Agroforestry and Biodiversity


The Center for Agroforestry, School of Natural Resources, University of Missouri, Columbia, MO 65211, USA
*
Author to whom correspondence should be addressed.
Sustainability 2019, 11(10), 2879; https://0-doi-org.brum.beds.ac.uk/10.3390/su11102879
Submission received: 5 April 2019
/
Revised: 13 May 2019
/
Accepted: 16 May 2019
/
Published: 21 May 2019
(This article belongs to the Special Issue Sustainable Agroforestry Systems)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Declining biodiversity (BD) is affecting food security, agricultural sustainability, and environmental quality. Agroforestry (AF) is recognized as a possible partial solution for BD conservation and improvement. This manuscript uses published peer-reviewed manuscripts, reviews, meta-analysis, and federal and state agency documents to evaluate relationships between AF and BD and how AF can be used to conserve BD. The review revealed that floral, faunal, and soil microbial diversity were significantly greater in AF as compared to monocropping, adjacent crop lands, and within crop alleys and some forests. Among the soil organisms, arbuscular mycorrhizae fungi (AMF), bacteria, and enzyme activities were significantly greater in AF than crop and livestock practices. Agroforestry also creates spatially concentrated high-density BD near trees due to favorable soil-plant-water-microclimate conditions. The greater BD was attributed to heterogeneous vegetation, organic carbon, microclimate, soil conditions, and spatial distribution of trees. Differences in BD between AF and other management types diminished with time. Evenly distributed leaves, litter, roots, dead/live biological material, and microclimate improve soil and microclimate in adjacent crop and pasture areas as the system matures. Results of the study prove that integration of AF can improve BD in agricultural lands. Selection of site suitable tree/shrub/grass-crop combinations can be used to help address soil nutrient deficiencies or environmental conditions. Future studies with standardized management protocols may be needed for all regions to further strengthen these findings and to develop AF establishment criteria for BD conservation and agricultural sustainability.
1. Introduction
Despite numerous benefits of biodiversity (BD), the global BD is changing at an unprecedented and alarming rate [1,2]. The continued population growth increased per capita consumption and improved dietary patterns with increasing prosperity have resulted in unsustainable exploitation of Earth’s biological diversity. These trends will continue as human population is expected to reach 9.5 billion by 2050. Climate change, ocean acidification, other anthropogenic, and environmental impacts also contribute to further decline of floral and faunal richness, habitat, and BD [3,4,5,6]. Agricultural intensification and deforestation are major contributors for loss of BD, ecosystem functions, and economies depend on natural resources [7,8]. For example, domestication of crops from wild relative plants has reduced the number of plant species and varieties cultivated thus reducing BD worldwide [9]. Planting one or two crops continuously on the same land further erodes agricultural BD and impacts food security, human nutrition, and other ecosystem services [10,11].
Although modern agriculture is largely blamed for declining BD, agricultural lands can support BD provided better management plans are implemented to support their survival [12,13]. For example, in Europe ~50% of plant and animal species depend on agricultural habitats [14]. Therefore, agricultural practices that favor BD can be used to conserve and improve BD.
The widening focus from the traditional tree-based land use practice to more advanced landscape scale agroforestry (AF) approach creates a stronger link between AF and BD conservation [15]. Adoption of AF as a measure for BD conservation has received increased attention during the last two decades [15,16,17,18,19,20,21,22,23]. Agroforestry has been identified as a tool to preserve rich species diversity around the world [24,25,26] because AF plays five major roles in BD conservation [2]. These include: (1) agroforestry provides habitat for species that can tolerate a certain level of disturbance; (2) agroforestry helps preserve germplasm of sensitive species; (3) agroforestry helps reduce the rates of conversion of natural habitat by providing a more productive, sustainable alternative to traditional agricultural systems that may involve clearing natural habitats; (4) agroforestry provides connectivity by creating corridors between habitat remnants which may support the integrity of these remnants and the conservation of area-sensitive floral and faunal species; and (5) agroforestry helps conserve biological diversity by providing other ecosystem services such as erosion control and water recharge, thereby preventing the degradation and loss of surrounding habitat. Likewise, AF can be used as a tool in conjunction with appropriate conservation practices to buffer BD loss because some AF practices have 50–80% of the diversity of comparable natural forests and this can contribute to further preservation of BD.
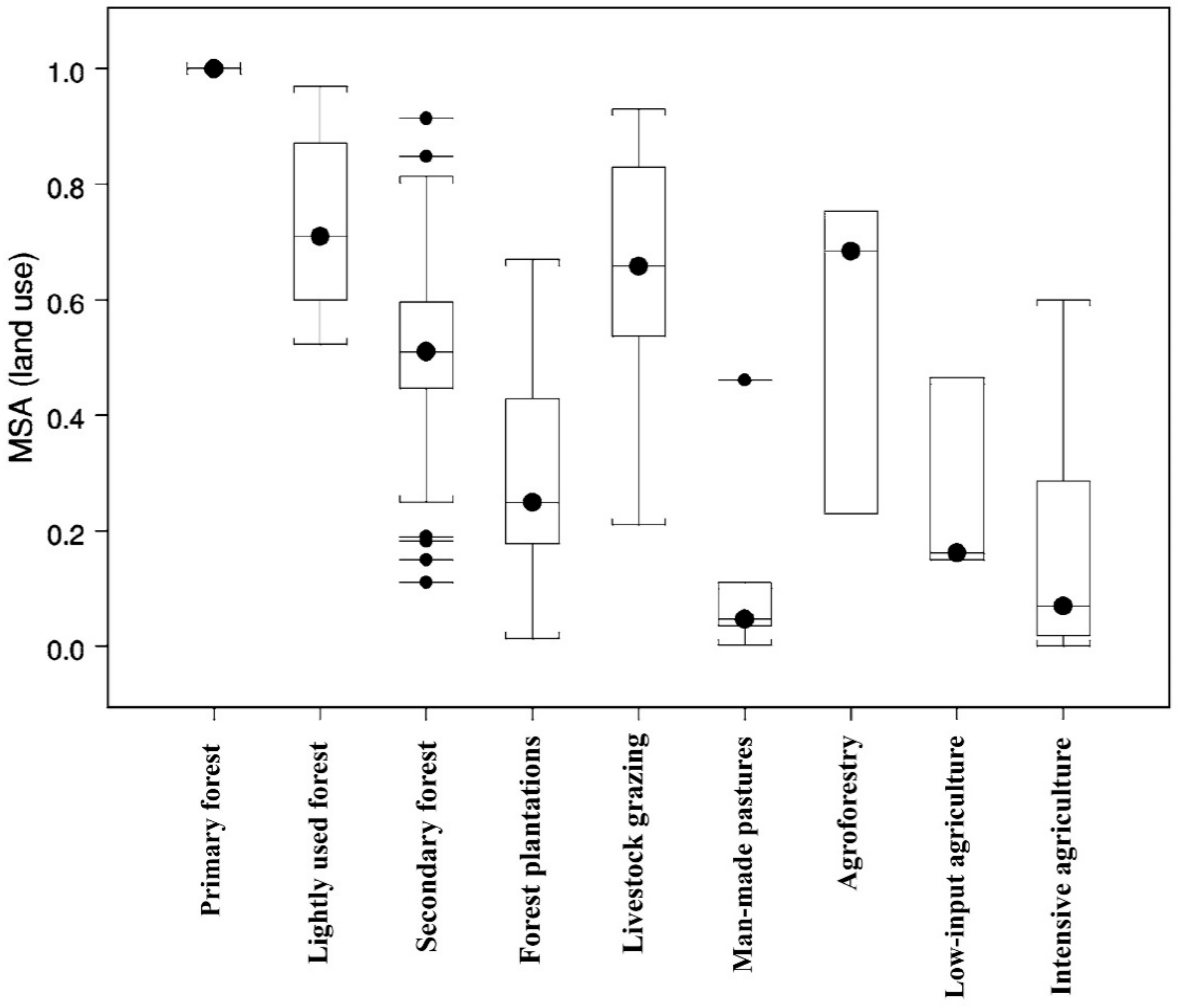
Agroforestry induced BD improvements have been reported in both temperate and tropical regions [22,27,28,29]. Some studies have indicated significantly greater diversity in AF compared to forests and tree monoculture management (Figure 1) [7,11,22]. In a meta-analysis Bhagwat et al. [30] reported 60% greater mean richness of taxa in AF as compared to forests. Another meta-analysis in Europe showed an overall positive effect of AF on BD [31]. Conversion of AF to monocultures has reduced BD [32,33,34]. In spite of greater BD in AF as compared to adjacent forests and agricultural systems, AF usually have lower number of endemic species due to intensive management [27,30].
Recently, studies have indicated a strong connection between BD and ecosystem services and strongly emphasize the importance of BD conservation for enhanced ecosystem services [4,35,36]. Biodiversity provides many essential services to the society including material and non-material benefits and regulating environmental functions [4,37]. Biodiversity improves human physical and mental health [38], positively contributes to agriculture via pest control and pollination, and provides long-term resilience to disturbances and environmental changes [35], substantially contribute to economic and social development [36], provides valuable ecosystem services and functions for agricultural production at genetic, species, and farming system levels [10]. Loss of BD reduces ecosystem functions, crop yields, and income from wildlife and plants while increasing health risks and malnutrition [39].
A significant number of studies have identified positive relationships between AF and ecosystem services of AF [2,7,31]. Agroforestry can increase land productivity as combinations of trees with crops and pasture could utilize resources more efficiently (solar radiation, water, soil volume etc.) than mono cropping or trees [31,40]. Agroforestry improves regulatory ecosystem services including erosion control, nutrient retention, water quality, soil health, air quality, pollination, pest control, and fire retardation as well as cultural services such as improvements in recreational, aesthetic, and cultural values [31,41,42,43]. Agroforestry has been recognized as a tool for carbon sequestration in the Kyoto Protocol for mitigation of the climate change. These benefits can be related to BD of AF. For example, multi-species riparian buffers with trees, shrubs, and grasses improve water and soil quality by retaining soil and nutrients on the land and reducing losses to water bodies.
Although AF enhances BD, the role of AF for BD is misunderstood and understated [44]. It is therefore, imperative to appraise multiple links between AF and BD, and how AF can be used to improve BD. This review addresses AF’s potential for reversing the negative impacts of domesticating crops and adoption of monoculture systems, and the role of AF with the hypothesis that AF practices increase the species variability and species richness thus help conserve the global BD and possibly contributing to enhance ecosystem services.
2. Materials and Methods
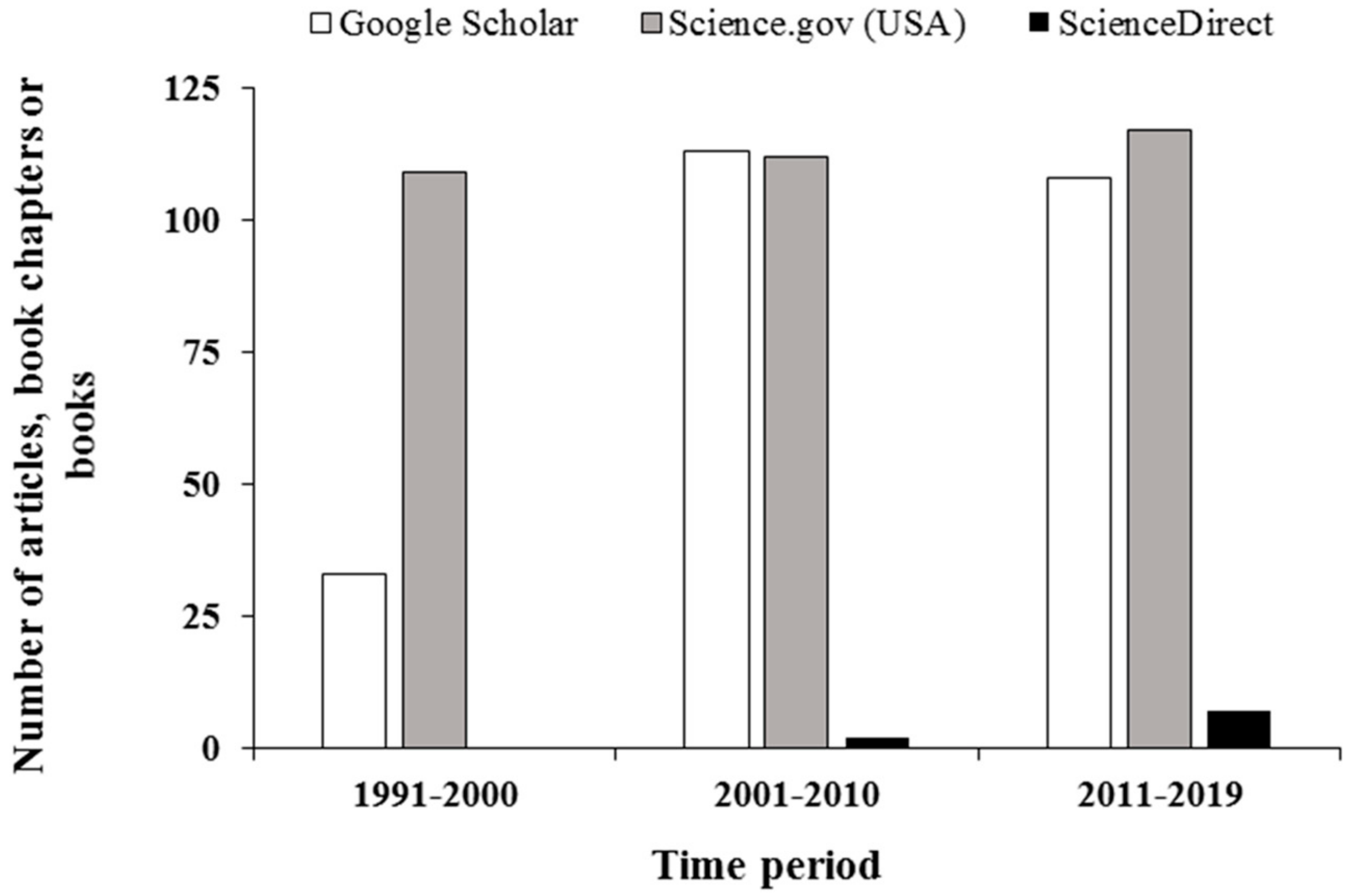
A literature search was conducted on the Google Scholar, Science.gov (USA), and ScienceDirect, using ‘agroforestry biodiversity’ as keywords for three time periods (Figure 2). The Google Scholar showed a marked increase in publications from ~33 during 1991–2000 to over 110 after 2000. Science.gov showed over 110 publications during the entire search period. The number of publications found using ScienceDirect was <5 during the two initial search periods. During the 2011–2019 period, the search located 17 publications. We have used selected publications from these search activities and other related articles to develop the review.
These selected manuscripts or reviews were published in peer-reviewed journals or books and consisted of experimental designs with replications and controls. Among these publications, about 75% of the results and data were on floral and soil BD comparisons with monoculture management practices with either crops or forestry. Only a few publications were available on faunal BD and AF. Although the number of publications are limited for the temperate region, more and more publications have surfaced during the last 20 years.
3. Agroforestry and Diversity
3.1. Agroforestry and Floral Diversity
Modern agricultural farming methods focus on providing just one ecosystem service: food production, which is achieved by reducing environmental complexity and growing large areas of monocultures for better economies of scale [45]. This lower species diversity can lead to lower functional diversity, which eventually results in reduced ecosystem functions [46]. In contrast, integration of AF into farming systems increase BD [45], as AF can harbor a much higher species richness and diversity compared to monoculture cropping systems [2]. Numerous studies and reviews from tropical and temperate regions have reported increased plant diversity due to AF (Figure 3).
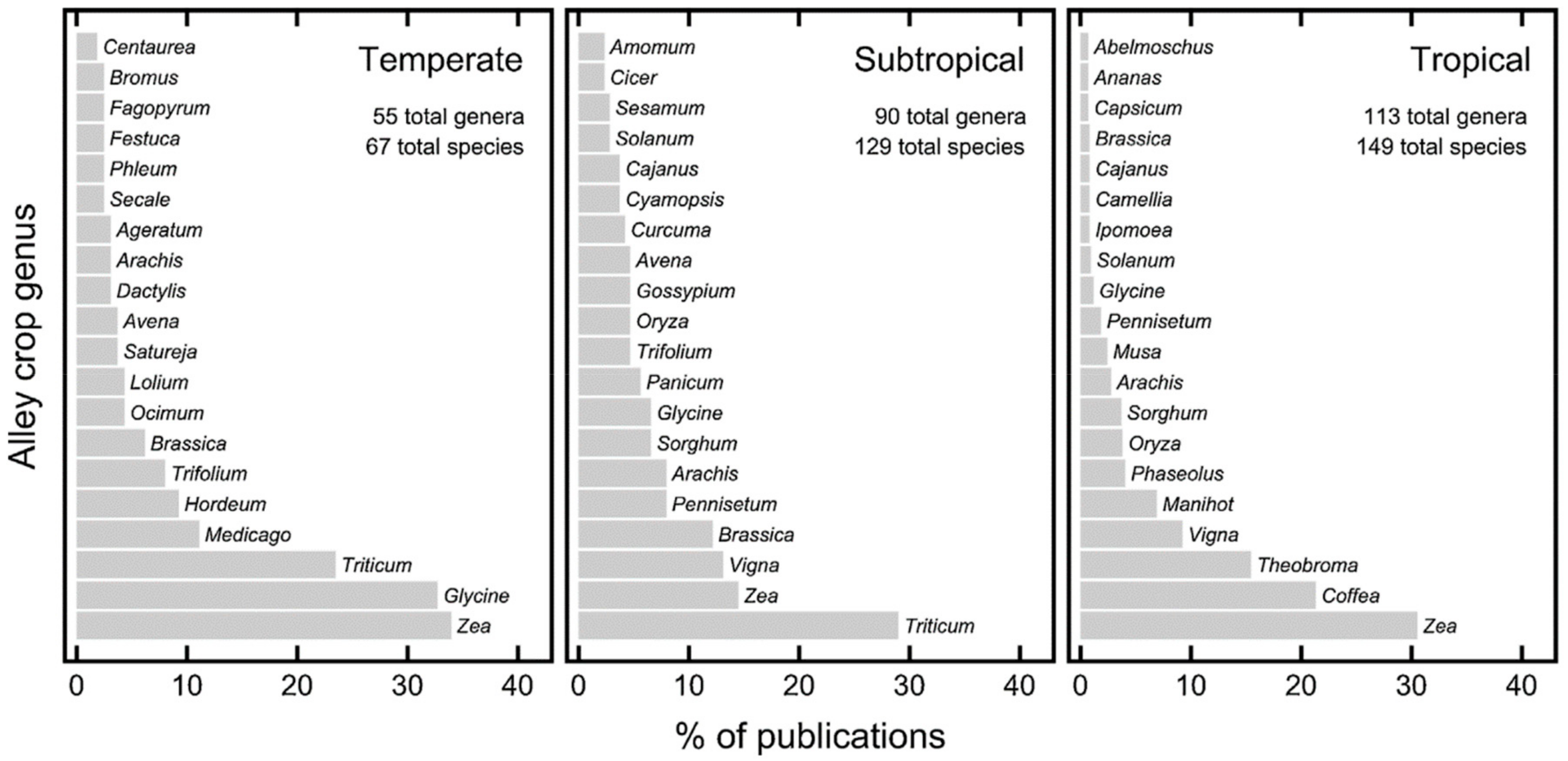
Alley cropping, forest farming, riparian buffers, silvopasture, and windbreaks are the five main AF practices in North America. In the temperate zone the most commonly used alley cropping trees were dominated by Juglans (walnut) or Populus (poplar) genera and those were found in 55% of the publications used for a review on floral diversity of AF [47]. Pecan (Carya illinoensis (Wanganh.) K. Koch), walnut (Juglan Negra), chesnut (Castanea spp.), oaks (Quercus spp.), ashes (Fraxinus spp.), and basswood (Tilia spp.) can be used in alley cropping [48]. Forage crops, cereal, vegetable, specialty crops, and biomass crops can be grown in alley until light, water, and nutrient become limiting.
In forest farming for edible products such as fruits, nuts, berries, greens, mushrooms, and wild vegetable; medicines and herbal supplements such as ginseng, echinacea, goldenseal, black cohosh, and witch hazel; decorative products such as flowers, Spanish moss, vines, stems, seedheads, leaves, and fruiting structures used in floral arrangements; handicrafts and specialty woods such as grape vines and branches are also commonly used [49].
Riparian buffers also contribute to greater diversity. In Appalachian headwater catchments, herbaceous species density varied from 4 to 17 m−2 and woody species density varied from 3 to 8 m−2, in a four years study [50]. The diversity in riparian corridors vary along longitudinal, lateral, and vertical dimensions [51].
Silvopasture is the most prevalent AF practice found in USA and Canada [52,53]. Both hardwood and evergreens are used with warm and cold season grass species. In addition, N-fixing species are also integrated within silvopasture. Many tree species found in forests and trees planted within pastures can increase floral diversity in silvopasture.
Tree planting for windbreaks has been promoted in USA and Canada and these systems created some form of AF in both countries. The Timber Culture Act of 1873 provided the title to the land if a homesteader would plant trees on a 10–40 acre until the offer was repealed in 1891. In 1930′s Prairie States Forestry Project was implemented to establish windbreaks to protect soils from wind erosion after the Dust bowl. About 217 million trees were planted in a 1150- by 100-mile zone from Canada border to Texas in six states. Windbreaks established in the US for farmstead use, field use, livestock shelter, living snow fence, wildlife habitat, and screening usually consist of several tree species including a combination of evergreens and deciduous as well as shrubs. Commonly used species include Arizona cypress (Cupressus arizonica), Austrian pine (Pinus nigra), Bur oak (Quercus macrocarpa), Eastern red cedar (Juniperus virginiana L.), Green ash (Fraxinus pennsylvanica), Ponderosa pine (Pinus ponderosa), Poplar (Populus tremula L.), Redbud (Cercis canadensis L.), Sycamore (Platanus occidentalis L.), Walnut (Juglans regia), White ash (Fraxinus americana L.), Willow species (Salix alba L.), and White spruce (Picea glauca). The Prairie Farm Rehabilitation Administrative under Agriculture and Agrifood Canada has initiated windbreaks and planted >610 million shelterbelt trees in Canada. Agriculture Canada has estimated >218 mega tons of C and over $600 million worth other goods and services from these shelterbelts.
The selection of these species for each practice varied by regions, climate conditions, and plant hardiness [54,55,56,57]. Biomass crops, specialty crops, N-fixing species, and medicinal plants can be incorporated in alley cropping, windbreaks, and riparian buffers for additional income and wildlife habitat.
Several studies in Central and South America also have reported increased diversity with AF integration [11,23,57]. In a plant species diversity study in Nicaragua Sistla et al. [11] reported that AF and secondary forests shared 38 species, secondary forest and pasture shared 15 species, AF and pasture shared 28 species. In their study, species richness per m2 was 8.14 ± 0.15, 7.97 ± 0.15 and 6.18 ± 0.6 for secondary forest, AF, and pasture. Additionally, secondary forest and AF had greater food and medicinal value than the pasture.
The species richness of tropical home gardens varied from 27 (Sri Lanka) to 602 (West Java) [58]. In another study using 402 home gardens from six regions, Kabir and Webb [59] found that 59% species were native out of 419 across southwestern Bangladesh. In a study conducted to identify plant diversity and multi-use, Kumari et al. [60] evaluated randomly selected 106 suburban home gardens in Sri Lanka and reported a total of 289 species of which 51% were ornamental plants, 36% were food plants and 12% were medicinal plants and 6% generated income whereas the rest was used for domestic usage.
According to Bardhan et al. [61], home garden AF can function as an “intermediary” for conserving tree species diversity in Bangladesh. Home gardens and natural forests shared 30% of the species and the species richness in home gardens increased as the size of home gardens increased. In Jambi area in central Sumatra, Murdiyarso et al. [62] observed 15 species per 1.5-ha plot in continuously cultivated cassava, 25 species per plot in oil palm plantations, 90 species per plot in rubber AF, and 120 species per plot in primary forests. Neo tropical and old-world tropical home gardens are considered as AF systems with high floristic diversity [63] and many ecologists consider such systems mimic the natural forests to the closest level, both structurally and functionally [64].
Diversity of tree species in AF were much greater than previously reported in Cameron, Kenya, and Uganda [65]. Greater level of tree diversity has been reported in complex and shaded AF systems in West Africa, Central America, and Asia [66,67,68,69]. In Cameroon and Nigeria, cocoa AF play a major role in their agriculture than monocrop cocoa. In Nigeria, tree species including Cola nitida, Persea Americana, Mangifera indica, Citrus sisnensis, Elaeis guinensis, Millicia excelsa, Irvingia gabonensis, Terminalia superba, Garcinia kola, Triplochiton scleroxylon, Dacryodes edulis, Khaya ivorensis, and Cola acuminate are generally associated with cocoa [70,71]. In Ghana, Rauvolfia vomitoria, Milicia excelsa, Sterculia tragacantha, Alstonia boonei, Milicia excelsa, Terminalia ivorensis, T. superba, Triplochiton scleroxylon, Ceiba pentandra, Pycnanthus angolensis, Entandrophragma angolense, Funtumia elastica, Ricinodendron heudelotti, Tetrapleura tetraptera, Citrus sinensis, Persea americana, Mangifera indica, and Elaeis guinensis are commonly found on cocoa farms [71,72].
Studying plant diversity in Southeast Asia, the Congo Basin, and the Amazon basin, Tomich et al. [73,74] stated that diversity of multistrata AF systems was between primary forests and monocrop perennials or field crops. A review using 89 published data sets, Alkemade et al. [37] showed that AF had diversity similar to lightly used forests, secondary forests, and forest plantations (Figure 1). The same review showed that primary forest contained greater diversity and agriculture contained lower diversity than AF. Another review evaluated global floral diversity in alley cropping AF and found that alley cropping used 410 tree species from 192 genera [47]. Globally, 276 species from 181 genera were found in alleys, but were dominated by a few species (Figure 4). Findings from studies across the world suggest that AF has greater diversity as compared to monocropping practices and perhaps greater than forests in some regions.
3.2. Agroforestry and Faunal Diversity
AF systems with higher floristic and structural diversity have been shown to support greater faunal diversity compared to monoculture systems because the faunal diversity is closely linked with floral diversity [2]. Agroforestry induced faunal diversity in small and large farms as well as for temperate and tropical regions have shown many direct and indirect benefit on sustainability, land productivity, and environmental services. For instance, AF can be used to increase pollinator diversity, which is essential for food production as well as maintenance of population levels of wild plants [45]. The pollinator service is invaluable as ~90% of flowering plants are pollinated by insects and over 75% of world’s most important crops and 35% of food production depend on animal pollination [75,76].
The Nature Conservancy and the Americas Bird Conservation Program commenced an initiation in 1997 to enhance the environmental conservation of a biological corridor connecting the Talamanca region in Costa Rica using shaded cocoa (Theobroma cacao) as an AF system [77]. Shade coffee and multistrata cocoa AF systems provide habitat for avian, mammalian, and other species and thereby enhance faunal diversity [2]. Gibbs et al. [78] observed an almost doubled (increased from 17 to 32 species) avian species richness between 1995 and 2014 in a tree-based intercropping system in Canada. Their study also reported the highest species richness of birds in the tree-based system among studied systems for both time periods. Another study in southern Canada also found greater bird species richness in tree-based systems compared to adjacent soybean fields [79]. Agroforestry systems favor greater diversity due to greater species richness [80].
A review by Buck et al. [23] showed greater bird diversity in cocoa and coffee AF in Southeast Asia and Central America as compared to monocrop. Tree density and the distance to intact forest had the greatest impact on number of forest bird species in intact primary forest, regenerated secondary forest, and agricultural fields in and around Mabira forest in Uganda [81]. Studying bird diversity in Chagga home gardens of Kilimanjaro, Tanzania, Soini [82] reported that the diversity and richness were determined by the size of the land parcel and human pressure. Assessing bird biodiversity during the fires of 1998 in two AF farms in the buffer zone of the Maya Biosphere Reserve in Guatemala, Griffith [83] found high numbers of bird species in AF areas, including forest specialists and forest generalists. These sites provided fruit, nectar, nesting sites, protection from predators, and refuge both in AF and forestry patches.
The integration of trees provides connectivity, nesting sites, protection against predators, low risk areas, breeding areas, food sources, landscape complexity, and heterogeneity, and thereby integrating aquatic systems, pollinators, and beneficial species into the landscape. For example, in Makalu Barun National Park and Conservation Area of Nepal, AF with Alnus nepalensis and cardamom (Elettaria cardamomum) have contributed to integrity of riparian corridors for wildlife conservation [84]. Only windbreaks and riparian buffers offer woody habitat for wildlife in many agricultures dominated and monocropping landscapes thus leading to improved wild life habitats [85] and species richness of birds [86].
As compared to forests, the pollinator decrease was 40% under monocrops and 15% under mixed-tree AF [87]. Furthermore, the authors [87] have noticed a 15% increase of less important pollinators in the mixed-tree AF and no increase in the monocrop. The mixed-tree AF had 93% of the pollinators found in forests and maintained 85% of the pollinators found in forests. Increased vegetation cover provided by perennial vegetation, microclimate, flowers, and nesting sites, associated with diversity of plants have been identified as major contributors for increased pollinator BD [88,89,90,91,92]. According to Barroios et al. [87], the multiple tree species were more beneficial than a single tree species for greater pollinator BD. Insects in AF also provides indirect benefits such as pest and disease control. In West Africa, coffee berry borer infestation was 69% lower in AF coffee practices due to natural enemies of ants, parasitiods, [32] and birds [91], as compared to monocrops [87].
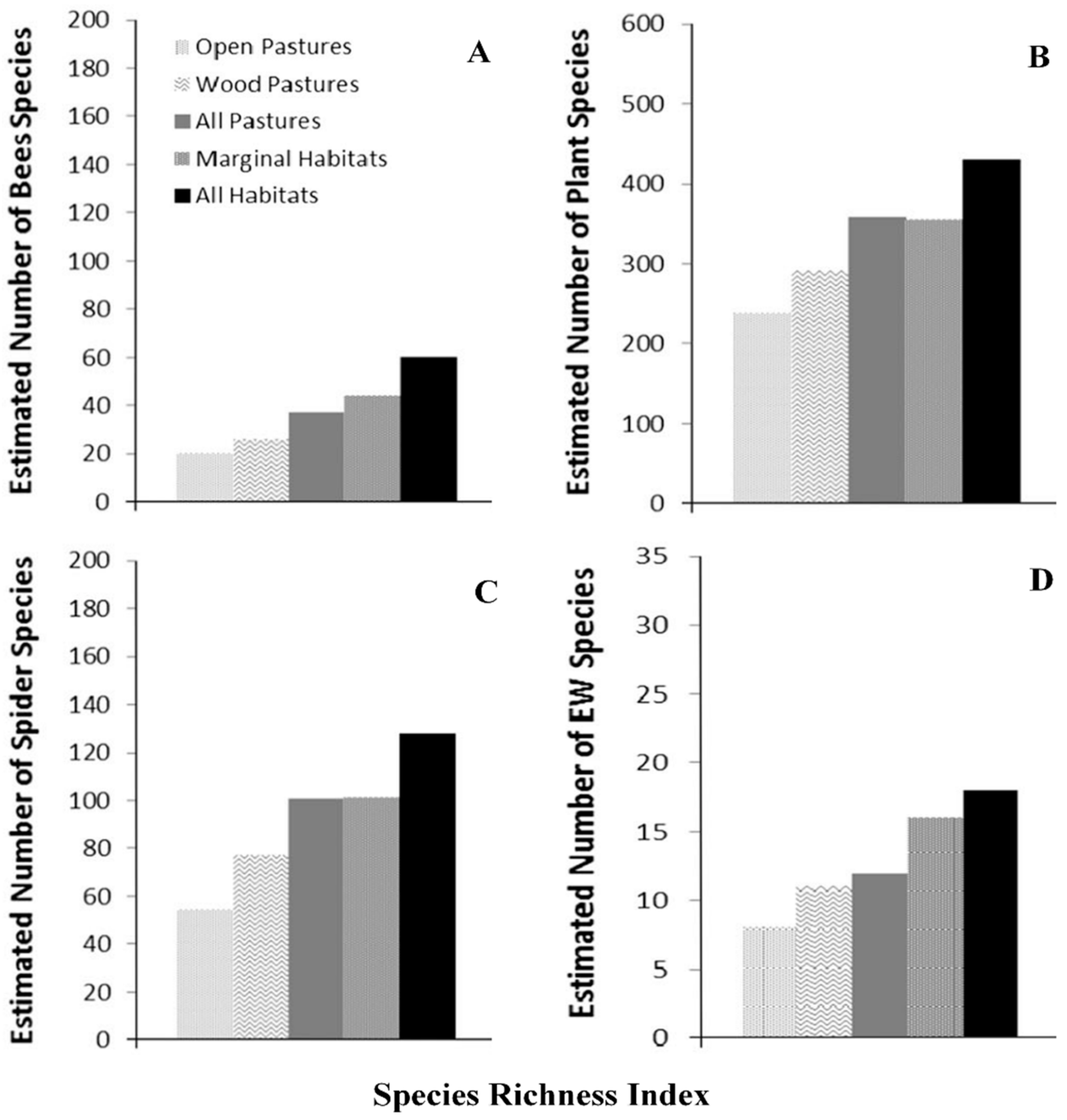
Various tree-crop combinations and spatial arrangements in AF systems have been reported to affect insect population density and species diversity [63]. Greater density and diversity of insect populations have been reported in windbreaks and tree areas within pastures than monocrop areas Figure 5; [92,93]. In Iberian Dehesas, bees and spider species richness were greater with wooded pastures than open pastures [93]. The insect species richness increased with the increasing plant species richness.
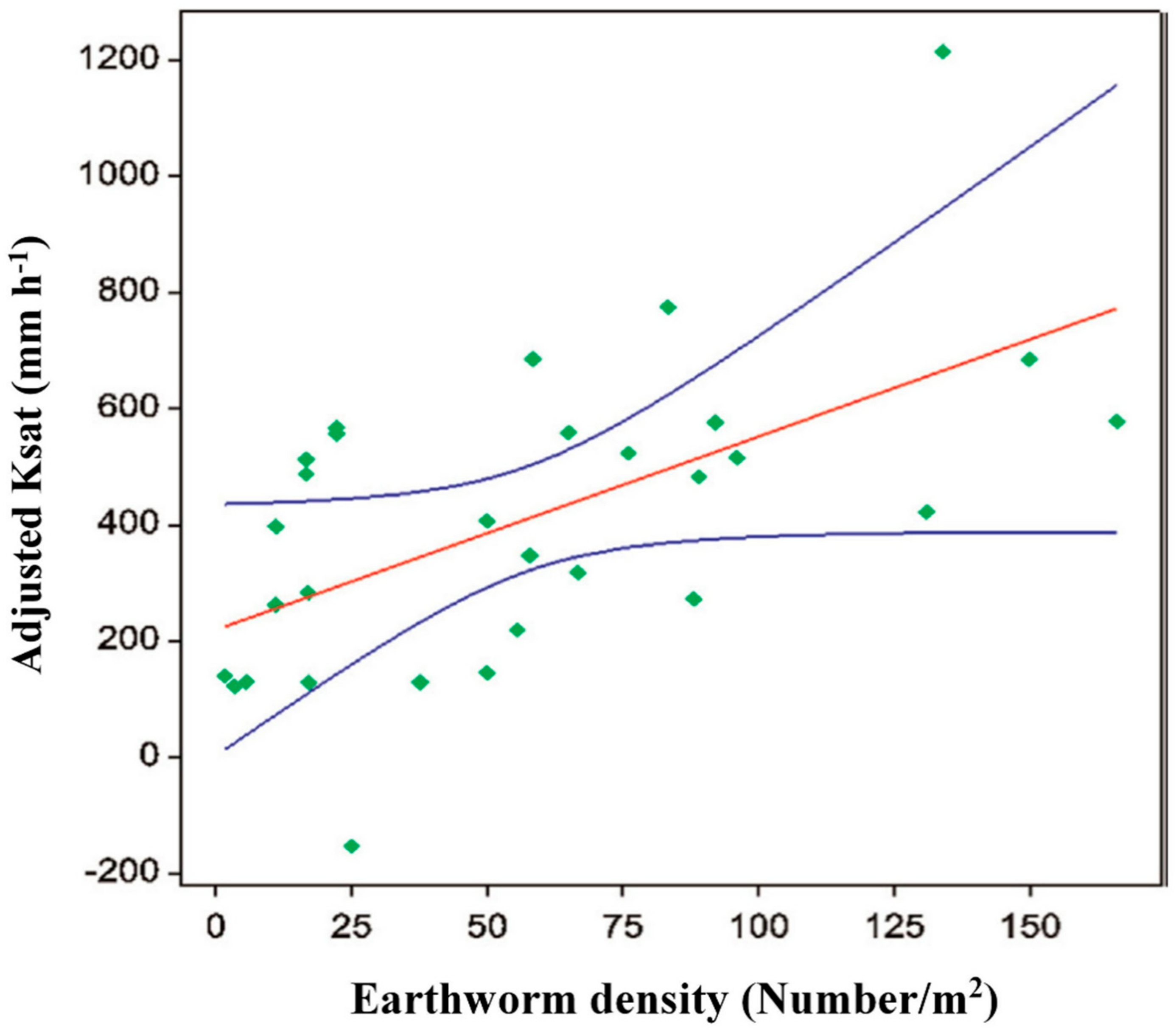
In southern Ontario, Canada, earthworm populations were greater in AF as compared to conventional cropping systems and the population density was greater near the trees [94]. The density ranged from 71 m−2 in silver maple (Acer saccharinum L.) to 90 m−2 for white ash (Fraxinus Americana L.) to 182 m−2 for poplar (Populus spp.). In the extensive silvopastural systems of Iberian dehesas, earthworm species numbers and richness were greater for wooded pastures than open pastures [93]. Another study in Canada showed greater population numbers of arthropods and detritivores in AF than conventional cropping [28]. The spatial distribution of earthworm casts was closely related to the distribution of trees Figure 6 [95], indicating the favorable conditions provided by trees and spatially heterogeneous soil properties even within a small area.
These larger organisms like birds, earth worms, and insects also help reduce parasites and improve soil properties and soil fertility (Figure 6 and Figure 7). A survey of 13 AF practices in France showed that soil tillage and inorganic fertilization significantly increased total earthworm abundance and biomass in tree rows due to increased soil organic carbon and lack of disturbance relative to alleys and treeless control plots [96]. These studies emphasize the importance of strategies planning of trees and more than one species of trees/shrubs for maximum BD and other benefits.
In a review of BD in tropical AF, Bhadwat et al. [30] found 112–139% richness of bats and lower plants (bryophytes and ferns) than the neighboring forests. The AF showed 91% of mammals, 98% of insects, and 100% of trees found in the forest. Current literature supports that integration of AF helps improve faunal diversity, although literature is limited for selected individual species. Only a few studies have reported relationships between BD for medium to small animals. The smaller parcel size and lack of research studies may have contributed to lack of data on BD and AF for medium to small size fauna.
3.3. Agroforestry and Soil Microbial Diversity
A large portion of Earth’s BD resides belowground; one m2 may contain ~20,000 species of fungi, bacteria, virus, protozoa, nematodes, and many other species [98]. Soil microbial communities play a vital role in most biogeochemical processes and they are important for biogeochemical cycles, nutrient cycling, mineralization, and nutrient supply, degradation of chemicals, aboveground BD, soil formation, and soil health. These soil microbial communities live within the soil, rhizosphere, and other materials to carryout various processes that can even help decontaminate soils and control of plant, animal, and human pests and diseases. Their functions also help improve the quality of water and soil and climate regulation.
Differences in fungi densities and community structures have been found between AF and monocropping practices for temperate and tropical regions. A study at the University of Guelph Research Station, Canada has shown greater arbuscular mycorrhizal fungi (AMF) richness in a tree-based cropping system and several taxa that were not present in a monocrop system [99]. In their study, AMF composition was significantly different between the monocrop and tree-based cropping systems. AMF communities growing on corn roots were influenced by the tree species and communities were significantly different for Norway spruce and silver maple.
In Germany, a greater abundance of saprotrophic fungi and ectomycorrhizal fungi, as well as a greater fungalC:bacterialC ratio were found in tree rows of the silvopasture [100]. In Canada, phospholipid fatty acid (PLFA) measured fungal abundance was greater in a tree-based system compared to adjacent cropping systems [101]. In another study, Chifflot et al. [102] observed significantly greater spatial differences and diversity of fungi pores on tree based intercropping sites compared to a monoculture. As the system matures shifting towards saprotrophic and ectomycorrhizal fungi occurs and this shift was attributed to more complex and diverse organic material, reduced soil disturbance, and soil pore geometry [100]. Studying three AF systems using molecular techniques, Zhang et al. [103] mentioned that fungal diversity was greater in the rhizosphere compared to bulk soil, but differences were not significant among systems.
Another Canadian study using phospholipid fatty acid (PLFA) profiling found significantly greater AMF in a tree-based AF system compared to an adjacent conventional monocropping system [101]. A long-term fruit tree AF practice with perennial native vegetation also increased AMF compared to tree fruit and single-species alleys of tall fescue [104]. Using molecular analysis, Tomich et al. [73,74] found greater richness in AMF communities with the tree-based cropping systems in Southeast Asia, the Congo Basin, and the Amazon basin. Significantly greater fungal diversity and spores near trees of tree-based alley cropping than monocrop and forests have been noticed for many regions [99,102,104]. According to a review by Bainard et al. [99], integration of AF increases fungal diversity and abundance than monocrop systems.
Numerous benefits can occur as AMF can increase soil structure, nutrient status, microbial community structure, and suppress weed populations [99]. This in turn will help enhance soil microbial components (bacteria and protozoa) within AF systems. Because of the high density of roots of multiple vegetation including trees, shrubs, grass, and crops as well as contributions from livestock, inevitable belowground interactions occur within the soil [40,63]. Researchers have found a higher level of root colonization and greater spore densities in the rhizosphere of crops growing in close proximity to trees than away from trees [1,105,106].
Greater bacterial abundance and species richness have been observed in AF as compared to monocrop in Canada [107]. They have concluded that soil carbon and pH were the major determinants of bacterial communities. In an AF alley-cropping with fruit trees with alleys of native grasses and forbs in Missouri, Kremer and Hezel [108] observed higher total soil microbial biomass and more robust microbial community compositions especially more gram-negative bacteria in AF compared with alleys of tall fescue and adjacent unmanaged pasture and row-cropped fields.
A study using 16S rRNA gene copies, Banarjee et al. [107] found significantly greater bacterial abundance and species richness in hedgerows and woodlands compared to agricultural lands across a 270-km soil climate gradient in Alberta, Canada. Similarly, Zak et al. [109] observed an increasing microbial abundance with an increasing number of plants. Field and laboratory studies show these diverse groups are more resilient due to diversity of the litter and rhizodeposition products of trees [110,111]. Due to plant diversity and favorable conditions higher microbial biomass and N mineralization rates are reported in temperate windbreaks compared to conventional agriculture [112,113]. These benefits have been attributed to quantity and quality of litter, complex organic compounds, rhizodeposition products, and soil physio-chemical properties, aggregate stability, thermal parameters, microclimate, and absence of disturbance [99,114,115,116,117,118,119,120].
The spatial distribution of crops, pasture, shrubs, and trees in AF is different from monocrop management and these factors influence quality and quantity of organic matter, soil properties, and microclimate thus influencing soil biological activities such as enzymes [63,107]. Plot and field studies have shown that AF relatively has diverse and greater microbial communities and functions than monoculture crop and grazing areas [99,107,116,121,122,123,124]. Individual findings as well as meta-analysis have reported greater mean richness of taxa in AF as compared to forests [30]. The meta-analysis estimated 60% greater richness in AF than cropping systems. In alley cropping AF practices, soil enzymes were significantly higher in both grass and grass plus tree strips than in continuously cropped alleys [116,118,125]. On the same watershed, Weerasekera et al. [126] noticed significantly greater enzyme activities between AF and conventional crop areas 10 years after the establishment of buffers. Paudel et al. [123,124] observed greater enzyme activities in a silvopatsure site with eastern cottonwood trees (Populus deltoides) plus tall fescue grass (Festuca arundinacea) buffers, tall fescue grass buffers, and permanent pasture alleys of tall fescue plus forage legumes, compared with a row-cropping (corn-soybean rotation) system. Similar findings of greater enzyme activities can be found in the literature for AF cropping systems and silvopasture from the temperate and tropical regions [104,116]. Enzymes degrade and convert complex molecules to simple molecules and also synthesize compounds and molecules for their needs thus greater enzyme activities indicate enhanced potential to degrade cellulose, hemicellulose, chitin, peptidoglycan and proteins which leads to improved mineralization and nutrient cycling.
The influence of AF on microbial parameters are inconsistent [96,116,118,123,124,127,128]. Greater biological activity and microbiological properties, decreasing differences with time, or no difference between AF and monocrop managements also have been reported in the literature. Bambrick et al. [127] found no significant differences in C and microbiological properties between 4-, 8-, and 21-year old tree-based oat (Avena sativa L.)-maize-maize rotational system in Canada. At the Greenley paired watersheds, Weerasekera et al. [126] also observed no differences in enzyme activities at 30-, 90-, and 150-cm from the tree base. In Germany, Nii-Annang et al. [129] found no significant difference for microbial indicators at 0–3 and 3–10 cm soil depths nine years after tree establishment. These findings may imply uniform influence of AF as a system matures such as better distribution of leave, branches, litter, roots, and other improvements in soil-plant-water relationships thus diminishing differences between AF and cropping systems. For example, Mungai et al. [116] and Bardhan et al. [128] reported no difference between silver maple (Acer saccharinum) tree rows and alley cropped soils for a 21-year old AF system at two different time intervals since establishment.
4. Practical Implication of Agroforestry on Biodiversity
Since crop land and pasture lands represent 40% of the global terrestrial land area [130] and more than 43% of the global crop lands contain at least 10% tree cover [84], AF has a greater potential to conserve and improve BD. Agroforestry can be used for BD conservation as AF naturally has significantly greater BD compared with monocrop management practices [28,31,61,131,132]. The plant diversity is one of the main reasons for increased BD in AF. For example, AF in tropics resemble natural forests and therefore those have been suggested as promising wildlife friendly practices [7]. Adoption of AF with diverse trees/shrubs/grasses and crops may mimic natural forest conditions and may enhance BD while providing additional income and aesthetic value. This also helps preserve natural forests as firewood, bee honey, nuts, and other products will satisfy daily needs. There are several approaches that AF can implement for BD conservation and improvements.
Recent reviews have highlighted that AF contributes to conservation and protection of BD [23,133] and abundant benefits arising from the BD conservation. Research shows that diverse aboveground vegetation and their dynamics strongly influence BD of the system and the interactions between the above- and below-ground determines below ground communities. For example, alley cropping and silvopasture have better soil properties and thus improved soil porosity, water dynamics, and nutrient cycling efficiency, better plant growth, as well as minimum leaching losses compared to monocropping practices of row crops or pasture alone management [11,118,123,134,135]. Removal of shade trees increase the surface soil temperature and thereby affects humidity-moisture conditions, decomposition rates, nutrient status, and soil communities. In Missouri, AF buffers have shown better soil thermal properties and better buffering against extreme temperature conditions as compared to monocrop areas [120]. Microclimate properties were also different between crop and buffer areas with favorable soil temperature, humidity, and wind speeds were found in the buffers than the crop areas [136]. Studies from other parts of the world also have shown favorable soil properties, microclimate, and nutrient status and thus supporting greater diversity of AF management practices. These findings imply that selected species combinations at proper spacing and configurations can help improve and conserve BD through creating favorable microclimate and soil conditions.
Diverse vegetation and/or livestock provides a more complex system that can support a wide range of organisms. For example, leguminous trees and shrubs can affect the BD in unique ways although the number of species are limited in the temperate regions relative to the tropics. According to a recent study [137], plant diversity is positively correlated with microbial beta diversity which indicates greater microbial diversity with greater species communities [107]. Furthermore, the correct composition of leguminous trees at strategic locations can help promote BD, soil fertility, and the quality of water and soil. Integration of trees that can provide additional benefits such as fodder, nutrients, and biomass and may serve BD conservation.
Since AF reduces disturbance, synthetic chemical use, and mono cropping, strategical planning and implementation can help further improve BD. Fertilizers, pesticides, herbicides, and other chemicals exert a strong impact on BD [138]. The use of synthetic inorganic fertilizers reduces soil BD [119,138] while reduced or no-tillage favors greater diversity as tillage destroys fungal mycelial networks [139,140]. These impacts can cause shifts within soil and affect all functions. According to Thiele-Bruhn et al. [141] agricultural system with less input may promote higher BD. Fertilizer application and the use of other chemical can be minimized as AF naturally improves soil C, nutrients, and other physical properties. This approach can help especially, for rapidly developing tropics, as small and diversified AF is a viable land use strategy for biodiversity conservation, sustainable agriculture, and ecosystem services [11,142].
Agroforestry also serves as a corridor between the forest and the other lands and thus serves as a habitat outside of protected forests. In Nicaragua, coffee AF has served as a wildlife habitat corridor for mantled howling monkeys, Alouatta palliate [143]. In Khao Luang Mountains, Thailand, Round et al. [144] found 38–48% of bird species inhabiting neighboring forests in mixed fruit orchards. Establishment and maintenance of AF adjacent to forest can help conserve BD while improving the aesthetic values of the landscape.
Agroforestry causes spatially heterogeneous and/or concentrated communities due to spatial arrangement of trees, shrubs, and grasses [123,145]. For instance, the richness of bacteria was greater with white ash and poplar than Norway spruce in Canada [146]. Another study reported specific AMF communities in corn roots growing in Norway spruce AF compared to silver maple in Canada [145]. In Iberian Dehesas, high BD can be partly explained by the existence of scattered trees that create disproportionate differences in above- and below-ground in AF are the key features for niche densities and high BD [147]. Supporting the above observation, Fischer et al. [148], in an Australian livestock grazing landscape, found a strong relationship between number of trees, birds, and bats. In their study, bird richness doubled with one tree as compared to treeless landscapes. Furthermore, the study found three times more bats with 3–5 trees as compared to no tree and bat activity increased by a factor of 100. These findings may imply that proper selection of specific tree/crop combinations can be used for further enrich soil and nutrients and/or decontaminate soil and water for environmental benefits by AF induced BD.
Although diversity and abundance are beneficial within the AF, Barrios et al. [149] have cautioned about making generalizations. For instance, Sileshi and Mafongoya [150] reported an abundance of earthworms and beetles with legumes producing high quality biomass and increases of millipedes and beetles with legumes producing low quality biomass. Organisms obtain energy from carbon with a few exceptions and thus changes in plant communities, plant litter, litter composition and forms, as well as dead and live soil fauna can manipulate soil functions such as energy source and affecting their functions. Agroforestry’s diverse plant communities and structures can modify the important soil functions and with possible feedback to the above- and below-ground components of the AF itself and accompanying commodity crops.
Conversion of AF to monocultures reduce BD and conversion of monoculture to AF increases BD as AF harbors greater species richness and diversity [2,32,33,34,45]. However, due to intensive management, AF supports lower number of species as compared to adjacent undisturbed forests [27,30]. Despite numerous benefits of AF on BD, conservation there are some negative consequences of AF adoption. Agroforestry can promote undesirable species including invasive species if proper measures were not implemented. For example, a large number of biomass/fuel wood projects introduced in 1970s and 1980s in developing countries have impacted wildlife, water availability, and soil nutrient status [44]. In Kenya and India, ground water level has dropped in tea plantations with Eucalyptus trees and now both countries are trying to stop new eucalyptus plantings.
The role of AF has been controversial in regions with forest expansion for agriculture and settlements [151]. The conversion of forests for cocoa or palm oil plantations have reduced the forest cover and BD. Secondary, management practices such as shade-grown cocoa and palm plants with other trees have enhanced the BD as compared to the monocrop [66]. A comparison between the shade-grown crop and the monocrop implied BD enhancement with both cocoa and coffee. However, monocrop and the shade-grown systems had reduced diversity as compared to the original forest plantations.
Proper planning and strategic placement of soil-site-climate appropriate species combinations can help reduce negative impacts to the environment and landscapes while improving numerous ecosystem benefits including BD. For example, establishment of tall fescue (Festuca arundinacea) on AF buffers and other strategic locations within a watershed can help degradation of herbicides and antibiotics by soil communities and retention of these chemicals without further damaging water bodies [152,153]. In AF, strategic tree/shrub/plant selection and arrangements can enhance certain soil activities as some AMF preferentially select host plant [154]. It is also important to consider societal interest and needs when proposing new tree species and long-term impacts of those trees on the landscape. A good understanding of ecosystem services, society needs, and AF knowledge could also facilitate improvements in policies and support services.
5. Conclusions
Global scale agricultural intensification, population growth, dietary patterns, industrialization, and climate change are major causes of the degradation of BD and ecosystem functions at an exponential rate. Adoption of AF could help reverse both types of damages and improve BD and ecosystem services. Numerous studies have reported increased BD by AF. Agroforestry induced BD can be attributed to food, shelter, habitat, protection, refuge, favorable microclimate, improved soil-plant-water relationships, and other resources provided by multi-species vegetation of AF. The review also emphasizes the importance of multi-species integration for greater heterogeneity and for creation of spatially heterogeneous patched that could help enhance specific soil-plant communities. The selection of site-climate suitable combinations can be used to further enhance BD and services provided by the enhanced BD. However, proper planning must be conducted before the practices are adopted. These include selection of soil-site-climate suitable species and consideration of social and local needs.
Newer molecular techniques including DNA analysis, PLFA, metagenomics, and metabolomics can be used to improve our knowledge and benefits of AF induced BD. These may include understanding morphological and genetic diversity and their specific contributions for ecosystem services. Simulation of appropriate models may enhance our understanding and to predict changes for short- and long-term benefits of adopted practices. For example, Agricultural Production Systems sIMulator (APSIM) and HiSAFE could be used to understand and quantify various benefits and interactions. Long-term impacts of benefits should be evaluated for proposing new management strategies for enhancement of BD and ecosystem services including environmental, production, and economic benefits.
Author Contributions
Investigation, R.U., L.R.; Project administration, R.U. and S.J.; Supervision, R.U.; Visualization, L.R.; Writing—original draft, R.U.; Writing—review & editing, L.R. and S.J.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AF | agroforestry |
| AMF | arbuscular mycorrhizae fungi |
| BD | biodiversity |
| SOC | soil organic carbon |
References
- Pimm, S.L.; Russell, G.J.; Gittleman, J.L.; Brooks, T.M. The future of biodiversity. Science 1995, 269, 269–350. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for conserving and enhancing biodiversity. Agrofor. Syst. 2012, 85, 1–8. [Google Scholar] [CrossRef]
- Pimm, S.L. Biodiversity: Climate change or habitat loss-which will kill more species? Curr. Biol. 2008, 18, R117–R119. [Google Scholar] [CrossRef]
- Rands, M.R.; Adams, W.M.; Bennun, L.; Butchart, S.H.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Kessler, M.; Barkmann, J.; Bos, M.M.; Buchori, D.; Erasmi, S.; Faust, H.; Gerold, G.; Glenk, K.; Gradstein, S.R.; et al. Tradeoffs between income, biodiversity, and ecosystem functioning during tropical rainforest conversion and agroforestry intensification. Proc. Natl. Acad. Sci. USA 2007, 104, 4973–4978. [Google Scholar] [CrossRef]
- Culman, S.W.; Young-Mathews, A.; Hollander, A.D.; Ferris, H.; Sanchez-Moreno, S.; O’Green, A.T.; Jackson, L.E. Biodiversity is associated with indicators of soil ecosystem functions over a landscape gradient of agricultural intensification. Landsc. Ecol. 2010, 25, 1333–1348. [Google Scholar] [CrossRef]
- Vigouroux, Y.; Barnaud, A.; Scarcelli, N.; Thuillet, A.C. Biodiversity, evolution and adaptation of cultivated crops. C. R. Biol. 2011, 334, 450–457. [Google Scholar] [CrossRef]
- Thrupp, L.A. Linking agricultural biodiversity and food security: The valuable role of agrobiodiversity for sustainable agriculture. Int. Aff. 2000, 76, 283–297. [Google Scholar] [CrossRef]
- Sistla, S.A.; Roddy, A.B.; Williams, N.E.; Kramer, D.B.; Stevens, K.; Allison, S.D. Agroforestry practices promote biodiversity and natural resource diversity in Atlantic Nicaragua. PLoS ONE 2016, 11, e0162529. [Google Scholar] [CrossRef]
- Kleijn, D.; Sutherland, W.J. How effective are European agri-environment schemes in conserving and promoting biodiversity? J. Appl. Ecol. 2003, 40, 947–969. [Google Scholar] [CrossRef]
- Opermmann, R.; Beaufoy, G.; Jones, G. High Nature Value Farming in Europe: 35 European Countries—Experiences and Perspectives; Verlag Regionanlkultur: Ubstadt-Weiher, Germany, 2012. [Google Scholar]
- Kristensen, P. EEA Core Set of Indicators; Revised Version April 2003; European Environment Agency: Copenhagen, Denmark, 2003. [Google Scholar]
- McNeely, J.A.; Schroth, G. Agroforestry and biodiversity conservation—Traditional practices, present dynamics, and lessons for the future. Biodivers. Conserv. 2006, 15, 549–554. [Google Scholar] [CrossRef]
- Sanchez, P.A. Science in agroforestry. Agrofor. Syst. 1995, 30, 5–55. [Google Scholar] [CrossRef]
- Dobson, A.P.; Bradshaw, A.D.; Baker, A.J.M. Hopes for the future: Restoration ecology and conservation biology. Science 1997, 277, 515–522. [Google Scholar] [CrossRef]
- Huang, W.; Kanninen, M.; Xu, Q.; Huang, B. Agroforestry in China: Present state and future potential. Ambio 1997, 26, 393–397. [Google Scholar]
- Huang, W. Productive coexistence and gain in agroforestry systems. Acta For. Fenn. 1998, 260, 1–72. [Google Scholar] [CrossRef]
- Leakey, R.R.B. Agroforestry for biodiversity in farming systems. In Biodiversity in Agroecosystems; Collins, W.W., Qualset, C.O., Eds.; CRC Press: New York, NY, USA, 1999; pp. 127–145. [Google Scholar]
- Boffa, J.M. Agroforestry Parklands in Sub-Saharan Africa. In FAO Conservation Guide 34; Food and Agriculture Organization of the United Nations: Rome, Italy, 1999; Available online: http://www.fao.org/DOCREP/005/X3940E/X3940E00.htm (accessed on 7 May 2019).
- Huang, W.; Luukkanen, O.; Johanson, S.; Kaarakka, V.; Räisänen, S.; Vihemäki, H. Agroforestry for biodiversity conservation of nature reserves: Functional group identification and analysis. Agrofor. Syst. 2002, 55, 65–72. [Google Scholar] [CrossRef]
- Buck, L.E.; Gavin, T.A.; Lee, D.R.; Uphoff, N.T.; Behr, D.C.; Drinkwater, L.E.; Hively, W.D.; Werner, F.R. Ecoagriculture: A Review and Assessment of its Scientific Foundations; Cornell University: Ithaca, NY, USA, 2004. [Google Scholar]
- Méndez, V.E.; Lok, R.; Somarriba, E. Interdisciplinary analysis of home gardens in Nicaragua: Micro- zonation, plant use and socioeconomic importance. Agrofor. Syst. 2001, 51, 85–96. [Google Scholar] [CrossRef]
- Hemp, A. The banana forests of Kilimanjaro: Biodiversity and conservation of the Chagga home gardens. In Forest Diversity and Management. Topics in Biodiversity and Conservation; Hawksworth, D.L., Bull, A.T., Eds.; Springer: Amsterdam, The Netherlands, 2006; pp. 133–155. [Google Scholar]
- Borkhataria, R.R.; Collazo, J.A.; Groom, M.J. Species abundance and potential biological control services in shade vs. sun coffee in Puerto Rico. Agric. Ecosyst. Environ. 2012, 151, 1–5. [Google Scholar] [CrossRef]
- Noble, I.R.; Dirzo, R. Forests as human-dominated ecosystems. Science 1997, 277, 522–525. [Google Scholar] [CrossRef]
- Thevathasan, N.V.; Gordon, A.M. Ecology of tree intercropping system in the North temperate region: Experience from southern Ontario, Canada. Agrofor. Syst. 2004, 61, 257–268. [Google Scholar]
- Dollinger, J.; Jose, S. Agroforestry for soil health. Agrofor. Syst. 2018, 92, 213–219. [Google Scholar] [CrossRef]
- Bhagwat, S.A.; Willis, K.J.; Briks, H.J.B.; Whittaker, R.J. Agroforestry: A refuge for tropical biodiversity? Trends Ecol. Evol. 2008, 23, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Torralba, M.; Fagerholm, N.; Burgess, P.J.; Moreno, G.; Plieninger, T. Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agric. Ecosyst. Environ. 2016, 230, 150–161. [Google Scholar] [CrossRef]
- Perfecto, I.; Rice, R.A.; Greenberg, R.; Van der Voort, M.E. Shade coffee: A disappearing refuge for biodiversity. Bioscience 1996, 46, 598–608. [Google Scholar] [CrossRef]
- Lawton, J.H.; Bignell, D.E.; Bolton, B.; Bloemers, G.F.; Eggleton, P.; Hammond, P.M.; Hodda, M.; Holt, R.D.; Larsen, T.B.; Mawdsley, N.A.; et al. Biodiversity inventories, indicator taxa and effects of habitat modification in tropical forest. Nature 1998, 391, 72–76. [Google Scholar] [CrossRef]
- Schroth, G.; da Fonseca, G.A.B.; Harvey, C.A. Agroforestry and Biodiversity Conservation in Tropical Landscapes; Island Press: Washington, DC, USA, 2004; p. 523. [Google Scholar]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Alkemade, R.; Van Oorschot, M.; Miles, L.; Nellemann, C.; Bakkenes, M.; Brink, B. GLOBIO3: A framework to investigate options for reducing global terrestrial biodiversity loss. Ecosystems 2009, 12, 374–390. [Google Scholar] [CrossRef]
- Barton, J.; Pretty, J. What is the best dose of nature and green exercise for improving mental health? A multi-study analysis. Environ. Sci. Technol. 2010, 44, 947–3955. [Google Scholar] [CrossRef]
- Leakey, R.; Tchoundjeu, Z.; Schreckenberg, K.; Simons, T.; Shackleton, S.; Mander, M.; Wynberg, R.; Shackleton, C.; Sullivan, C. Trees and markets for agroforestry tree products: Targeting poverty reduction and enhanced livelihoods. In World Agroforestry into the Future; Garrity, D., Okono, A., Grayson, M., Parrott, S., Eds.; World Agroforestry Centre: Nairobi, Kenya, 2006; pp. 11–22. [Google Scholar]
- Jose, S.; Gillespie, A.R.; Pallardy, S.G. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 61, 237–255. [Google Scholar]
- Schulze, C.H.; Waltert, M.; Kessler, P.J.; Pitopang, R.; Veddeler, D.; Mühlenberg, M.; Gradstein, S.R.; Leuschner, C.; Steffan-Dewenter, I.; Tscharntke, T. Biodiversity indicator groups of tropical land-use systems: Comparing plants, birds, and insects. Ecol. Appl. 2004, 14, 1321–1333. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Garrett, H.E.; Kallenbach, R.L. Agroforestry buffers for nonpoint source pollution reductions from agricultural watersheds. J. Environ. Qual. 2011, 40, 800–806. [Google Scholar] [CrossRef]
- Benayas, J.M.R.; Bullock, J.M. Restoration of biodiversity and ecosystem services on agricultural land. Ecosystems 2012, 15, 883–889. [Google Scholar] [CrossRef]
- Swallow, B.; Boffa, J. The potential for agroforestry to contribute to the conservation and enhancement of landscape biodiversity. In World Agroforestry into the Future; Garrity, D., Okono, A., Grayson, M., Parrott, S., Eds.; World Agroforestry Centre: Nairobi, Kenya, 2006; pp. 95–101. [Google Scholar]
- Varah, A.; Jones, H.; Smith, J.; Potts, S.G. Enhanced biodiversity and pollination in UK agroforestry systems. J. Sci. Food Agric. 2013, 93, 2073–2075. [Google Scholar] [CrossRef]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends. Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef]
- Wolz, K.J.; DeLucia, E.H. Alley cropping: Global patterns of species composition and functions. Agric. Ecosyst. Environ. 2018, 252, 61–68. [Google Scholar] [CrossRef]
- Garrett, H.E.; McGraw, R.L.; Walter, W.D. Alley Cropping Practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 133–162. [Google Scholar]
- Chamberlain, J.L.; Mitchell, D.; Bringham, T.; Hobby, T.; Zabek, L.; Davis, J. Forest framing practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 219–256. [Google Scholar]
- Elliot, K.; Vose, J.M. Riparian area harvesting impacts on vegetation composition and diversity: Headwaters to Estuaries. In Advances in Watershed Science and Management; Younos, T., Parece, T.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; p. 104. [Google Scholar]
- Naiman, R.J.; Décamps, H.; McClain, M.E. Structutal pattern. In Riparia; Naiman, R.J., Décamps, H., McClain, M.E., Eds.; Elsevier Academic Press: New York, NY, USA, 2005; pp. 79–123. [Google Scholar]
- Sharrow, S.H.; Brauer, D.; Clason, T.R. Silvopastu practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 105–131. [Google Scholar]
- Orefice, J.; Carroll, J.; Conroy, D.; Ketner, L. Silvopasture practices and perspectives in the Northeasten United States. Agrofor. Syst. 2017, 91, 149–160. [Google Scholar] [CrossRef]
- Kort, J.; Turnock, R. Carbon Reservoir and Biomass in Canadian Prairie Shelterbelts. Agrofor. Syst. 1999, 44, 175–186. [Google Scholar] [CrossRef]
- Schultz, R.C.; Isenhart, T.M.; Colletti, J.P.; Simpkins, W.W.; Udawatta, R.P.; Schultz, P.L. Riparian and upland buffer practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy Inc.: Madison, WI, USA, 2009; pp. 163–218. [Google Scholar]
- Afta.org. Available online: https://www.aftaweb.org/about/what-is-agroforestry/silvopasture.html (accessed on 7 May 2019).
- Bucheli, V.J.P.; Bokelmann, W. Agroforestry systems for biodiversity and ecosystem services: The case of the Sibundoy Valley in the Colombian province of Putumayo. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2017, 13, 380–397. [Google Scholar]
- Kumar, B.M.; Nair, P.K.R. Introduction, Tropical Home Gardens: A Time-Tested Example of Sustainable Agroforestry; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–12. [Google Scholar]
- Kabir, M.E.; Webb, E.L. Can home gardens conserve biodiversity in Bangladesh? Biotropica 2008, 40, 95–103. [Google Scholar]
- Kumari, M.A.S.; Kansuntisukmongkol, K.; Brockelman, W.Y. Plant diversity in home gardens and its contribution to household economy in suburban areas in Sri Lanka. Environ. Nat. Resour. J. 2009, 7, 12–30. [Google Scholar]
- Bardhan, S.; Jose, S.; Biswas, S.; Kabir, K.; Rogers, W. Home garden agroforestry systems: An intermediary for biodiversity conservation in Bangladesh. Agrofor. Syst. 2012, 85, 29–34. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Van Noordwijk, M.; Wasrin, U.M.; Tomich, T.P.; Gillison, A.N. Environmental benefits and sustainable land-use options in the Jambi transect, Sumatra, Indonesia. J. Veg. Sci. 2002, 13, 429–438. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Ewel, J.J. Natural systems as models for the design of sustainable systems of land use. Agrofor. Syst. 1999, 45, 1–21. [Google Scholar] [CrossRef]
- Kindt, R. Methodology for Tree Species Diversification Planning for African Agroecosystems. Ph.D. Thesis, University of Ghent, Ghent, Belgium, 2002. [Google Scholar]
- Donald, P.F. Biodiversity impacts of some agricultural commodity production systems. Conserv. Biol. 2004, 18, 17–37. [Google Scholar] [CrossRef]
- Gillison, A.N.; Liswanti, N.; Budidarsono, S.; Noordwijk, M.V.; Tomich, T.P. Impact of cropping methods on biodiversity in coffee agroecosystems in Sumatra, Indonesia. Ecol. Soc. 2004, 9, 7. [Google Scholar] [CrossRef]
- Khan, M.L.; Arunachalam, A. Traditional agroforestry as a viable choice to conserve agro-biodiversity in the northeast India. In Agroforestry: Potentials and Opportunities; Pathak, P.S., Newaj, R., Eds.; Agrobios (India) and Indian Society of Agroforestry: Jodhpur, India, 2003; pp. 95–105. [Google Scholar]
- Michon, G.; De Foresta, H. The Indonesian agro-forest model. Forest resource management and biodiversity conservation. In Conserving Biodiversity Outside Protected Areas. The Role of Traditional Agro-Ecosystems; Halladay, P., Gilmour, D.A., Eds.; World Conservation Union, Forest Conservation Programme: Andalucía, Spain, 1995; pp. 90–106. [Google Scholar]
- Fanaye, A.O.; Adeyemi, E.A.; Olaiya, A.O. Spacing experiments in cocoa/kola/citrus intercrop. In Proceedings of the 14th International Cocoa Research Conference, Accra, Ghana, 13–18 October 2003. [Google Scholar]
- Asare, R. Cocoa Agroforests in West Africa: A Look at Activities on Preferred Trees in the Farming Systems; Forest & Landscape Working Papers No. 6-2005; Forest & Landscape Denmark (FLD): Hørsholm, Denmark, 2005. [Google Scholar]
- Osei-Bonsu, K.; Ameyaw-Oduro, C.; Tetteh, J.P. Traditional cocoa agroforestry: 1. Species encountered in the cocoa ecosystem of a typical cocoa-growing district in Ghana. In Proceedings of the 14th International Cocoa Research Conference, Accra, Ghana, 13–18 October 2003; pp. 13–18. [Google Scholar]
- Tomich, T.P.; Van Noordwijk, M.; Vosti, S.A.; Witcover, J. Agricultural development with rainforest conservation: Methods for seeking best bet alternatives to slash-and-burn, with applications to Brazil and Indonesia. Agric. Econ. 1998, 19, 159–174. [Google Scholar] [CrossRef]
- Tomich, T.P.; Van Noordwijk, M.; Budidarsono, S.; Gillison, A.; Kusumanto, T.; Murdiyarso, M.; Stolle, F.; Fagi, A.M. Agricultural intensification, deforestation, and the environment: Assessing tradeoffs in Sumatra, Indonesia. In Tradeoffs or Synergies? Agricultural Intensification, Economic Development and the Environment; Lee, D.R., Barrett, C.B., Eds.; CAB International: Wallingford, UK, 2001. [Google Scholar]
- Kearns, C.A.; Inouye, D.W. Pollinators, flowering plants, and conservation biology. Bioscience 1997, 47, 297–307. [Google Scholar] [CrossRef]
- Klein, A.M.; Steffan–Dewenter, I.; Tscharntke, T. Fruit set of highland coffee increases with the diversity of pollinating bees. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 955–961. [Google Scholar] [CrossRef]
- Reitsma, R.; Parrish, J.D.; McLarney, W. The role of cacao plantations in maintaining forest avian diversity in southeastern Costa Rica. Agrofor. Syst. 2001, 53, 185–193. [Google Scholar] [CrossRef]
- Gibbs, S.; Koblents, H.; Coleman, B.; Gordon, A.; Thevathasan, N.; Williams, P. Avian diversity in a temperate tree-based intercropping system from inception to now. Agrofor. Syst. 2016, 90, 905–916. [Google Scholar] [CrossRef]
- Williams, P.; Koblents, H.; Gordon, A. Bird use of an intercropped corn field and old field in southern Ontario, Canada. In Proceedings of the 4th North American Agroforestry Conference, Boise, ID, USA, 23–28 July 1995; Ehrenreich, J.H., Ehrenreich, D.L., Lee, H.W., Eds.; University of Idaho: Moscow, ID, USA, 1995; pp. 158–162. [Google Scholar]
- Harvey, C.A.; Gonzalez-Villalobos, J.A. Agroforestry systems conserve species-rich but modified assemblages of tropical birds and bats. Biodivers. Conserv. 2007, 16, 2257–2292. [Google Scholar] [CrossRef]
- Naidoo, R. Species richness and community composition of songbirds in a tropical forest-agriculture landscape. Anim. Conserv. 2004, 7, 93–105. [Google Scholar] [CrossRef]
- Soini, E. Dynamics of livelihood strategies in the changing landscapes of the southern slopes of Mt. Kilimanjaro, Tanzania. Agric. Syst. 2005, 85, 306–323. [Google Scholar] [CrossRef]
- Griffith, D.M. Agroforestry: A refuge for tropical biodiversity after fire. Conserv. Biol. 2000, 14, 325–326. [Google Scholar] [CrossRef]
- Zomer, R.J.; Neufeldt, H.; Xu, J.; Ahrends, A.; Bossio, D.; Trabucco, A.; Van Noordwijk, M.; Wang, M. Global Tree Cover and Biomass Carbon on Agricultural Land: The contribution of agroforestry to global and national carbon budgets. Sci. Rep. 2016, 6, 29987. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Beck, M.M. Influences of shelterbelts on wildlife management and biology. Agric. Ecosyst. Environ. 1998, 22, 301–335. [Google Scholar]
- Söderström, B.O.; Pärt, T. Influence of landscape scale on farmland birds breeding in semi-natural pastures. Conserv. Biol. 2000, 14, 522–533. [Google Scholar]
- Barrios, E.; Valencia, V.; Jonsson, M.; Brauman, A.; Hairiah, K.; Mortimer, P.E.; Okubo, S. Contribution of trees to the conservation of biodiversity and ecosystem services in agricultural landscapes. Int. J. Biodiv. Sci. Ecosyst. Serv. Manag. 2017, 14, 1–16. [Google Scholar] [CrossRef]
- Donaldson, J.; Nänni, I.; Zachariades, C.; Kemper, J. Effects of habitat fragmentation on pollinator diversity and plant reproductive success in Renosterveld shrub-lands of South Africa. Conserv. Biol. 2002, 5, 1267–1276. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.M.; Steffan-Dewenter, I.; Tscharntke, T. Rain forest promotes trophic interactions and diversity of trap-nesting Hymenoptera in adjacent agroforestry. J. Anim. Ecol. 2006, 75, 315–323. [Google Scholar] [CrossRef]
- Karp, D.S.; Mendenhall, C.D.; Sandi, R.F.; Chaumont, N.; Ehrlich, P.R.; Hadly, E.A.; Daily, G.C. Forest bolsters bird abundance, pest control and coffee yield. Ecol. Lett. 2013, 16, 1339–1347. [Google Scholar] [CrossRef]
- Brandle, J.R.; Hodges, L.; Zhou, X. Windbreaks in sustainable agriculture. Agrofor. Syst. 2004, 61, 65–78. [Google Scholar]
- Moreno, G.; Gonzalez-Bornay, G.; Pulido, F.; Lopez-Diaz, M.L.; Bertomeu, M.; Jua’rez, E.; Diaz, M. Exploring the causes of high biodiversity of Iberian dehesas: The importance of wood pastures and marginal habitats. Agrofor. Syst. 2016, 90, 87–105. [Google Scholar] [CrossRef]
- Price, G.W.; Gordon, A.M. Spatial and temporal distribution of earthworms in a temperate intercropping system in southern Ontario, Canada. Agrofor. Syst. 1999, 44, 141–149. [Google Scholar] [CrossRef]
- Pauli, N.; Oberthur, T.; Barrios, E.; Conacher, A.J. Fine-scale spatial and temporal variation in earthworm surface casting activity in agroforestry fields, western Honduras. Pedobiologia 2010, 53, 127–139. [Google Scholar] [CrossRef]
- Cardinael, R.; Guenet, B.; Chevallier, T.; Dupraz, C.; Cozzi, T.; Chenu, C. High organic inputs explain shallow and deep SOC storage in a long-term agroforestry system–combining experimental and modeling approaches. Biogeosciences 2018, 15, 297–317. [Google Scholar] [CrossRef]
- Spurgeon, D.J.; Keith, A.M.; Schmidt, O.; Lammertsma, D.R.; Faber, J.H. Land-use and land-management change: Relationships with earthworm and fungi communities and soil structural properties. BMC Ecol. 2013, 13, 46. [Google Scholar] [CrossRef] [PubMed]
- Keesstra, S.D.; Bouma, J.; Wallinga, J.; Tittonell, P.; Smith, P.; Cerdà, A.; Montanarella, L.; Quinton, J.N.; Pachepsky, Y.; Van Der Putten, W.H.; et al. The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. Soil 2016, 2, 111–128. [Google Scholar] [CrossRef]
- Bainard, L.D.; Klironomos, J.N.; Gordon, A.M. Arbuscular mycorrhizal fungi in tree-based intercropping systems: A review of their abundance and diversity. Pedobiologia 2011, 54, 57–61. [Google Scholar] [CrossRef]
- Beuschel, R.; Piepho, H.P.; Joergensen, R.G.; Wachendorf, C. Similar spatial patterns of soil quality indicators in three poplar-based silvo-arable alley cropping systems in Germany. Biol. Fertil. Soils 2019, 55, 1–14. [Google Scholar] [CrossRef]
- Lacombe, S.; Bradley, R.L.; Hamel, C.; Beaulieu, C. Do tree-based intercropping systems increase the diversity and stability of soil microbial communities? Agric. Ecosyst. Environ. 2009, 131, 25–31. [Google Scholar] [CrossRef]
- Chifflot, V.; Rivest, D.; Olivier, A.; Cogliastro, A.; Khasa, D. Molecular analysis of arbuscular mycorrhizal community structure and spores distribution in tree-based intercropping and forest systems. Agric. Ecosyst. Environ. 2009, 131, 32–39. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, M.; Zhou, P.; Fang, Y.; Ji, Y. Impact of tree species on barley rhizosphere-associated fungi in an agroforestry ecosystem as revealed by 18S rDNA PCRDGGE. Agrofor. Syst. 2018, 92, 541–554. [Google Scholar]
- Kremer, R.J.; Kussman, R.D. Soil quality in a pecan–kura clover alley cropping system in the Midwestern USA. Agrofor. Syst. 2011, 83, 213–223. [Google Scholar] [CrossRef]
- Mutabaruka, R.; Mutabaruka, C.; Fernandez, I. Diversity of arbuscular mycorrhizal fungi associated to tree species in semiarid areas of Machakos, Kenya. Arid Land Res. Manag. 2002, 16, 385–390. [Google Scholar] [CrossRef]
- Pande, M.; Tarafdar, J.C. Arbuscular mycorrhizal fungal diversity in neem-based agroforestry systems in Rajasthan. Appl. Soil Ecol. 2004, 26, 233–241. [Google Scholar] [CrossRef]
- Banarjee, S.; Baah-Acheamfour, M.; Carlyle, C.M.; Bissett, A.; Richardon, A.E.; Siddique, T.; Bork, E.W.; Chang, S.X. Determinants of bacterial communities in Canadian agroforestry systems. Environ. Microbiol. 2015, 18, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Kremer, R.J.; Hezel, L.F. Soil quality improvement under an ecologically based farming system in northwest Missouri. Renew. Agric. Food Syst. 2013, 28, 245–254. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef]
- Keith, A.M.; Van der Wal, R.; Brooker, R.W.; Osler, G.H.R.; Chapman, S.J.; Burslem, D.F.R.P.; Elson, D.A. Increasing litter species richness reduces variability in a terrestrial decomposer system. Ecology 2008, 89, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Rivest, D.; Lorente, M.; Olivier, A.; Messier, C. Soil biochemical properties and microbial resilience in agroforestry systems: Effects on wheat growth under controlled drought and flooding conditions. Sci. Total Environ. 2013, 463, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Gupta, S.R.; Singh, G. Soil carbon, microbial activity, and nitrogen availability in a agroforestry systems on moderately alkaline soils in northern India. Appl. Soil Ecol. 2000, 15, 283–294. [Google Scholar] [CrossRef]
- Wojewoda, D.; Russel, S. The impact of a shelterbelt on soil properties an microbial activity in an adjacent corp field. Pol. J. Ecol. 2003, 51, 291–307. [Google Scholar]
- Amador, J.A.; Glucksman, A.M.; Lyons, J.B.; Gorres, J.H. Spatial distribution of soil phosphatase activity within a riparian forest. Soil Sci. 1997, 162, 808–825. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Decker, K.L.M.; Sutherland, E.K. Prescribed burning effects on soil enzyme activity in a southern Ohio hardwood forest: A landscape-scale analysis. Soil Biol. Biochem. 2000, 32, 899–908. [Google Scholar] [CrossRef]
- Mungai, N.W.; Motavalli, P.P.; Kremer, R.J.; Nelson, K.A. Spatial variation of soil enzyme activities and microbial functional diversity in temperate alley cropping practices. Biol. Fertil. Soils 2006, 42, 129–136. [Google Scholar] [CrossRef]
- Dornbush, M.E. Grasses, litter and their interaction affects microbial biomass and soil enzyme activity. Soil Biol. Biochem. 2007, 39, 2241–2249. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Kremer, R.J.; Adamson, B.W.; Anderson, S.H. Variations in soil aggregate stability and enzyme activities in a temperate agroforestry practice. Appl. Soil Ecol. 2008, 39, 153–160. [Google Scholar] [CrossRef]
- Helgason, B.L.; Walley, F.L.; Germida, J.J. No-till and soil management increases microbial biomass and alters community profiles in soil aggregates. App. Soil Ecol. 2010, 46, 390–397. [Google Scholar] [CrossRef]
- Adhikari, K.; Hartemink, A.E.; Minasny, B.; BouKheir, R.; Greve, M.B.; Greve, M.H. Digital mapping of soil organic carbon contents and stocks in Denmark. PLoS ONE 2014, 9, e105519. [Google Scholar] [CrossRef] [PubMed]
- Udawatta, R.P.; Anderson, S.H. CT-measured pore characteristics of surface and subsurface soils as influenced by agroforestry and grass buffers. Geoderma 2008, 145, 381–389. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Kremer, R.J.; Garrett, H.E.; Anderson, S.H. Soil enzyme activities and physical properties in a watershed managed under agroforestry and row-crop system. Agric. Ecosyst. Environ. 2009, 131, 98–104. [Google Scholar] [CrossRef]
- Paudel, B.; Udawatta, R.P.; Kremer, R.J.; Anderson, S.H. Agroforestry and grass buffer effects on soil quality parameters for grazed pasture and row-crop systems. Appl. Soil Ecol. 2011, 48, 125–132. [Google Scholar] [CrossRef]
- Paudel, B.; Udawatta, R.P.; Kremer, R.J.; Anderson, S.H. Soil quality indicator responses to row crop, grazed pasture, and agroforestry buffer management. Agrofor. Syst. 2012, 84, 311–323. [Google Scholar] [CrossRef]
- Meyers, R.T.; Zak, D.R.; White, D.C.; Peacock, A. Landscape-level patterns of microbial community composition and substrate use in upland forest ecosystems. Soil Sci. Soc. Am. J. 2001, 65, 359–367. [Google Scholar] [CrossRef]
- Weerasekara, C.; Udawatta, R.P.; Jose, S.; Kremer, R.J.; Weerasekara, C. Soil quality differences in a row-crop watershed with agroforestry and grass buffers. Agrofor. Syst. 2016, 90, 829–838. [Google Scholar] [CrossRef]
- Bambrick, A.D.; Whallen, J.K.; Bradley, R.L.; Cogliastro, A.; Gordon, A.M.; Olivier, A.; Thevathasan, N.V. Spatial heterogeneity of organic carbon in tree-based inter cropping systems in Quebec and Ontario, Canada. Agrofor. Syst. 2010, 79, 343–353. [Google Scholar] [CrossRef]
- Bardhan, S.; Jose, S.; Udawatta, R.P.; Fritschi, F. Microbial community diversity in a 21-year old temperate alley cropping system. Agrofor. Syst. 2013, 87, 1031–1041. [Google Scholar] [CrossRef]
- Nii-Annang, S.; Grünewald, H.; Freese, D.; Hüttl, R.F.; Dilly, O. Microbial activity, organic C accumulation and 13C abundance in soils under alley cropping systems after 9 years of recultivation of quaternary deposits. Biol. Fertil. Soils 2009, 45, 531–538. [Google Scholar] [CrossRef]
- Foley, J.A.; Defries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Burgess, P.J. Effects of agroforestry on farm biodiversity in the UK. Scott. For. 1999, 53, 24–27. [Google Scholar]
- Stamps, W.T.; Woods, T.W.; Linit, M.J.; Garrett, H.E. Arthropod diversity in alley cropped black walnut (Juglans nigra L.) stands in eastern Missouri, USA. Agrofor. Syst. 2002, 56, 167–175. [Google Scholar] [CrossRef]
- McNeely, J.A. Nature vs. nurture: Managing relationships between forests, agroforestry and wild biodiversity. Agrofor. Syst 2004, 61, 155–165. [Google Scholar]
- Seobi, T.; Anderson, S.H.; Udawatta, R.P.; Gantzer, C.J. Influence of grass and agroforestry buffer strips on soil hydraulic properties for an Albaqualf. Soil Sci. Soc. Am. J. 2005, 69, 893–901. [Google Scholar] [CrossRef]
- Kumar, S.; Anderson, S.H.; Bricknell, L.G.; Udawatta, R.P.; Gantzer, C.J. Soil hydraulic properties influenced by agroforestry and grass buffers for grazed pasture systems. J. Soil Water Conserv. 2008, 63, 224–232. [Google Scholar] [CrossRef]
- Svoma, B.M.; Fox, N.I.; Pallardy, Q.; Udawatta, R.P. Evapotranspiration differences between agroforestry and grass buffer systems. Agric. Water Manag. 2016, 176, 214–221. [Google Scholar] [CrossRef]
- Prober, S.M.; Left, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D.; et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Kleijn, D.; Kohler, F.; Báldi, A.; Batáry, P.; Concepción, E.D.; Clough, Y.; Díaz, M.; Gabriel, D.; Holzschuh, A.; Knop, E.; Kovács, A. On the relationship between farmland biodiversity and land-use intensity in Europe. Proc. R. Soc. Lond. Biol. Sci. 2008, 276, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Simmons, B.L.; Coleman, D.C. Microbial community response to transition from conventional to conservation tillage in cotton fields. App. Soil Ecol. 2008, 40, 518–528. [Google Scholar] [CrossRef]
- Mathimaran, N.; Ruh, R.; Jama, B.; Verchot, L.; Frossard, E.; Jansa, J. Impact of agricultural management on arbuscular mycorrhizal fungal communities in Ken-yan ferra sol. Agric. Ecosyst. Environ. 2007, 119, 22–32. [Google Scholar] [CrossRef]
- Thiele-Bruhn, S.; Bloem, J.; De Vries, F.T.; Kalbitz, K.; Wagg, C. Linking soil biodiversity and agricultural soil management. Curr. Opin. Environ. Sustain. 2012, 4, 523–528. [Google Scholar] [CrossRef]
- Swallow, B.; Russell, D.; Fay, C. Agroforestry and environmental governance. In World Agroforestry into the Future; Garrity, D., Okono, A., Grayson, M., Parrott, S., Eds.; World Agroforestry Centre: Nairobi, Kenya, 2006; pp. 85–94. [Google Scholar]
- Williams-Guillén, K.; McCann, C.; Sanchez, J.M.; Koontz, F. Resource availability and habitat use by mantled howling monkeys in a Nicaraguan coffee plantation: Can agroforests serve as core habitat for a forest mammal? Anim. Conserv. 2006, 9, 331–338. [Google Scholar] [CrossRef]
- Round, P.D.; Gale, G.A.; Brockelman, W.Y. A comparison of bird communities in mixed fruit orchards and natural forest at Khao Luang, southern Thailand. Biodivers. Conserv. 2006, 15, 2873–2891. [Google Scholar] [CrossRef]
- Bainard, L.D.; Koch, A.M.; Gordon, A.M.; Klironomos, J.N. Temporal and compositional differences of arbuscular mycorrhizal fungal communities in conventional monocropping and tree-based intercropping systems. Soil Biol. Biochem. 2012, 45, 172–180. [Google Scholar] [CrossRef]
- Bainard, L.D.; Kochb, A.M.; Gordon, A.M.; Newmaster, S.G.; Thevathasan, N.V.; Klironomosb, J.N. Influence of trees on the spatial structure of arbuscular mycorrhizal communities in a temperate tree-based intercropping system. Agric. Ecosyst. Environ. 2011, 144, 13–20. [Google Scholar] [CrossRef]
- Bergmeier, E.; Petermann, J.; Schröder, E. Geobotanical survey of wood-pasture habitats in Europe: Diversity, threats and conservation. Biodivers. Conserv. 2010, 19, 2995–3014. [Google Scholar] [CrossRef]
- Fischer, J.; Stott, J.; Law, B.S. The disproportionate value of paddock trees. Biol. Conserv. 2010, 143, 1564–1567. [Google Scholar] [CrossRef]
- Barrios, E.; Sileshi, G.W.; Shepherd, K.; Ferguson, S. Agroforestry and Soil Health: Linking Trees, Soil Biota, and Ecosystem Services. In Soil Ecology and Ecosystem Services, 1st ed.; Wall, D.H., Bardgett, R.D., Behan-Pelletier, V., Herrick, J.E., Jones, H., Ritz, K., Six, J., Strong, D.R., van der Putten, W.H., Eds.; Oxford University Press: Oxford, England, 2012; pp. 315–330. [Google Scholar]
- Sileshi, G.; Mafongoya, P.L. Quantity and quality of organic inputs from coppicing leguminous trees influence abundance of soil macrofauna in maize crops in eastern Zambia. Biol. Fertil. Soils 2007, 43, 333–340. [Google Scholar] [CrossRef]
- Angelsen, A.; Kaimowitz, D. Is agroforestry likely to reduce deforestation? In Agroforestry and Biodiversity Conservation in Tropical Landscapes; Schroth, G., da Fonseca, G.A.B., Harvey, C.A., Gascon, C., Vasconcelos, H.L., Izac, A.M.N., Eds.; Island Press: Washington, DC, USA, 2004; Chapter 5; pp. 87–106. [Google Scholar]
- Lin, C.H.; Lerch, R.N.; Goyne, K.W.; Garrett, H.E. Reducing herbicides and veterinary antibiotic losses from agroecosystems using vegetative buffers. J. Environ. Qual. 2011, 40, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.; Goyne, K.W.; Anderson, S.H.; Lin, C.H.; Udawatta, R.P. Veterinary antibiotic sorption to agroforestry buffer, grass buffer, and cropland soils. Agrofor. Syst. 2010, 79, 67–80. [Google Scholar] [CrossRef]
- Lerat, S.; Lapointe, L.; Gutjahr, S.; Piche, Y.; Vierheilig, H. Carbon partitioning in a split-root system of arbuscular mycorrhizal plants is fungal and plant species dependent. New Phytol. 2003, 157, 589–595. [Google Scholar] [CrossRef]
Figure 1.
Mean species abundance (MSA) for forests, pastures, agroforestry, and agricultural land management practices Source: [37].
Figure 1.
Mean species abundance (MSA) for forests, pastures, agroforestry, and agricultural land management practices Source: [37].

Figure 2.
Development of scientific data base during the last 30 years on relationships between agroforestry and biodiversity.
Figure 2.
Development of scientific data base during the last 30 years on relationships between agroforestry and biodiversity.

Figure 3.
Floral diversity of different agroforestry practices, Iberian Dehesa system in Extremadura, Spain (A), Silvopsature system in southeastern US (B), Alley cropping in UK (C), Forest farming with medicinal herbs (D), Silvoarable agroforestry system in UK (E), Alley cropping in Kenya (F), Cocoa based agroforest in Talamanca, Costa Rica (G) and Kandian Home Gardens in Sri Lanka (H).
Figure 3.
Floral diversity of different agroforestry practices, Iberian Dehesa system in Extremadura, Spain (A), Silvopsature system in southeastern US (B), Alley cropping in UK (C), Forest farming with medicinal herbs (D), Silvoarable agroforestry system in UK (E), Alley cropping in Kenya (F), Cocoa based agroforest in Talamanca, Costa Rica (G) and Kandian Home Gardens in Sri Lanka (H).

Figure 4.
Occurrence of genus in alley cropping Agroforestry (AF) of temperate, subtropical, and tropical zones. Source: [47].
Figure 4.
Occurrence of genus in alley cropping Agroforestry (AF) of temperate, subtropical, and tropical zones. Source: [47].

Figure 5.
Measured species richness (95% CI) of bee species (A), plant species (B), spider species (C), and earth worm species (D) in Iberian Dehesas, central western Spain [93].
Figure 5.
Measured species richness (95% CI) of bee species (A), plant species (B), spider species (C), and earth worm species (D) in Iberian Dehesas, central western Spain [93].

Figure 6.
Relationships between weight of earthworm casts (black filled circles, g), number of pruned trees (light gray), and canopy size (open circles) of a tree-based agroforesty practice in Quesungual Slash-and-Mulch, western Honduras [95].
Figure 6.
Relationships between weight of earthworm casts (black filled circles, g), number of pruned trees (light gray), and canopy size (open circles) of a tree-based agroforesty practice in Quesungual Slash-and-Mulch, western Honduras [95].

Figure 7.
Relationship between earthworm density and saturated hydraulic conductivity of soil confidence limits [97].
Figure 7.
Relationship between earthworm density and saturated hydraulic conductivity of soil confidence limits [97].

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
P. Udawatta, R.; Rankoth, L.; Jose, S. Agroforestry and Biodiversity. Sustainability 2019, 11, 2879. https://0-doi-org.brum.beds.ac.uk/10.3390/su11102879
AMA Style
P. Udawatta R, Rankoth L, Jose S. Agroforestry and Biodiversity. Sustainability. 2019; 11(10):2879. https://0-doi-org.brum.beds.ac.uk/10.3390/su11102879
Chicago/Turabian StyleP. Udawatta, Ranjith, Lalith Rankoth, and Shibu Jose. 2019. "Agroforestry and Biodiversity" Sustainability 11, no. 10: 2879. https://0-doi-org.brum.beds.ac.uk/10.3390/su11102879
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.