Reclamation of Saline–Sodic Soils with Combined Amendments: Impact on Quinoa Performance and Biological Soil Quality


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Soil Sampling
2.3. Germination Assay
2.4. Leaching
2.5. Amendments
2.6. Experimental Procedure
2.7. Plant Pparameters Studied
2.8. Seed Parameters Measured
2.9. Chemical and Biological Soil Analysis, and Leaf Tissue Analysis
2.10. Statistical Analysis
3. Results
3.1. Effects of the Amendments on Growth and Physiological Parameters of Quinoa Plants
3.2 Polyphenols and Protein Content in Quinoa Seeds
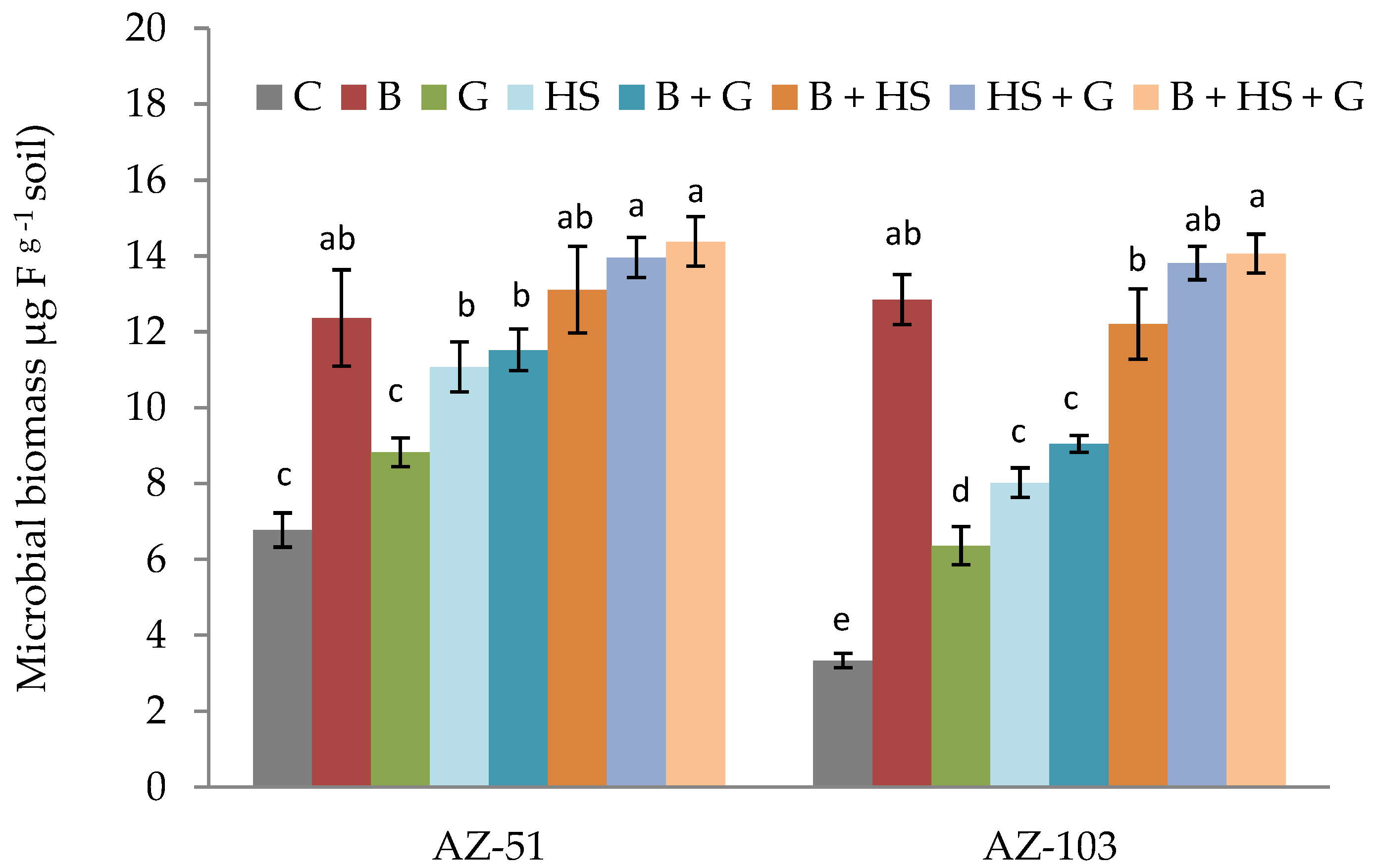
3.3. Effects of Amendments on the Chemical and Biological Properties of Saline–Sodic Soils
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Young, J.; Udeigwe, T.K.; Weindorf, D.C.; Kandakji, T.; Gautam, P.; Mahmoud, M.A. Evaluating management-induced soil salinization in golf courses in semi-arid landscapes. Solid Earth 2015, 6, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Wang, Z.; Meixner, F.X.; Yang, F.; Wu, H.; Chen, X. Biogeochemical characterizations and reclamation strategies of saline sodic soil in northeastern China. CLEAN–Soil, Air, Water 2010, 38, 1010–1016. [Google Scholar] [CrossRef]
- Wong, V.N.; Dalal, R.C.; Greene, R.S. Salinity and sodicity effects on respiration and microbial biomass of soil. Biol. Fertil. Soils 2008, 44, 943–953. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: a review. Ecotox. Environ. Safe. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Levizou, E.; Ntatsi, G.; Fernandes, Â.; Petrotos, K.; Akoumianakis, K.; Barros, L.; Ferreira, I.C. Salinity effect on nutritional value, chemical composition and bioactive compounds content of Cichorium spinosum L. Food Chem. 2017, 214, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, K.B.; Aloisi, I.; Del Duca, S.; Canelo, V.; Torrigiani, P.; Silva, H.; Biondi, S. Salares versus coastal ecotypes of quinoa: salinity responses in Chilean landraces from contrasting habitats. Plant Physiol. Bioch. 2016, 101, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Razzaghi, F.; Jacobsen, S.-E.; Jensen, C.R.; Andersen, M.N. Ionic and photosynthetic homeostasis in quinoa challenged by salinity and drought–mechanisms of tolerance. Funct. Plant Biol. 2015, 42, 136–148. [Google Scholar] [CrossRef]
- Nowak, V.; Du, J.; Charrondière, U.R. Assessment of the nutritional composition of quinoa (Chenopodium quinoa Willd.). Food Chem. 2016, 193, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Hariadi, Y.; Marandon, K.; Tian, Y.; Jacobsen, S.-E.; Shabala, S. Ionic and osmotic relations in quinoa (Chenopodium quinoa Willd.) plants grown at various salinity levels. J. Exp. Bot. 2010, 62, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ramzani, P.M.A.; Shan, L.; Anjum, S.; Ronggui, H.; Iqbal, M.; Virk, Z.A.; Kausar, S. Improved quinoa growth, physiological response, and seed nutritional quality in three soils having different stresses by the application of acidified biochar and compost. Plant Physiol. Bioch. 2017, 116, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Yazdanpanah, N.; Pazira, E.; Neshat, A.; Mahmoodabadi, M.; Sinobas, L.R. Reclamation of calcareous saline sodic soil with different amendments (II): Impact on nitrogen, phosphorous and potassium redistribution and on microbial respiration. Agr. Water Manage. 2013, 120, 39–45. [Google Scholar] [CrossRef]
- Agriculture Organization. The State of Food Insecurity in the World 2014; FAO: Rome, Italy, 2014. [Google Scholar]
- Mahmoodabadi, M.; Yazdanpanah, N.; Sinobas, L.R.; Pazira, E.; Neshat, A. Reclamation of calcareous saline sodic soil with different amendments (I): Redistribution of soluble cations within the soil profile. Agr. Water Manage. 2013, 120, 30–38. [Google Scholar] [CrossRef]
- Yaduvanshi, N.P.S.; Sharma, D.R. Tillage and residual organic manures/chemical amendment effects on soil organic matter and yield of wheat under sodic water irrigation. Soil Till. Res. 2008, 98, 11–16. [Google Scholar] [CrossRef]
- Yaduvanshi, N.P. Nutrient management for sustained crop productivity in sodic soils: a review. In Soil Salinity Management in Agriculture: Technological Advances and Applications; Gupta, S.K., Goyal, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 365–394. [Google Scholar]
- Tejada, M.; Garcia, C.; Gonzalez, J.L.; Hernandez, M.T. Use of organic amendment as a strategy for saline soil remediation: influence on the physical, chemical and biological properties of soil. Soil Biolo. Bioch. 2006, 38, 1413–1421. [Google Scholar] [CrossRef]
- Gupta, M.; Srivastava, P.K.; NIRANJAN, A.; TEWARI, S.K. Use of a bioaugmented organic soil amendment in combination with gypsum for Withania somnifera growth on sodic soil. Pedosphere 2016, 26, 299–309. [Google Scholar] [CrossRef]
- Clemente, R.; Bernal, M.P. Fractionation of heavy metals and distribution of organic carbon in two contaminated soils amended with humic acids. Chemosphere 2006, 64, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Ciarkowska, K. Effect of fertilization on the structure of upland grassland soil. Pol. J. Environ. Stud. 2010, 19, 693–697. [Google Scholar]
- Ciarkowska, K.; Solek-Podwika, K.; Filipek-Mazur, B.; Tabak, M. Comparative effects of lignite-derived humic acids and FYM on soil properties and vegetable yield. Geoderma 2017, 303, 85–92. [Google Scholar] [CrossRef]
- Lehmann, J.; Joseph, S. Biochar for environmental management: An introduction. In Biochar for Environmental Management: Science, Technology and Implementation, 2nd ed.; Lehmann, J., Joseph, S., Eds.; Earths can: London, UK, 2015; pp. 1–15. [Google Scholar]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota–a review. Soil Biolo. Bioch. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Chaganti, V.N.; Crohn, D.M. Evaluating the relative contribution of physiochemical and biological factors in ameliorating a saline–sodic soil amended with composts and biochar and leached with reclaimed water. Geoderma 2015, 259, 45–55. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, X.; Chen, L.; Wang, Z.; Xia, Y.; Zhang, Y.; Wang, H.; Luo, X.; Xing, B. Enhanced growth of halophyte plants in biochar-amended coastal soil: roles of nutrient availability and rhizosphere microbial modulation. Plant Cell Environ. 2018, 41, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Soil Survey Staff. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; US Government Printing Office: Washington, DC, USA, 1999.
- Cade-Idepe. Consultores en Ingeniería. 2004. Cuenca del Río Huasco: diagnóstico y clasificación de agua según objetivos de calidad [en línea]. Ministerio de Obras Públicas, Chile. Available online: http://portal.mma.gob.cl/diagnostico-y-clasificacion-de-cursos-y-cuerpos-de-agua-segun-objetivos-de-calidad/ (accessed on 15 January 2018).
- Gatica, S. Estudio de Caso; diagnóstico de los suelos y de las aguas para regadío del sector de Huasco Bajo, Región de Atacama, Chile. Tesis Pregrado, Universidad de Concepción, Chillán, 2013. [Google Scholar]
- Muñoz, C.; Quilodrán, C.; Navia, R. Evaluation of biochar-plant extracts complexes on soil nitrogen dynamics. J. Biobased Mater. Bio. 2014, 8, 377–385. [Google Scholar] [CrossRef]
- Sayegh, A.H.; Khan, N.A.; Khan, P.; Ryan, J. Factors Affecting Gypsum and Cation Exchange Capacity Determinations in Gypsiferous SOILS1. Soil Sci. 1978, 125, 294–300. [Google Scholar] [CrossRef]
- Kim, H.-S.; Kim, K.-R.; Yang, J.E.; Ok, Y.S.; Owens, G.; Nehls, T.; Wessolek, G.; Kim, K.-H. Effect of biochar on reclaimed tidal land soil properties and maize (Zea mays L.) response. Chemosphere 2016, 142, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Quezada, C.; Sandoval, M.; Stolpe, N.; Olmos, D. Uso de enmiendas orgánicas y químicas para la remediación de suelos inundados por el tsunami de 2010 en la zona de Tomé, Chile. Chilean J. Agric. Anim. Sci. 2016, 32, 13–22. [Google Scholar]
- Lachman, J.; Dudjak, J.; Orsák, M.; Pivec, V. Effect of accelerated ageing on the content and composition of polyphenolic complex of wheat (Triticum aestivum L.) grains. Plant Soil Environ. 2003, 49, 1–7. [Google Scholar] [CrossRef]
- Sadzawka, M.A.R.; Carrasco, A.M.R.; Demanet, F.R.; Flores, P.H.; Grez, Z.R.; Mora, G.M.L.; Neaman, A. Métodos de Análisis Recomendados Para los Suelos de Chile. In Proceedings of the INIA Serie Actas Nº 34, Santiago, Chile, 2006; Revisión 2006. Available online: http://www.cofes.org.ar/descargas/relas/4_jornada/Manual_chilenos_suelos_2006.pdf (accessed on 7 August 2018).
- Alef, K. Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 232–233. [Google Scholar]
- Sadzawka, M.A.R.; Carrasco, A.M.R.; Demanet, F.R.; Flores, P.H.; Grez, Z.R.; Mora, G.M.L.; Neaman, A. Métodos de Análisis de Tejidos Vegetales. In Proceedings of the INIA Serie Actas Nº 40, Santiago, Chile, 2007; Available online: http://www.cofes.org.ar/descargas/relas/4_jornada/Manual_chilenos_tejido_vegetal.pdf (accessed on 7 August 2018).
- Qadir, M.; Ghafoor, A.; Murtaza, G. Amelioration strategies for saline soils: a review. Land Degrad. Dev. 2000, 11, 501–521. [Google Scholar] [CrossRef]
- Kammann, C.I.; Linsel, S.; Gößling, J.W.; Koyro, H.-W. Influence of biochar on drought tolerance of Chenopodium quinoa Willd and on soil–plant relations. Plant Soil 2011, 345, 195–210. [Google Scholar] [CrossRef]
- Arjumend, T.; Abbasi, M.K.; Rafique, E. Effects of lignite-derived humic acid on some selected soil properties, growth and nutrient uptake of wheat (Triticum aestivum L.) grown under greenhouse conditions. Pakistan J. Bot. 2015, 47, 2231–2238. [Google Scholar]
- Akhtar, S.S.; Andersen, M.N.; Liu, F. Biochar mitigates salinity stress in potato. J. Agron. Crop Sci. 2015, 201, 368–378. [Google Scholar] [CrossRef]
- Gill, J.S.; Sale, P.W.G.; Peries, R.R.; Tang, C. Changes in soil physical properties and crop root growth in dense sodic subsoil following incorporation of organic amendments. Field Crop. Res. 2009, 114, 137–146. [Google Scholar] [CrossRef]
- Khan, M.A.; Shirazi, M.U.; Khan, M.A.; Mujtaba, S.M.; Islam, E.; Mumtaz, S.; Shereen, A.; Ansari, R.U.; Ashraf, M.Y. Role of proline, K/Na ratio and chlorophyll content in salt tolerance of wheat (Triticum aestivum L.). Pak. J. Bot. 2009, 41, 633–638. [Google Scholar]
- González, J.A.; Ponessa, G.I.; Buedo, S.; Mercado, M.I.; Prado, F.E. Asimilación fotosintética máxima en variedades de quinoa (Chenopodium quinoa) de diferentes orígenes geográficos y su relación con la morfología foliar. Lilloa 2014, 51. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Andersen, M.N.; Liu, F. Residual effects of biochar on improving growth, physiology and yield of wheat under salt stress. Agr. Water Manage. 2015, 158, 61–68. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Muscolo, A.; Vianello, A. Physiological effects of humic substances on higher plants. Soil Biolo. Bioch. 2002, 34, 1527–1536. [Google Scholar] [CrossRef]
- Benzarti, M.; Rejeb, K.B.; Debez, A.; Messedi, D.; Abdelly, C. Photosynthetic activity and leaf antioxidative responses of Atriplex portulacoides subjected to extreme salinity. Acta Physiol. Plant. 2012, 34, 1679–1688. [Google Scholar] [CrossRef]
- Lashari, M.S.; Liu, Y.; Li, L.; Pan, W.; Fu, J.; Pan, G.; Zheng, J.; Zheng, J.; Zhang, X.; Yu, X. Effects of amendment of biochar-manure compost in conjunction with pyroligneous solution on soil quality and wheat yield of a salt-stressed cropland from Central China Great Plain. Field Crop. Res. 2013, 144, 113–118. [Google Scholar] [CrossRef]
- Trivedi, P.; Singh, K.; Pankaj, U.; Verma, S.K.; Verma, R.K.; Patra, D.D. Effect of organic amendments and microbial application on sodic soil properties and growth of an aromatic crop. Ecol. Eng. 2017, 102, 127–136. [Google Scholar] [CrossRef]
- Turan, M.A.; Baris, B.A.; Katkat, A.V.; Celik, H. The effects of soil-applied humic substances to the dry weight and mineral nutrient uptake of maize plants under soil-salinity conditions. Not. Bot. Horti. Agro. 2011, 39, 171. [Google Scholar] [CrossRef]
- Spokas, K.A.; Baker, J.M.; Reicosky, D.C. Ethylene: potential key for biochar amendment impacts. Plant Soil 2010, 333, 443–452. [Google Scholar] [CrossRef]
- Agegnehu, G.; Bird, M.I.; Nelson, P.N.; Bass, A.M. The ameliorating effects of biochar and compost on soil quality and plant growth on a Ferralsol. Soil Res. 2015, 53, 1–12. [Google Scholar] [CrossRef]
- Mahmood, I.A.; Salim, M.; Ali, A.; Arshadullah, M.; Zaman, B.; Mir, A. Impact of calcium sulphate and calcium carbide on nitrogen use efficiency of wheat in normal and saline sodic soils. Soil Environ. 2009, 28, 29–37. [Google Scholar]
- Hartz, T.K.; Bottoms, T.G. Humic substances generally ineffective in improving vegetable crop nutrient uptake or productivity. HortScience 2010, 45, 906–910. [Google Scholar]
- Srivastava, A.K.; Srivastava, S.; Lokhande, V.H.; D’Souza, S.F.; Suprasanna, P. Salt stress reveals differential antioxidant and energetics responses in glycophyte (Brassica juncea L.) and halophyte (Sesuvium portulacastrum L.). Front. Env. Sci. 2015, 3, 19. [Google Scholar] [CrossRef]
- Ramzani, P.M.A.; Khalid, M.; Naveed, M.; Ahmad, R.; Shahid, M. Iron biofortification of wheat grains through integrated use of organic and chemical fertilizers in pH affected calcareous soil. Plant Phy. Bioch. 2016, 104, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, A.K.; Lewińska, K.; Iqbal, J.; Ali, Q.; Iqbal, M.; Abbas, F.; Tauqeer, H.M.; Ramzani, P.M.A. Improvement in productivity, nutritional quality, and antioxidative defense mechanisms of sunflower (Helianthus annuus L.) and maize (Zea mays L.) in nickel contaminated soil amended with different biochar and zeolite ratios. J. Environ. Manage. 2018, 218, 256–270. [Google Scholar] [CrossRef] [PubMed]
- Dahlawi, S.; Naeem, A.; Rengel, Z.; Naidu, R. Biochar application for the remediation of salt-affected soils: Challenges and opportunities. Sci. Total Environ. 2018, 625, 320–335. [Google Scholar]
- Nan, J.; Chen, X.; Wang, X.; Lashari, M.S.; Wang, Y.; Guo, Z.; Du, Z. Effects of applying flue gas desulfurization gypsum and humic acid on soil physicochemical properties and rapeseed yield of a saline-sodic cropland in the eastern coastal area of China. J. Soil. Sediment. 2016, 16, 38–50. [Google Scholar] [CrossRef]
- Chaganti, V.N.; Crohn, D.M.; Šimŭnek, J. Leaching and reclamation of a biochar and compost amended saline–sodic soil with moderate SAR reclaimed water. Agr. Water Manage. 2015, 158, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Jalali, M. Effect of sodium and magnesium on kinetics of potassium release in some calcareous soils of western Iran. Geoderma 2008, 145, 207–215. [Google Scholar] [CrossRef]
- Çimrin, K.M.; Türkmen, Ö.; Turan, M.; Tuncer, B. Phosphorus and humic acid application alleviate salinity stress of pepper seedling. Afr.J. Biotechnol. 2010, 9, 5845–5851. [Google Scholar]
- Bhaduri, D.; Saha, A.; Desai, D.; Meena, H.N. Restoration of carbon and microbial activity in salt-induced soil by application of peanut shell biochar during short-term incubation study. Chemosphere 2016, 148, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Pukalchik, M.; Mercl, F.; Panova, M.; Břendová, K.; Terekhova, V.A.; Tlustoš, P. The improvement of multi-contaminated sandy loam soil chemical and biological properties by the biochar, wood ash, and humic substances amendments. Environ. Pollut. 2017, 229, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Whitman, T.; Pepe-Ranney, C.; Enders, A.; Koechli, C.; Campbell, A.; Buckley, D.H.; Lehmann, J. Dynamics of microbial community composition and soil organic carbon mineralization in soil following addition of pyrogenic and fresh organic matter. ISME J. 2016, 10, 2918. [Google Scholar] [CrossRef] [PubMed]
- Lipczynska-Kochany, E. Humic substances, their microbial interactions and effects on biological transformations of organic pollutants in water and soil: A review. Chemosphere 2018, 202, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Kumari, S.; Chakraborty, A.; Gupta, A.; Chakrabarti, K.; Bandyapadhyay, B.K. Microbial biomass and its activities in salt-affected coastal soils. Biol. Fertil Soils 2006, 42, 273–277. [Google Scholar] [CrossRef]




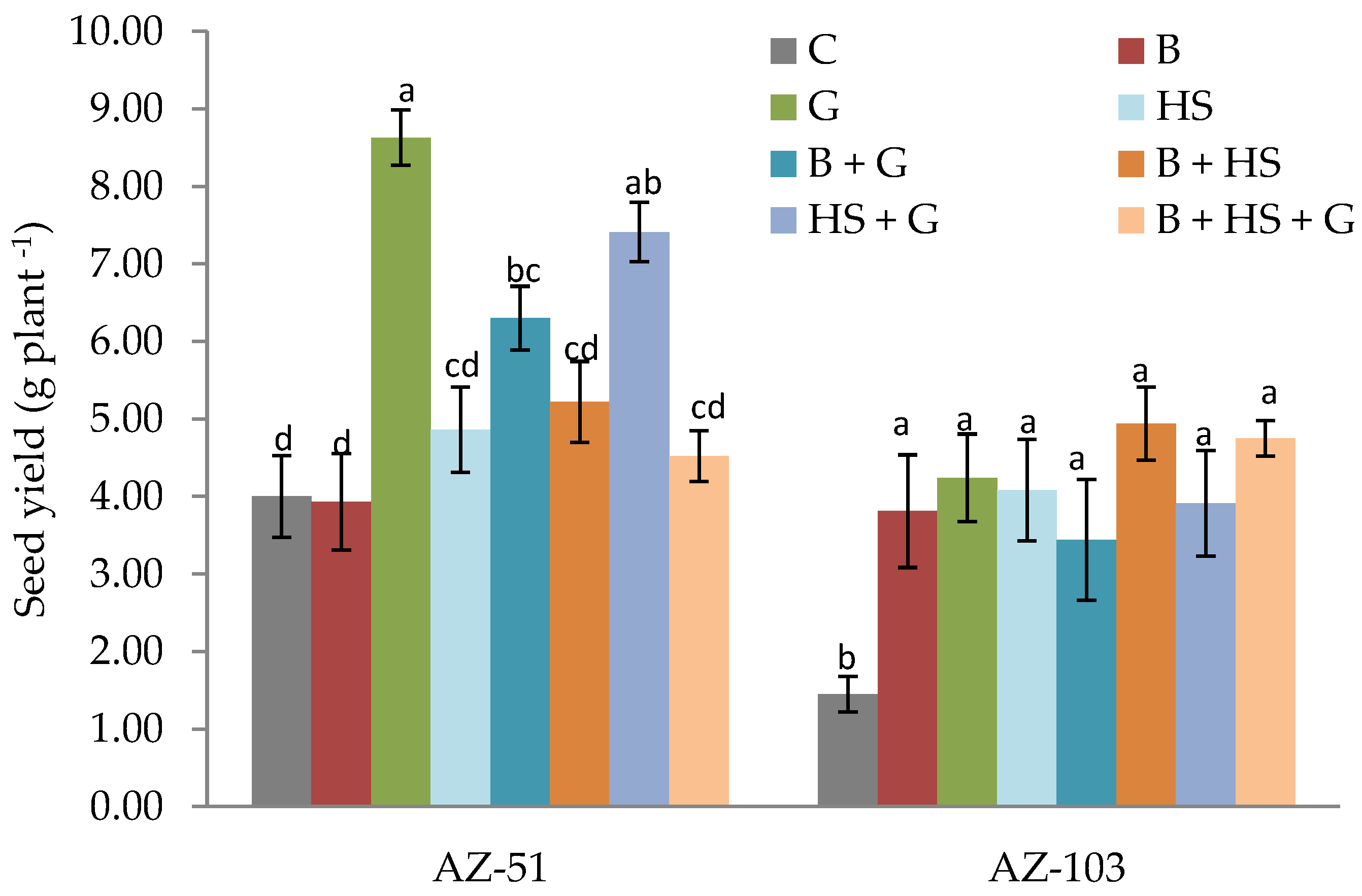{kind=link}
{kind=link}
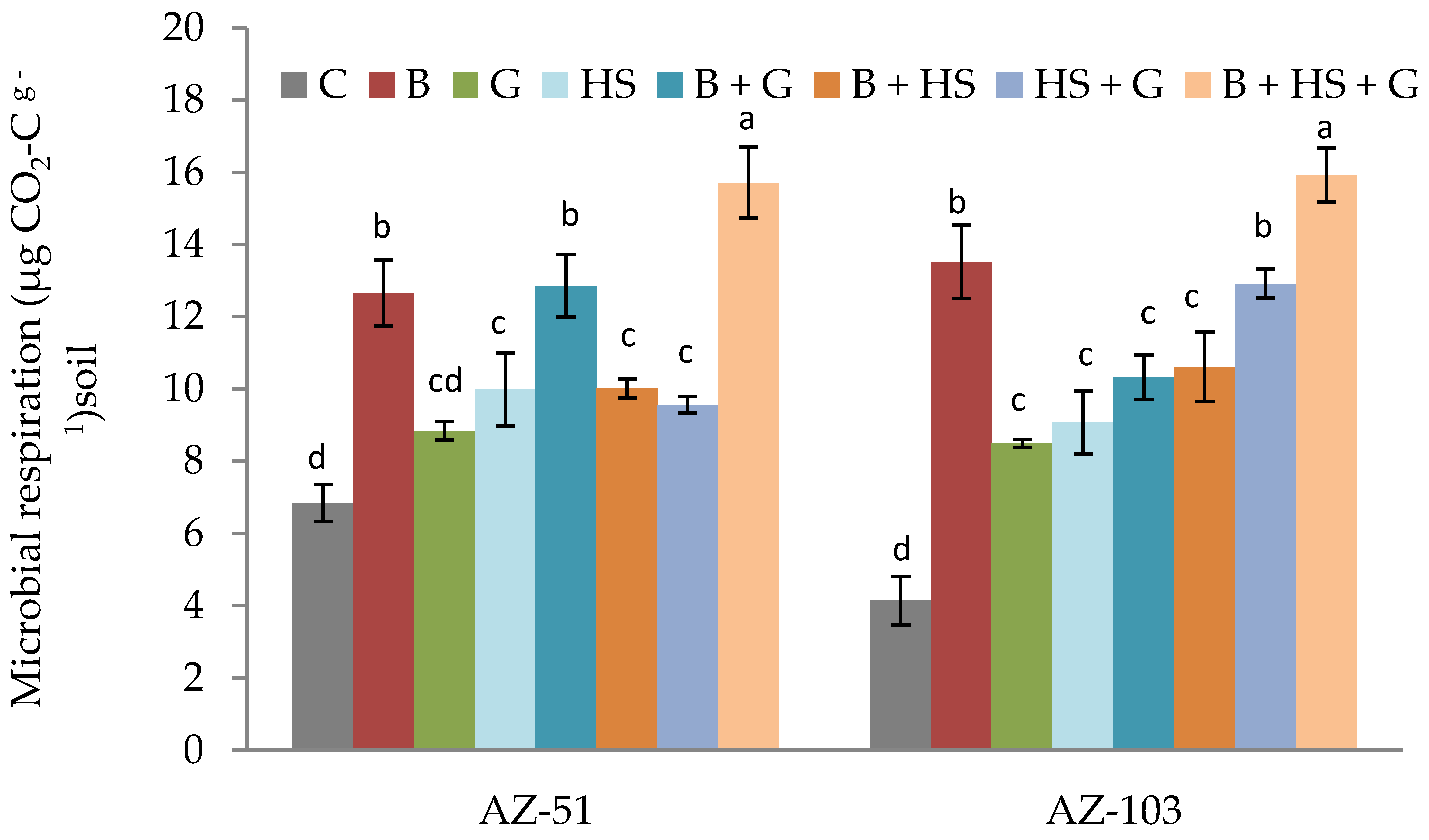{kind=link}
| Treatments | Root Dry Weight (g plant−1) | Root Length (cm) | ||
|---|---|---|---|---|
| AZ-51 | AZ-103 | AZ-51 | AZ-103 | |
| C | 0.48 ± 0.06 b | 1.13 ± 0.12 c | 415 ± 79.2 cd | 329 ± 41.8 d |
| B | 0.83 ± 0.23 b | 1.49 ± 0.06 bc | 729 ± 73.6 ab | 480 ± 31.5 abc |
| G | 0.63 ± 0.16 b | 1.39 ± 0.22 bc | 357 ± 40.1 d | 369 ± 48.3 bcd |
| HS | 0.82 ± 0.11 b | 1.63 ± 0.21 abc | 533 ± 77.1 bcd | 365 ± 23.4 cd |
| B + G | 0.81 ± 0.13 b | 1.77 ± 0.22 ab | 569 ± 61.6 bc | 511 ± 44.1 ab |
| B + HS | 0.65 ± 0.15 b | 1.95 ± 0.10 ab | 388 ± 51.2 cd | 515 ± 58.7 a |
| HS + G | 0.60 ± 0.05 b | 1.51 ± 0.24 bc | 625 ± 45.0 b | 319 ± 56.9 d |
| B + HS + G | 2.46 ± 0.27 a | 2.15 ± 0.14 a | 854 ± 75.6 a | 452 ± 44.7 abcd |
| (p-value) | <0.0001 | 0.0124 | 0.0001 | 0.0148 |
| Treatments | Chlorophyll Index SPAD (Soil Plant Analysis Development) | Stomatal Conductance (mmol m −2 s −1) | ||
|---|---|---|---|---|
| AZ-51 | AZ-103 | AZ-51 | AZ-103 | |
| C | 47.49 ± 1.20 a | 46.85 ± 1.57 d | 106 ± 10.64 c | 100 ± 4.93 d |
| B | 53.40 ± 2.17 a | 61.70 ± 1.24 ab | 116 ± 4.51 bc | 136 ± 7.04 bc |
| G | 54.37 ± 1.67 a | 62.13 ± 1.04 ab | 137 ± 6.98 ab | 147 ± 9.23 bc |
| HS | 50.33 ± 1.49 a | 54.98 ± 0.85 c | 138 ± 7.75 ab | 133 ± 7.74 c |
| B + G | 55.80 ± 2.43 a | 61.03 ± 0.66 b | 137 ± 8.63 ab | 137 ± 8.09 bc |
| B + HS | 52.31 ± 1.23 a | 59.65 ± 1.37 b | 132 ± 5.12 ab | 127 ± 8.66 c |
| HS + G | 52.13 ± 1.40 a | 65.27 ± 2.45 a | 122 ± 9.38 abc | 160 ± 5.92 ab |
| B + HS + G | 50.13 ± 2.80 a | 55.48 ± 0.37 c | 145 ± 8.39 a | 181 ± 8.78 a |
| (p-value) | 0.1028 | <0.0001 | 0.0412 | 0.0001 |
| Treatments | Polyphenols (GAE mg g−1) | Protein (%) | ||
|---|---|---|---|---|
| AZ-51 | AZ-103 | AZ-51 | AZ-103 | |
| C | 1.02 ± 0.08 a | 1.2 ± 0.07 ab | 9.3 ± 0.72 d | 11.4 ± 0.27 b |
| B | 0.75 ± 0.06 a | 1.05 ± 0.09 bc | 11.1 ± 0.61 bcd | 14.0 ± 0.53 a |
| G | 0.91 ± 0.06 a | 1.22 ± 0.14 ab | 10.4 ± 0.82 cd | 10.8 ± 0.19 b |
| HS | 0.71 ± 0.03 a | 1.02 ± 0.09 bc | 13.5 ± 0.26 a | 10.3 ± 0.48 b |
| B + G | 0.81 ± 0.16 a | 1.38 ± 0.11 a | 10.7 ± 0.76 bcd | 11.9 ± 0.37 b |
| B + HS | 0.87 ± 0.12 a | 1.21 ± 0.11 ab | 12.0 ± 0.10 abc | 11.3 ± 0.70 b |
| HS + G | 0.78 ± 0.15 a | 1.12 ± 0.09 abc | 12.5 ± 0.44 ab | 11.8 ± 0.92 b |
| B+ HS + G | 0.73 ± 0.05 a | 0.87 ± 0.08 c | 11.8 ± 0.50 abc | 11.3 ± 0.37 b |
| (p-value) | 0.2239 | 0.0137 | 0.0018 | 0.0029 |
| Treatments | ECe (dS m−1) | SAR (meq l −1)1/2 | ESP (%) | |||
|---|---|---|---|---|---|---|
| AZ-51 | AZ-103 | AZ-51 | AZ-103 | AZ-51 | AZ-103 | |
| C | 9.1 ± 0.3 a | 13.2 ± 0.4 a | 9.7 ± 0.7 a | 10.0 ± 0.3 a | 15.0 ± 1.3 a | 21.5 ± 1.2 a |
| B | 8.1 ± 0.7 a | 5.5 ± 0.8 bc | 1.4 ± 0.2 b | 2.2 ± 0.4 b | 2.2 ± 0.4 bc | 4.5 ± 0.5 b |
| G | 5.7 ± 0.4 b | 6.1 ± 0.7 b | 0.9 ± 0.0 cd | 0.9 ± 0.1 de | 1.3 ± 0.1 d | 2.1 ± 0.2 def |
| HS | 5.3 ± 0.4 b | 4.5 ± 0.3 bc | 1.3 ± 0.1 bc | 1.4 ± 0.2 cde | 2.2 ± 0.1 b | 3.7 ± 0.4 bc |
| B + G | 6.1 ± 0.8 b | 6.3 ± 0.6 b | 0.6 ± 0.0 d | 0.7 ± 0.1 e | 1.2 ± 0.1 d | 1.7 ± 0.2 f |
| B + HS | 6.1 ± 0.8 b | 4.2 ± 0.4 c | 0.8 ± 0.1 d | 1.2 ± 0.1 de | 1.4 ± 0.2 cd | 2.0 ± 0.3 ef |
| HS + G | 4.9 ± 0.2 b | 4.7 ± 0.5 bc | 1.2 ± 0.2 bc | 1.9 ± 0.2 bc | 2.1 ± 0.1 b | 4.0 ± 0.5 bcd |
| B+ HS + G | 5.3 ± 0.4 b | 4.6 ± 0.6 bc | 0.7 ± 0.0 d | 1.36 ± 0.3 cd | 1.2 ± 0.0 bcd | 2.4 ± 0.5 cde |
| (p-value) | 0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Treatments | Ca2+ (cmol kg−1) | Mg2+ (cmol kg−1) | Na+ (cmol kg−1) | |||
|---|---|---|---|---|---|---|
| AZ-51 | AZ-103 | AZ-51 | AZ-103 | AZ-51 | AZ-103 | |
| C | 52 ± 0.9 d | 50 ± 0.9 d | 8.4 ± 0.8 ab | 11.1 ± 0.7 ab | 10.9 ± 1.2 a | 16.8 ± 1.6 a |
| B | 57 ± 4.2 bcd | 57 ± 1.7 c | 7.7 ± 0.2 bc | 10.5 ± 0.6 ab | 1.40 ± 0.2 bc | 3.20 ± 0.4 b |
| G | 73 ± 2.3 a | 76 ± 1.1 a | 8.0 ± 0.3 ab | 9.5 ± 0.4 bc | 1.03 ± 0.0 bc | 1.87 ± 0.1 cde |
| HS | 54 ± 2.1 cd | 54 ± 0.6 cd | 8.4 ± 0.2 ab | 11.1 ± 0.7 ab | 1.38 ± 0.1 bc | 2.50 ± 0.3 bcd |
| B + G | 59 ± 1.9 bcd | 67 ± 2.3 b | 6.7 ± 0.2 c | 8.4 ± 0.5 c | 0.80 ± 0.1 c | 1.29 ± 0.2 e |
| B + HS | 57 ± 2.9 bcd | 77 ± 0.6 a | 8.4 ± 0.2 ab | 10.2 ± 0.3 ab | 0.95 ± 0.1 c | 1.77 ± 0.3 de |
| HS + G | 64 ± 1.8 bc | 70 ± 1.2 b | 8.9 ± 0.2 a | 11.7 ± 0.6 a | 1.59 ± 0.1 b | 3.07 ± 0.5 bc |
| B+ HS + G | 66 ± 5.7 ab | 65 ± 0.3 b | 8.2 ± 0.5 ab | 10.9 ± 0.7 ab | 0.89 ± 0.1 c | 1.94 ± 0.5 cde |
| (p-value) | 0.0011 | <0.0001 | 0.0192 | 0.0127 | <0.0001 | <0.0001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcívar, M.; Zurita-Silva, A.; Sandoval, M.; Muñoz, C.; Schoebitz, M. Reclamation of Saline–Sodic Soils with Combined Amendments: Impact on Quinoa Performance and Biological Soil Quality. Sustainability 2018, 10, 3083. https://0-doi-org.brum.beds.ac.uk/10.3390/su10093083
Alcívar M, Zurita-Silva A, Sandoval M, Muñoz C, Schoebitz M. Reclamation of Saline–Sodic Soils with Combined Amendments: Impact on Quinoa Performance and Biological Soil Quality. Sustainability. 2018; 10(9):3083. https://0-doi-org.brum.beds.ac.uk/10.3390/su10093083
Chicago/Turabian StyleAlcívar, María, Andrés Zurita-Silva, Marco Sandoval, Cristina Muñoz, and Mauricio Schoebitz. 2018. "Reclamation of Saline–Sodic Soils with Combined Amendments: Impact on Quinoa Performance and Biological Soil Quality" Sustainability 10, no. 9: 3083. https://0-doi-org.brum.beds.ac.uk/10.3390/su10093083