ed.; Murray, P.R., Baron, E.J., Jorgensen, J.H., Pfaller, M.A., Yolken, R.H., Eds.; American Society for Microbiology: Washington, DC, USA, 2003; Volume 1, pp. 384–404. [Google Scholar]
- Evangelista, A.T.; Truant, A.L.; Bourbeau, P.P. Rapid Systems and Instruments for the Identification of Bacteria. In Manual of Commercial Methods in Clinical Microbiology; Press, A., Ed.; American Society of Microbiology: Washington, DC, USA, 2002; pp. 22–49. [Google Scholar] [CrossRef]
- Evangelista, A.T.; Truant, A.L. Rapid Systems and Instruments for Antimicrobial Susceptibility Testing of Bacteria. In Manual of Commercial Methods in Clinical Microbiology; Press, A., Ed.; American Society of Microbiology: Washington, DC, USA, 2002; pp. 413–428. [Google Scholar] [CrossRef]
- Weist, K.; Cimbal, A.K.; Lecke, C.; Kampf, G.; Ruden, H.; Vonberg, R.P. Evaluation of six agglutination tests for Staphylococcus aureus identification depending upon local prevalence of meticillin-resistant S. aureus (MRSA). J. Med. Microbiol. 2006, 55, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Chatzigeorgiou, K.S.; Sergentanis, T.N.; Tsiodras, S.; Hamodrakas, S.J.; Bagos, P.G. Phoenix 100 versus Vitek 2 in the identification of gram-positive and gram-negative bacteria: A comprehensive meta-analysis. J. Clin. Microbiol. 2011, 49, 3284–3291. [Google Scholar] [CrossRef] [Green Version]
- Zelada-Guillen, G.A.; Sebastian-Avila, J.L.; Blondeau, P.; Riu, J.; Rius, F.X. Label-free detection of Staphylococcus aureus in skin using real-time potentiometric biosensors based on carbon nanotubes and aptamers. Biosens. Bioelectron. 2012, 31, 226–232. [Google Scholar] [CrossRef]
- Watanabe, K.; Kuwata, N.; Sakamoto, H.; Amano, Y.; Satomura, T.; Suye, S. A smart DNA sensing system for detecting methicillin-resistant Staphylococcus aureus using modified nanoparticle probes. Biosens. Bioelectron. 2015, 67, 419–423. [Google Scholar] [CrossRef]
- Wang, H.Y.; Kim, S.; Kim, J.; Park, S.D.; Uh, Y.; Lee, H. Multiplex real-time PCR assay for rapid detection of methicillin-resistant staphylococci directly from positive blood cultures. J. Clin. Microbiol. 2014, 52, 1911–1920. [Google Scholar] [CrossRef] [Green Version]
- Peterson, L.R. Molecular laboratory tests for the diagnosis of respiratory tract infection due to Staphylococcus aureus. Clin. Infect. Dis. 2011, 52, S361–S366. [Google Scholar] [CrossRef] [Green Version]
- Frye, A.M.; Baker, C.A.; Rustvold, D.L.; Heath, K.A.; Hunt, J.; Leggett, J.E.; Oethinger, M. Clinical impact of a real-time PCR assay for rapid identification of staphylococcal bacteremia. J. Clin. Microbiol. 2012, 50, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, S.; Kliem, M. Mass spectrometry tools for the classification and identification of bacteria. Nat. Rev. Microbiol. 2010, 8, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Fleurbaaij, F.; van Leeuwen, H.; Klychnikov, O.; Kuijper, E.; Hensbergen, P. Mass spectrometry in clinical microbiology and infectious diseases. Chromatographia 2015, 78, 379–389. [Google Scholar] [CrossRef]
- Szabados, F.; Woloszyn, J.; Richter, C.; Kaase, M.; Gatermann, S. Identification of molecularly defined Staphylococcus aureus strains using matrix-assisted laser desorption/ionization time of flight mass spectrometry and the Biotyper 2.0 database. J. Med. Microbiol. 2010, 59, 787–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, O.; Tanaka, S.; Nagasawa, Z.; Hanaki, H.; Shobuike, T.; Miyamoto, H. Development of a novel matrix-assisted laser desorption/ionization time-of-flight mass spectrum (MALDI-TOF-MS)-based typing method to identify meticillin-resistant Staphylococcus aureus clones. J. Hosp. Infect. 2015, 90, 147–155. [Google Scholar] [CrossRef]
- Bernardo, K.; Pakulat, N.; Macht, M.; Krut, O.; Seifert, H.; Fleer, S.; Hunger, F.; Kronke, M. Identification and discrimination of Staphylococcus aureus strains using matrix-assisted laser desorption/ionization-time of flight mass spectrometry. Proteomics 2002, 2, 747–753. [Google Scholar] [CrossRef]
- Majumdar, T.; Chakraborty, R.; Raychaudhuri, U. Development of PEI-GA modified antibody based sensor for the detection of S. aureus in food samples. Food Biosci. 2013, 4, 38–45. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Sorokulova, I.B.; Vodyanoy, V.J.; Simonian, A.L. Lytic phage as a specific and selective probe for detection of Staphylococcus aureus—A surface plasmon resonance spectroscopic study. Biosens. Bioelectron. 2007, 22, 948–955. [Google Scholar] [CrossRef]
- Bandara, A.B.; Zuo, Z.; Ramachandran, S.; Ritter, A.; Heflin, J.R.; Inzana, T.J. Detection of methicillin-resistant staphylococci by biosensor assay consisting of nanoscale films on optical fiber long-period gratings. Biosens. Bioelectron. 2015, 70, 433–440. [Google Scholar] [CrossRef]
- Singh, A.; Poshtiban, S.; Evoy, S. Recent advances in bacteriophage based biosensors for food-borne pathogen detection. Sensors 2013, 13, 1763–1786. [Google Scholar] [CrossRef] [Green Version]
- Abdalhai, M.H.; Maximiano Fernandes, A.; Bashari, M.; Ji, J.; He, Q.; Sun, X. Rapid and sensitive detection of foodborne pathogenic bacteria (Staphylococcus aureus) using an electrochemical DNA genomic biosensor and its application in fresh beef. J. Agric. Food Chem. 2014, 62, 12659–12667. [Google Scholar] [CrossRef] [PubMed]
- Tombelli, S.; Minunni, M.; Mascini, M. Analytical applications of aptamers. Biosens. Bioelectron. 2005, 20, 2424–2434. [Google Scholar] [CrossRef]
- Schoukroun-Barnes, L.; White, R. Rationally designing aptamer sequences with reduced affinity for controlled sensor performance. Sensors 2015, 15, 7754–7767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Li, S.; Chen, L.; Ding, H.; Xu, H.; Huang, Y.; Li, J.; Liu, N.; Cao, W.; Zhu, Y.; et al. Combining use of a panel of ssDNA aptamers in the detection of Staphylococcus aureus. Nucleic Acids Res. 2009, 37, 4621–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.; Kim, G.; Park, S.; Lim, J.; Mo, C. Comparison of whole-cell SELEX methods for theidentification of Staphylococcus aureus-specific DNA aptamers. Sensors 2015, 15, 8884–8897. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, X.; Duan, N.; Wu, S.; Wang, Z.; Wei, X.; Wang, Y. Selection and characterization of DNA aptamers against Staphylococcus aureus enterotoxin C1. Food Chem. 2015, 166, 623–629. [Google Scholar] [CrossRef]
- Lian, Y.; He, F.; Wang, H.; Tong, F. A new aptamer/graphene interdigitated gold electrode piezoelectric sensor for rapid and specific detection of Staphylococcus aureus. Biosens. Bioelectron. 2015, 65, 314–319. [Google Scholar] [CrossRef]
- Abbaspour, A.; Norouz-Sarvestani, F.; Noori, A.; Soltani, N. Aptamer-conjugated silver nanoparticles for electrochemical dual-aptamer-based sandwich detection of Staphylococcus aureus. Biosens. Bioelectron. 2015, 68, 149–155. [Google Scholar] [CrossRef]
- Ramlal, S.; Mondal, B.; Lavu, P.S.; Bhavanashri, N.; Kingston, J. Capture and detection of Staphylococcus aureus with dual labeled aptamers to cell surface components. Int. J. Food Microbiol. 2018, 265, 74–83. [Google Scholar] [CrossRef]
- Alizadeh, N.; Memar, M.Y.; Moaddab, S.R.; Kafil, H.S. Aptamer-assisted novel technologies for detecting bacterial pathogens. Biomed. Pharmacother. 2017, 93, 737–745. [Google Scholar] [CrossRef]
- Mairal, T.; Ozalp, V.C.; Lozano Sanchez, P.; Mir, M.; Katakis, I.; O’Sullivan, C.K. Aptamers: Molecular tools for analytical applications. Anal. Bioanal. Chem. 2008, 390, 989–1007. [Google Scholar] [CrossRef] [PubMed]
- Duzgun, A.; Maroto, A.; Mairal, T.; O’Sullivan, C.; Rius, F.X. Solid-contact potentiometric aptasensor based on aptamer functionalized carbon nanotubes for the direct determination of proteins. Analyst 2010, 135, 1037–1041. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Yang, C.Y.; Sun, R.L.; Cheng, Y.F.; Kao, W.C.; Yang, P.C. Rapid single cell detection of Staphylococcus aureus by aptamer-conjugated gold nanoparticles. Sci. Rep. 2013, 3, 1863. [Google Scholar] [CrossRef] [PubMed]
- Langer, R.; Tirrell, D.A. Designing materials for biology and medicine. Nature 2004, 428, 487–492. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, J. Functional DNA nanotechnology: Emerging applications of DNAzymes and aptamers. Curr. Opin. Biotechnol. 2006, 17, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Mitragotri, S.; Lahann, J. Physical approaches to biomaterial design. Nat. Mater. 2009, 8, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Kohn, J. New approaches to biomaterials design. Nat. Mater. 2004, 3, 745–747. [Google Scholar] [CrossRef]
- Negahdary, M. Aptamers in nanostructure-based electrochemical biosensors for cardiac biomarkers and cancer biomarkers: A review. Biosens. Bioelectron. 2020, 152, 112018. [Google Scholar] [CrossRef]
- Zang, Y.; Lei, J.; Ju, H. Principles and applications of photoelectrochemical sensing strategies based on biofunctionalized nanostructures. Biosens. Bioelectron. 2017, 96, 8–16. [Google Scholar] [CrossRef]
- Scopelliti, P.E.; Bongiorno, G.; Milani, P. High-throughput tools for the study of protein-nanostructured surface interaction. Comb. Chem. High Throughput Screen. 2011, 14, 205–216. [Google Scholar]
- Scopelliti, P.E.; Borgonovo, A.; Indrieri, M.; Giorgetti, L.; Bongiorno, G.; Carbone, R.; Podesta, A.; Milani, P. The effect of surface nanometre-scale morphology on protein adsorption. PLoS ONE 2010, 5, e11862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.; Ryu, B.; Deng, Q.; Pan, B.; Song, Y.; Tian, Y.; Alam, H.B.; Li, Y.; Liang, X.; Kurabayashi, K. An integrated plasmo-photoelectronic nanostructure biosensor detects an infection biomarker accompanying cell death in neutrophils. Small 2020, 16, e1905611. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Curras, N.; Scida, K.; Ploense, K.L.; Kippin, T.E.; Plaxco, K.W. High surface area electrodes generated via electrochemical roughening improve the signaling of electrochemical aptamer-based biosensors. Anal. Chem. 2017, 89, 12185–12191. [Google Scholar] [CrossRef]
- Zuppolini, S.; Quero, G.; Consales, M.; Diodato, L.; Vaiano, P.; Venturelli, A.; Santucci, M.; Spyrakis, F.; Costi, M.P.; Giordano, M.; et al. Label-free fiber optic optrode for the detection of class C β-lactamases expressed by drug resistant bacteria. Biomed. Opt. Express 2017, 8, 5191–5205. [Google Scholar] [CrossRef] [Green Version]
- Quero, G.; Zuppolini, S.; Consales, M.; Diodato, L.; Vaiano, P.; Venturelli, A.; Santucci, M.; Spyrakis, F.; Costi, M.P.; Giordano, M.; et al. Long period fiber grating working in reflection mode as valuable biosensing platform for the detection of drug resistant bacteria. Sens. Actuator B Chem. 2016, 230, 510–520. [Google Scholar] [CrossRef]
- Marabotti, A.; Spyrakis, F.; Facchiano, A.; Cozzini, P.; Alberti, S.; Kellogg, G.E.; Mozzarelli, A. Energy-based prediction of amino acid-nucleotide base recognition. J. Comput. Chem. 2008, 29, 1955–1969. [Google Scholar] [CrossRef]
- Cozzini, P.; Fornabaio, M.; Marabotti, A.; Abraham, D.J.; Kellogg, G.E.; Mozzarelli, A. Simple, intuitive calculations of free energy of binding for protein-ligand complexes. 1. Models without explicit constrained water. J. Med. Chem. 2002, 45, 2469–2483. [Google Scholar] [CrossRef]
- Ahmed, M.H.; Spyrakis, F.; Cozzini, P.; Tripathi, P.K.; Mozzarelli, A.; Scarsdale, J.N.; Safo, M.A.; Kellogg, G.E. Bound water at protein-protein interfaces: Partners, roles and hydrophobic bubbles as a conserved motif. PLoS ONE 2011, 6, e24712. [Google Scholar] [CrossRef] [Green Version]
- Amadasi, A.; Spyrakis, F.; Cozzini, P.; Abraham, D.J.; Kellogg, G.E.; Mozzarelli, A. Mapping the energetics of water-protein and water-ligand interactions with the “natural” HINT forcefield: Predictive tools for characterizing the roles of water in biomolecules. J. Mol. Biol. 2006, 358, 289–309. [Google Scholar] [CrossRef]
- Spyrakis, F.; Cozzini, P.; Bertoli, C.; Marabotti, A.; Kellogg, G.E.; Mozzarelli, A. Energetics of the protein-DNA-water interaction. BMC Struct. Biol. 2007, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Autiero, I.; Ruvo, M.; Improta, R.; Vitagliano, L. The intrinsic flexibility of the aptamer targeting the ribosomal protein S8 is a key factor for the molecular recognition. Biochim. Biophys. Acta 2018, 1862, 1006–1016. [Google Scholar] [CrossRef] [PubMed]
- Hermann, T.; Patel, D.J. Adaptive recognition by nucleic acid aptamers. Science 2000, 287, 820–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrino, S.; Annoni, C.; Contini, A.; Clerici, F.; Gelmi, M.L. Expedient chemical synthesis of 75mer DNA binding domain of MafA: An insight on its binding to insulin enhancer. Amino Acids 2012, 43, 1995–2003. [Google Scholar] [CrossRef]
- Li, Y.; Yan, X.; Feng, X.; Wang, J.; Du, W.; Wang, Y.; Chen, P.; Xiong, L.; Liu, B.-F. Agarose-based microfluidic device for point-of-care concentration and detection of pathogen. Anal. Chem. 2014, 86, 10653–10659. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Gao, H.; Zhang, Y.; Wang, X.; Chen, Y.; Sun, W. Electrochemical DNA biosensor with chitosan-Co3O4 nanorod-graphene composite for the sensitive detection of Staphylococcus aureus nuc gene sequence. Bioelectrochemistry 2012, 88, 42–47. [Google Scholar] [CrossRef]
- Parisien, M.; Major, F. The MC-Fold and MC-Sym pipeline infers RNA structure from sequence data. Nature 2008, 452, 51–55. [Google Scholar] [CrossRef]
- Albright, R.A.; Matthews, B.W. Crystal structure of λ-Cro bound to a consensus operator at 3.0 Å resolution. J. Mol. Biol. 1998, 280, 137–151. [Google Scholar] [CrossRef]
- Hess, B.; Kutzner, C.; Van Der Spoel, D.; Lindahl, E. GRGMACS 4: Algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Izadi, S.; Anandakrishnan, R.; Onufriev, A.V. Building water models: A different approach. J. Phys. Chem. Lett. 2014, 5, 3863–3871. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Grigera, J.R.; Straatsma, T.P. The missing term in effective pair potentials. J. Phys. Chem. 1987, 91, 6269–6271. [Google Scholar] [CrossRef]
- Piacenti, V.; Langella, E.; Autiero, I.; Nolan, J.C.; Piskareva, O.; Adamo, M.F.A.; Saviano, M.; Moccia, M. A combined experimental and computational study on peptide nucleic acid (PNA) analogues of tumor suppressive miRNA-34a. Bioorg. Chem. 2019, 91, 103165. [Google Scholar] [CrossRef] [PubMed]
- Abbruzzetti, S.; Grandi, E.; Bruno, S.; Faggiano, S.; Spyrakis, F.; Mozzarelli, A.; Cacciatori, E.; Dominici, P.; Viappiani, C. Ligand migration in nonsymbiotic hemoglobin AHb1 from Arabidopsis thaliana. J. Phys. Chem. B 2007, 111, 12582–12590. [Google Scholar] [CrossRef]
- Chawla, M.; Autiero, I.; Oliva, R.; Cavallo, L. Energetics and dynamics of the non-natural fluorescent 4AP:DAP base pair. PCCP 2018, 20, 3699–3709. [Google Scholar] [CrossRef] [PubMed]
- Autiero, I.; Saviano, M.; Langella, E. In silico investigation and targeting of amyloid β oligomers of different size. Molecular BioSystems 2013, 9, 2118–2124. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- DeLano, W.L. PyMOL. DeLano Sci. San Carlos CA 2002, 700, 5. [Google Scholar]
- Daura, X.; Gademann, K.; Jaun, B.; Seebach, D.; van Gunsteren, W.F.; Mark, A.E. Peptide folding: When simulation meets experiment. Angew. Chem. Int. Ed. 1999, 38, 236–240. [Google Scholar] [CrossRef]
- Macut, H.; Hu, X.; Tarantino, D.; Gilardoni, E.; Clerici, F.; Regazzoni, L.; Contini, A.; Pellegrino, S.; Luisa Gelmi, M. Tuning PFKFB3 bisphosphatase activity through allosteric interference. Sci. Rep. 2019, 9, 20333. [Google Scholar] [CrossRef] [Green Version]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, W668–W673. [Google Scholar] [CrossRef] [Green Version]
- Piseri, P.; Tafreshi, H.V.; Milani, P. Manipulation of nanoparticles in supersonic beams for the production of nanostructured materials. Curr. Opin. Solid State Mater. Sci. 2004, 8, 195–202. [Google Scholar] [CrossRef]
- Borghi, F.; Sogne, E.; Lenardi, C.; Podestà, A.; Merlini, M.; Ducati, C.; Milani, P. Cluster-assembled cubic zirconia films with tunable and stable nanoscale morphology against thermal annealing. J. Appl. Phys. 2016, 120, 055302. [Google Scholar] [CrossRef] [Green Version]
- Podestà, A.; Bongiorno, G.; Scopelliti, P.E.; Bovio, S.; Milani, P.; Semprebon, C.; Mistura, G. Cluster-Assembled nanostructured titanium oxide films with tailored wettability. J. Phys. Chem. C 2009, 113, 18264–18269. [Google Scholar] [CrossRef]
- Giorgetti, L.; Bongiorno, G.; Podestà, A.; Berlanda, G.; Scopelliti, P.E.; Carbone, R.; Milani, P. Adsorption and stability of streptavidin on cluster-assembled nanostructured TiOx films. Langmuir 2008, 24, 11637–11644. [Google Scholar] [CrossRef] [PubMed]
- Gailite, L.; Scopelliti, P.E.; Sharma, V.K.; Indrieri, M.; Podestà, A.; Tedeschi, G.; Milani, P. Nanoscale roughness affects the activity of enzymes adsorbed on cluster-assembled titania films. Langmuir 2014, 30, 5973–5981. [Google Scholar] [CrossRef]
- Mazumder, A.; Maiti, A.; Roy, K.; Roy, S. A Synthetic Peptide mimic of λ-Cro shows sequence-specific binding in vitro and in vivo. ACS Chem. Biol. 2012, 7, 1084–1094. [Google Scholar] [CrossRef]
- Ohlendorf, D.H.; Tronrud, D.E.; Matthews, B.W. Refined structure of Cro repressor protein from bacteriophage λ suggests both flexibility and plasticity11Edited by P. E. Wright. J. Mol. Biol. 1998, 280, 129–136. [Google Scholar] [CrossRef]
- Pellegrino, S.; Ferri, N.; Colombo, N.; Cremona, E.; Corsini, A.; Fanelli, R.; Gelmi, M.L.; Cabrele, C. Synthetic peptides containing a conserved sequence motif of the Id protein family modulate vascular smooth muscle cell phenotype. Bioorg. Med. Chem. Lett. 2009, 19, 6298–6302. [Google Scholar] [CrossRef]
- Cho, E.J.; Collett, J.R.; Szafranska, A.E.; Ellington, A.D. Optimization of aptamer microarray technology for multiple protein targets. Anal. Chim. Acta 2006, 564, 82–90. [Google Scholar] [CrossRef]
- Dell’Acqua, M.; Ronda, L.; Piano, R.; Pellegrino, S.; Clerici, F.; Rossi, E.; Mozzarelli, A. Based polarity-sensitive dyes. J. Org. Chem. 2015, 80, 10939–10954. [Google Scholar]
- Pirovano, V.; Marchetti, M.; Carbonaro, J.; Brambilla, E.; Rossi, E.; Ronda, L.; Abbiati, G. Synthesis and photophysical properties of isocoumarin-based D-π-A systems. Dyes Pigment. 2020, 173, 107917. [Google Scholar] [CrossRef] [Green Version]










| Peptide | Sequence | MW (Da) |
|---|---|---|
| IA | Ac-GQTKTAKDLGVYKSAIEEAIHAG | 2428.73 |
| IAser | Ac-GQTKTAKDLGVYKDAIEEAIHAG | 2456.73 |
| IB2A | Ac-GQTKTAKDLGVYDSAIEEAIHAG | 2415.63 |
| IIA | Ac-GQTKTAKDLGVYESAIEEAIHAG | 2429.67 |
| IIA2 | Ac-GQTKTAKDLGVYEDAIEKAIHAG | 2456.74 |
| IIA3 | Ac-GQTKTAKDLGVYEDAIEFAIHAG | 2475.74 |
| IIA2M | Carboxyfluorescein-GQTKTAKDLGVYEDAIEKAIHAG | 2773 |
| IA3M | Carboxyfluorescein- GQTKTAKDLGVYEDAIEFAIHAG | 2792 |
| Peptide | Helices | Strand | Turns | Disordered |
|---|---|---|---|---|
| IA | 17% | 28% | 18% | 36% |
| IAser | 21% | 24% | 20% | 35% |
| IB2 | 21% | 26% | 21% | 32% |
| IIA | 19% | 24% | 20% | 37% |
| IIA2 | 7% | 32% | 23% | 37% |
| IIA3 | 5% | 36% | 17% | 40% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronda, L.; Tonelli, A.; Sogne, E.; Autiero, I.; Spyrakis, F.; Pellegrino, S.; Abbiati, G.; Maffioli, E.; Schulte, C.; Piano, R.; et al. Rational Design of a User-Friendly Aptamer/Peptide-Based Device for the Detection of Staphylococcus aureus. Sensors 2020, 20, 4977. https://0-doi-org.brum.beds.ac.uk/10.3390/s20174977
Ronda L, Tonelli A, Sogne E, Autiero I, Spyrakis F, Pellegrino S, Abbiati G, Maffioli E, Schulte C, Piano R, et al. Rational Design of a User-Friendly Aptamer/Peptide-Based Device for the Detection of Staphylococcus aureus. Sensors. 2020; 20(17):4977. https://0-doi-org.brum.beds.ac.uk/10.3390/s20174977
Chicago/Turabian StyleRonda, Luca, Alessandro Tonelli, Elisa Sogne, Ida Autiero, Francesca Spyrakis, Sara Pellegrino, Giorgio Abbiati, Elisa Maffioli, Carsten Schulte, Riccardo Piano, and et al. 2020. "Rational Design of a User-Friendly Aptamer/Peptide-Based Device for the Detection of Staphylococcus aureus" Sensors 20, no. 17: 4977. https://0-doi-org.brum.beds.ac.uk/10.3390/s20174977