1. Introduction
Lead (Pb) is an abundant and pernicious environmental contaminant posing significant human health risks; the maximum safe concentration of Pb in drinking water is estimated at 0.01 ppm [
1]. Serious human health risks include immunological, neurological, cardiovascular, reproductive, as well as developmental impacts [
2]. The recovery of Pb, as opposed to the mere removal of Pb from the environment, is of prime interest as the United States Geological Survey estimates that only 17 years’ supply of global raw workable Pb reserves is available [
3]. Pb has an annual consumption rate of about 5 million t/a [
4] with a total of 83.3 × 10
6 t reserve remaining worldwide [
3].
Various chemical and physical methods have been implemented historically to remove Pb from polluted wastewaters, but few are financially and environmentally sustainable. Bioremediation is a cost-effective solution for the removal and recovery of heavy metals from solution. Various microorganisms have exhibited the capacity to remediate Pb(II) from polluted environments with the aid of various methods by reducing the mobility and bioavailability of Pb(II), including bioaccumulation, surface biosorption, extracellular sequestration, and bioprecipitation [
5]. Studies investigating biological Pb(II) removal are summarized in
Table 1.
The current research team has shown in previous studies that effective removal and recovery of Pb is possible using an industrially obtained microbial consortium, with the formation of a dark grey precipitate when using Luria Bertani broth (LB) as growth substrate [
10]. The precipitate in question was identified as predominantly PbS and a fractional amount of elemental Pb under anaerobic conditions [
11].
This study aimed to determine the effect of Pb(II) concentration and availability of nutrients on Pb(II) removal, growth, active Pb(II) precipitating microbes, and population distribution of the industrially obtained Pb(II)-resistant consortium. Experiments were conducted under anaerobic batch conditions at 35 °C. The study is applicable as a sustainable method is yet to be developed to not only remove but also recover Pb(II) from polluted wastewater. This method of bio-removal could provide an ideal removal method for large-scale implementation in various industries as a simple cost-effective method to remediate and rehabilitate Pb-containing effluents or environmental Pb contamination.
2. Materials and Methods
2.1. Project Overview
The experimental procedures implemented in this study are summarized in
Table 2 below. For comparison, the Pb(II) and substrate concentrations used were adopted from previous studies conducted by this team [
9,
12].
2.2. Materials
All batch reactor experiments were set up anaerobically in 100 mL serum bottles. A Pb stock solution was prepared with Pb(NO3)2 (Merck, Kennelworth, NJ, USA). Standard Miller Luria Bertani (LB) Broth (Sigma Aldrich, St Louis, MO, USA) was used as rich growth media made to a final concentration of 25 mg/L or simulated LB broth, which consists of double the amount of nutrients and a decreased concentration of NaCl. Metabolic activity measurements were conducted with the aid of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) and the organic solvent dimethyl sulfoxide (DMSO) at a wavelength of 550 nm (Sigma Aldrich, St. Louis, MO, USA). Nitrate levels were tested using a nitrate testing kit (Merck, Darmstadt, Germany) and measured photometrically using a Spectroquant Nova 600 (Merck, Darmstadt, Germany).
2.3. Microbial Culture
The specific Pb(II)-resistant microbial consortium was obtained from a borehole at an automotive battery recycling plant in Gauteng, South Africa. The inoculum was prepared by adding 1 g of Pb(II)-contaminated soil to a mixture of LB broth and 80 ppm Pb(II) in an anaerobic 100 mL serum bottle, and incubated for 24 h at 32 °C at 120 rpm. Glycerol was added to the matured inoculum to a final ratio of 20% v/v and stored cryogenically at −77 °C. The precultures were thereafter prepared from the cryogenically stored inoculum by adding one loop of inoculum to a 100 mL anaerobic serum bottle containing a mixture of LB broth and either 80 ppm Pb(II) or 500 ppm Pb(II). The reactors were purged with nitrogen gas for 3 min and sealed with a rubber stopper and metal clamp to enable an anaerobic environment, and they were incubated at 30 °C and 120 rpm for 3 d to test for contamination before inoculation of the experiments.
2.4. Experimental
The Pb(II) stock solution and growth medium (standard or simulated LB) were prepared and autoclaved separately, after which they were cooled to room temperature. The Pb(II) stock solution was added to the growth medium in a biological safety cabinet under sterile conditions. A 0.2 mL sample of inoculum (prepared pre-culture) was added to the serum bottles. The serum bottles were each purged with nitrogen gas for 3 min and sealed to ensure anaerobic conditions. The batch reactors were placed in a shaker incubator at 120 rpm and 35 °C for the duration of the experiment. All datasets were conducted in triplicate to ensure repeatability. Controlled sampling and analytical conditions were maintained to minimize experimental error. Abiotic controls were conducted to confirm that any changes that occur in the reactors are of biological and not chemical origin. The abiotic controls were run for a period of 72 h with no added inoculant to the reactor and incubating with the batch reactors to ensure equivalent conditions
2.5. Sampling
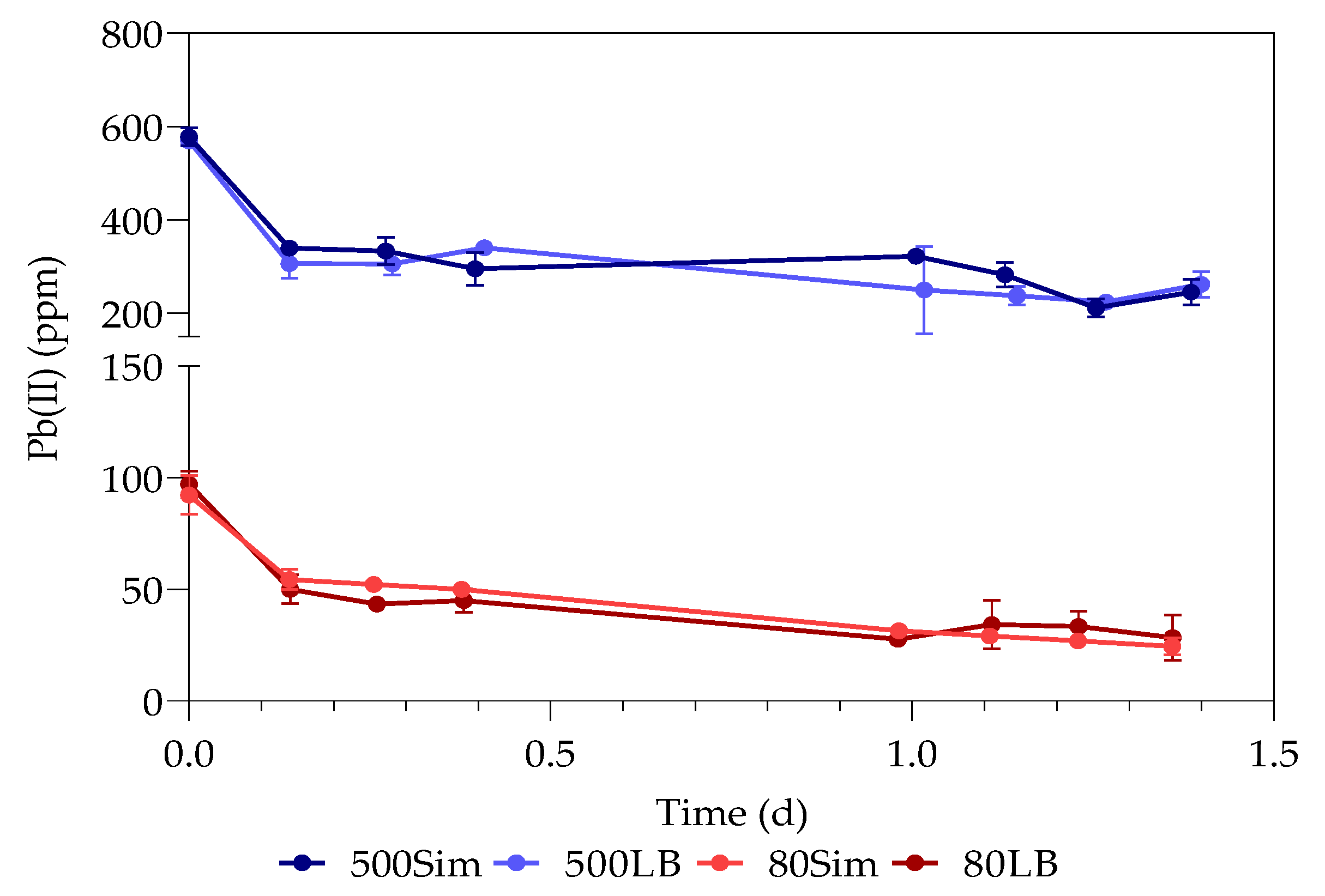
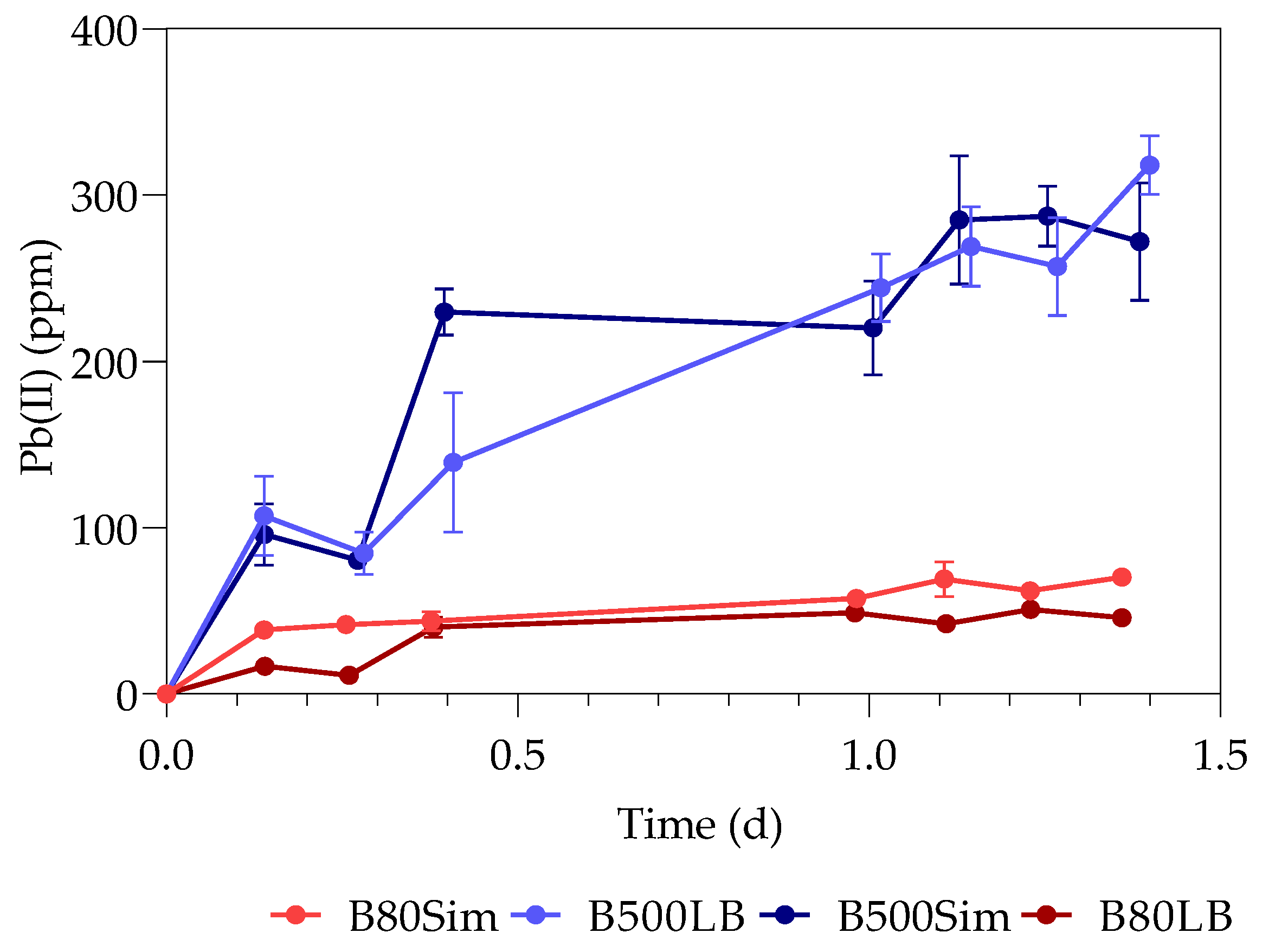
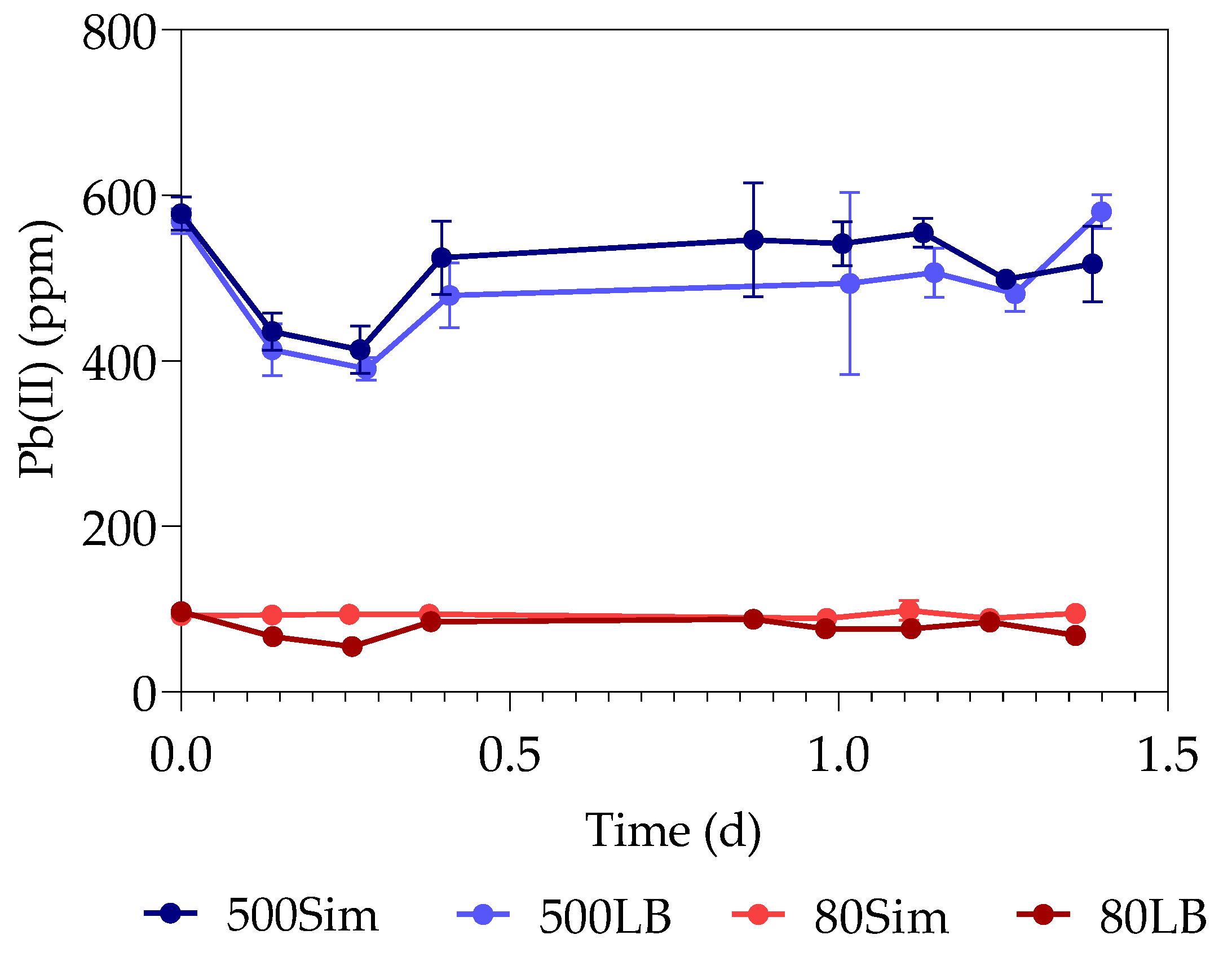
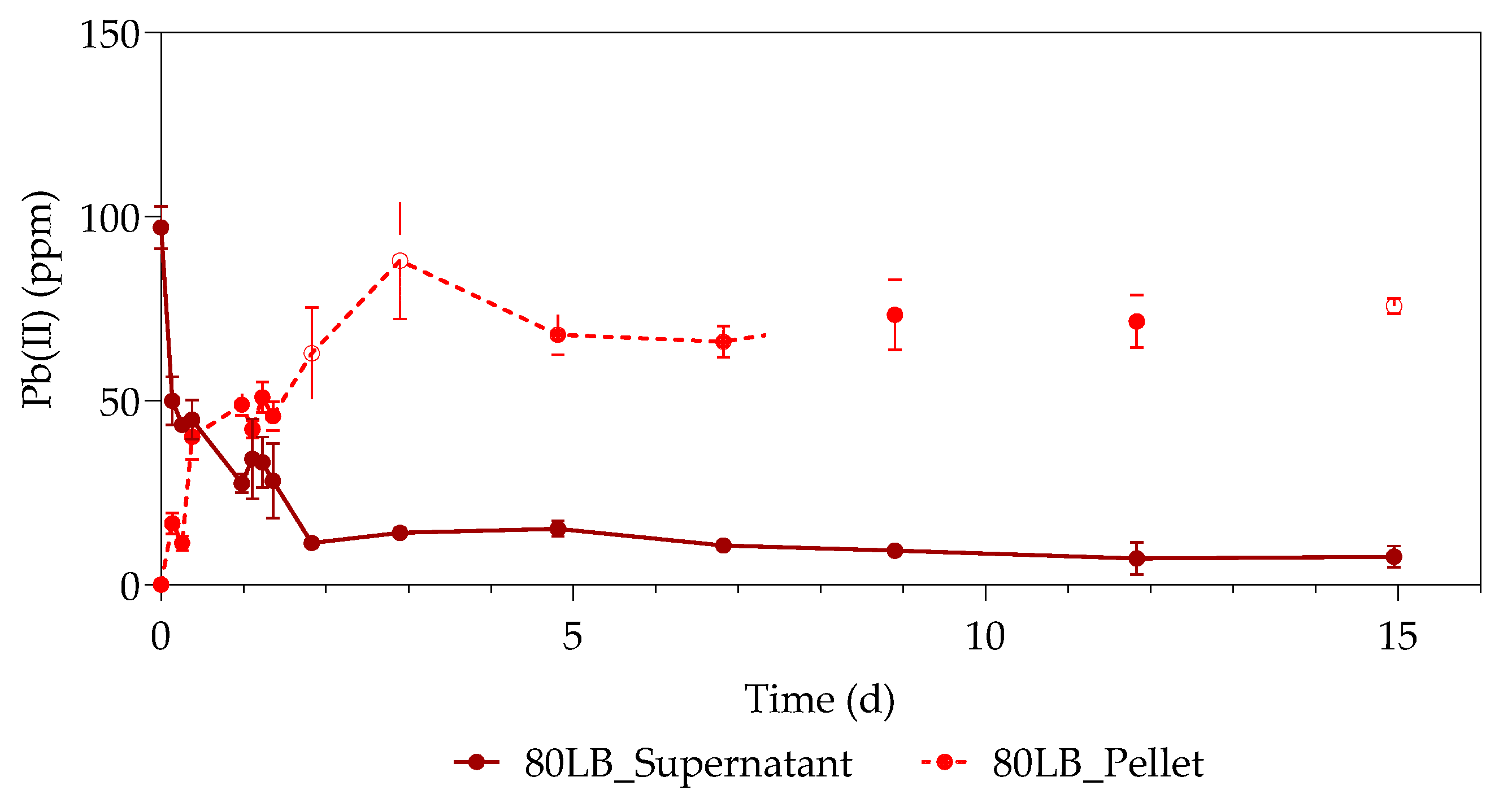
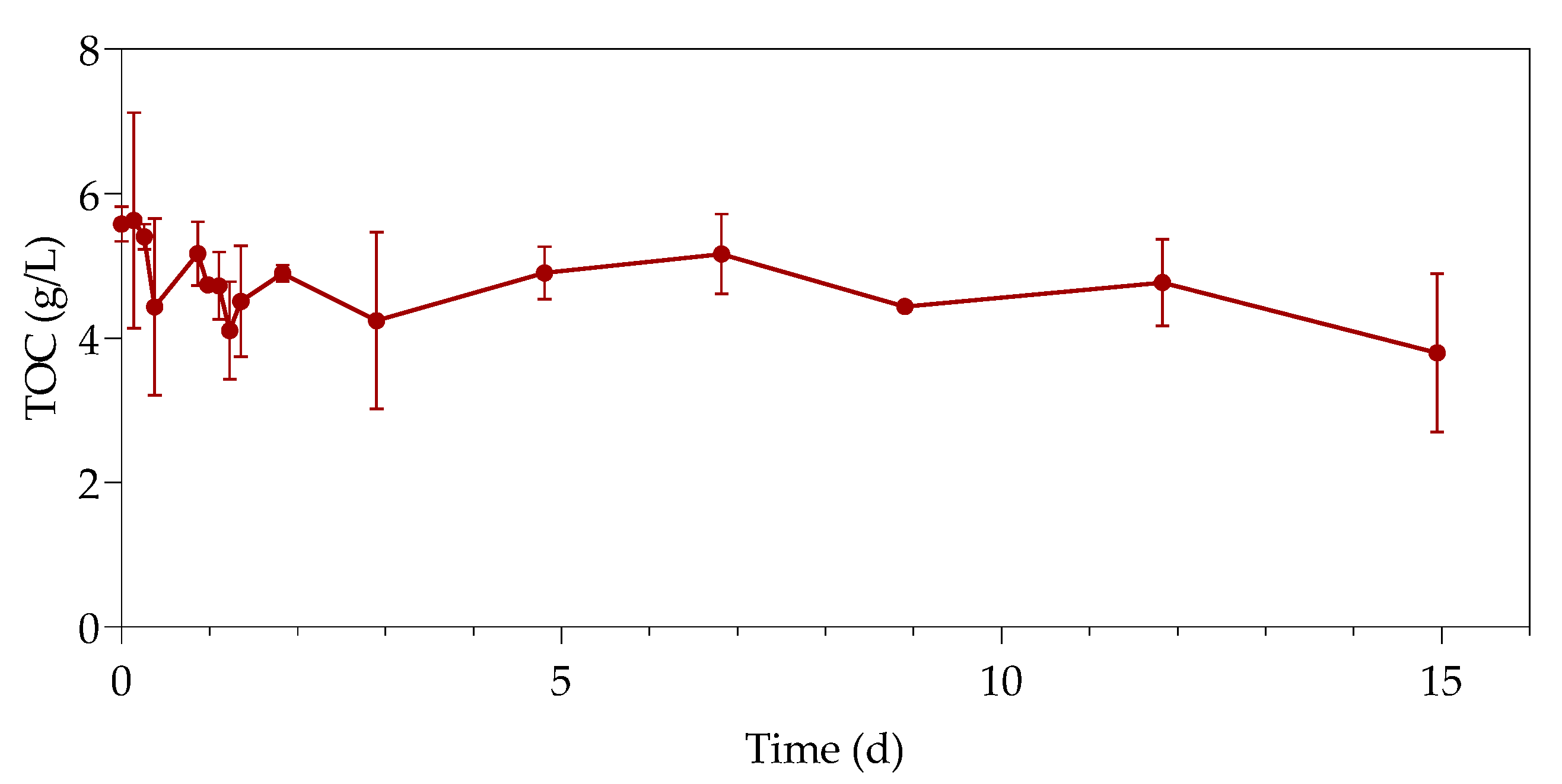
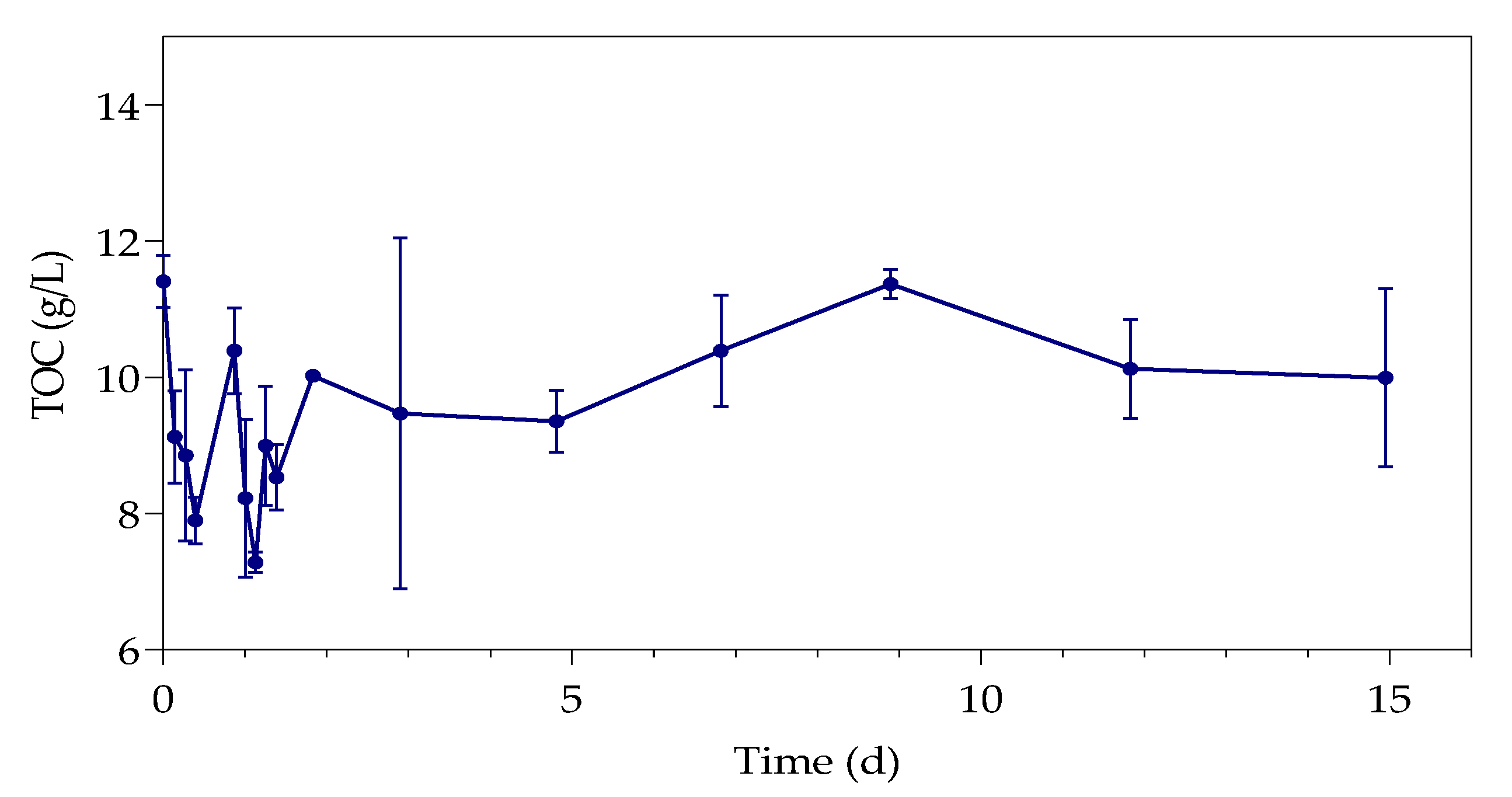
The first part of the study (all the datasets) was conducted over a period of 33 h, with samples taken every 3 h at 3, 6, 9, 24, 27, 30, and 33 h. The second part of the study (only samples containing 80 ppm with standard LB and 500 ppm with simulated LB) was extended for a duration of 15 days, taking samples every day from the 48 h onwards. The sealed serum bottles were shaken thoroughly before sampling, after which a hypodermic needle and sterile syringe were used to pierce the rubber stopper.
2.6. Batch Reactor Analysis
For each 1 mL sample, the residual aqueous Pb(II) in the sample supernatant and the Pb precipitated in the pellet were measured using an atomic absorption spectrometer (Perkin Elmer AAnalyst 400, Waltham, MS, USA) with a Pb Lumina hollow cathode lamp. The aqueous Pb(II) in the supernatant was directly analyzed from the respective 1 mL samples, while the precipitated pellet was digested in 0.1 mL 55% nitric acid (Glassworld, Johannesburg, South Africa) and 0.1 mL distilled water. Once the pellet was completely liquidized, water was added to a total volume of 1 mL dissolved sample and analyzed for Pb content.
Metabolic activity was quantified with a method using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) and spectrophotometric measurements at 550 nm. MTT is a water-based yellow dye that can be reduce to water-insoluble purple formazan crystals by dehydrogenase enzymes in viable cells. The crystals were then extracted with the aid of the organic solvent DMSO. Metabolic activity measurements were performed as soon as possible after sampling and quantified spectrophotometrically at 550 nm [
13]. Two sets of analyses were performed—one with biomass and the other without—to account for background interference and medium interaction with the MTT and DMSO. The samples were filtered with 25 mm nylon syringe filters with 0.45 μm pores (Anatech, Randburg, South Africa), to be recorded as the samples without biomass. The rest of the analysis was conducted by diluting the sample (with or without biomass), adding MTT, and finally incubated for precisely one hour at 35°C. The samples were then extracted with DMSO. The absorbance at 550 nm of the DMSO solution was measured and used as an indication of metabolic activity and, therefore, growth.
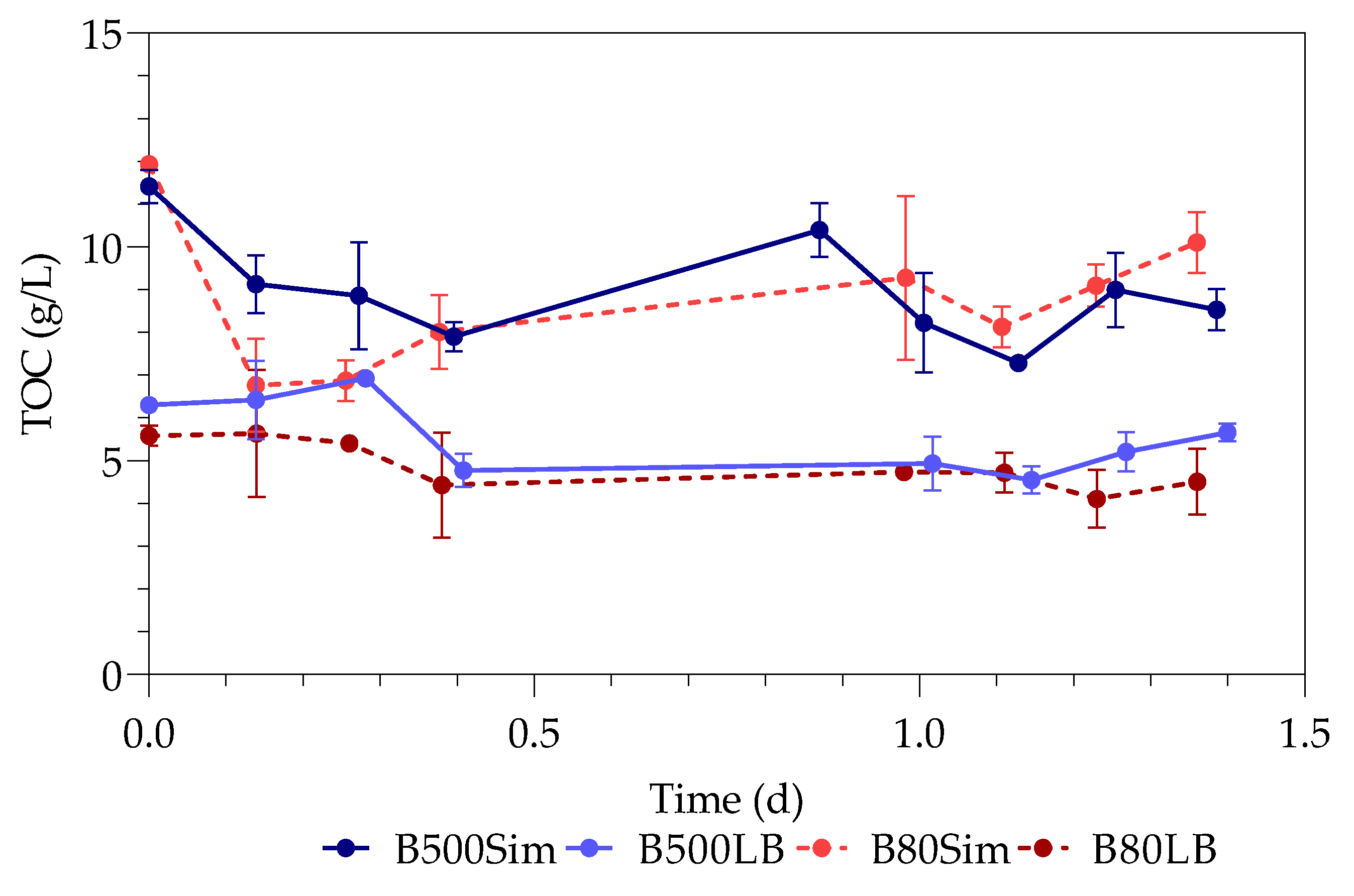
The supernatants and digested pellets of the samples were analyzed by using the TOC-V (option for liquid samples) on the Total Organic Carbon Analyzer (SHIMADZY, Kyoto, Japan) to test for their total organic carbon (TOC) content. The liquid samples (supernatant and digested pellet) were diluted, placed in 30 mL sample vials, and tested on the autosampler. Sodium persulfate and phosphoric acid were used by the instrument as oxidizers, with nitrogen as carrier gas. A method was created with a calibration curve using sucrose (Merck, Darmstadt, Germany).
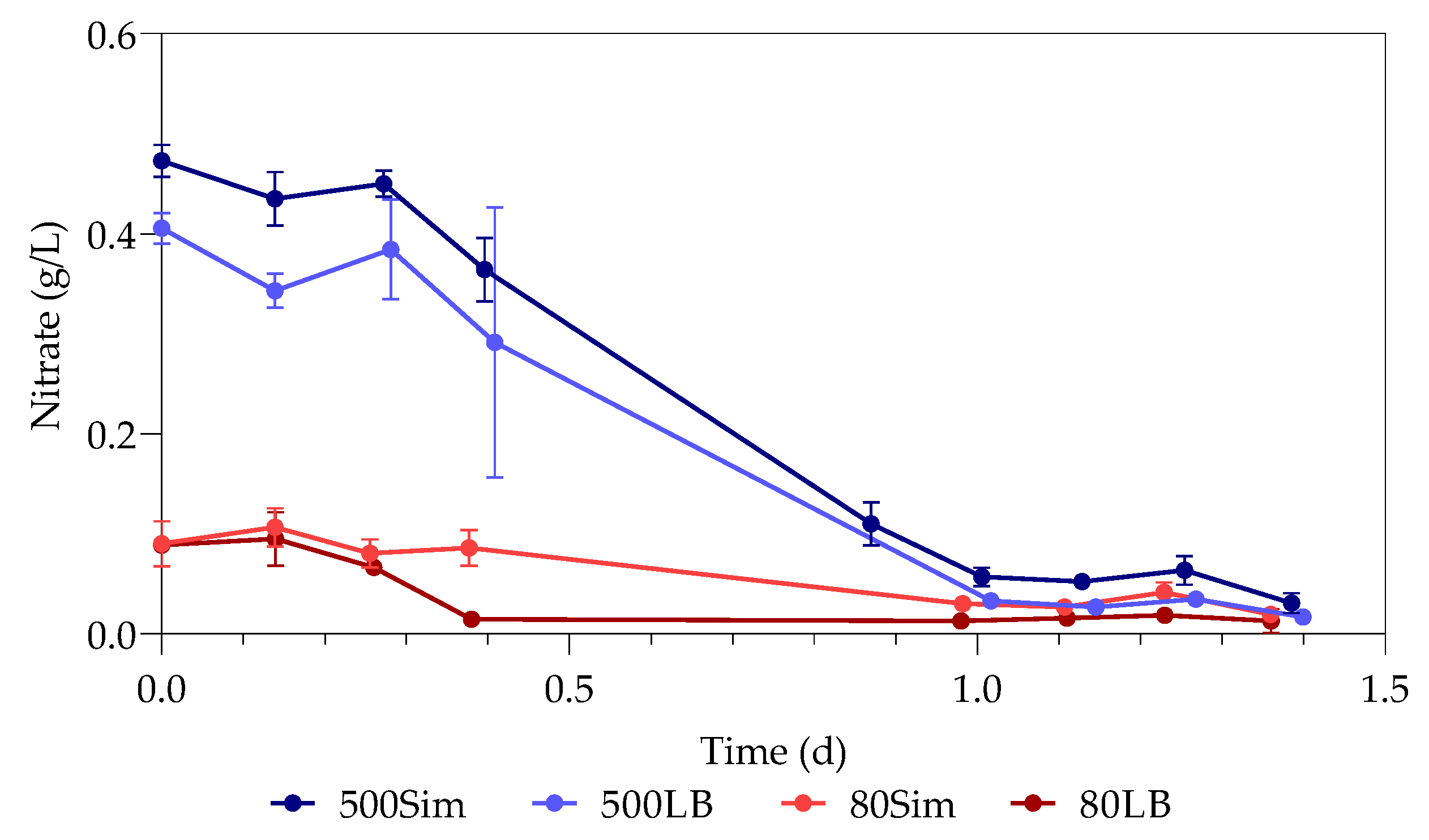
The samples used for nitrate testing were stored and measured at a later stage with a nitrate testing kit. Nitrate ions reacted with a form of benzoic acid in sulfuric acid to form a red nitro solution that was measured photometrically using the Spectroquant Nova 600 (Merck, Darmstadt, Germany).
2.7. Microbial Plate Preparation for Characterization
Spread and streak plates were prepared for the purpose of microbial characterization. A simulated LB agar was made and contained 20 g/L tryptone, 10 g/L yeast extract, 1 g/L NaCl, and 15 g/L agar (Sigma Aldrich, St Louis, MO, USA) as opposed to standard LB agar that contained 25 g/L LB (10 g/L tryptone, 5 g/L yeast extract, 10 g/L NaCl) and 15 g/L agar. The agar and Pb(II) stock solutions were autoclaved separately. The molten agar solutions were subsequently spiked with Pb(II) to produce 80 and 500 ppm concentrations. The agar was then poured into sterile petri dishes and left to solidify.
The serially diluted spread plates were prepared using the appropriate pre-cultures. The pre-cultures were diluted 4 times serially, each time diluting the previous concentrate 10 times with sterilized distilled water. The plates were evenly spread with a sterilized glass rod after adding 0.1 mL of the final dilution on each plate.
Streak plates were also prepared by dipping a sterile inoculation loop in the relevant pre-cultures and streaking it across the plate. Once all the plates were prepared, they were sealed with parafilm (Bemis, Oshkosh, WI, USA) and placed upside down in an airtight glass jar with an AneroGenTM sachet (Oxoid, Thermo Scientific, Basingstoke, Hampshire, UK), to ensure anaerobic conditions. The jars were incubated at 35 °C for seven days to allow for sufficient growth, followed by analysis.
2.8. Microbial Characterization
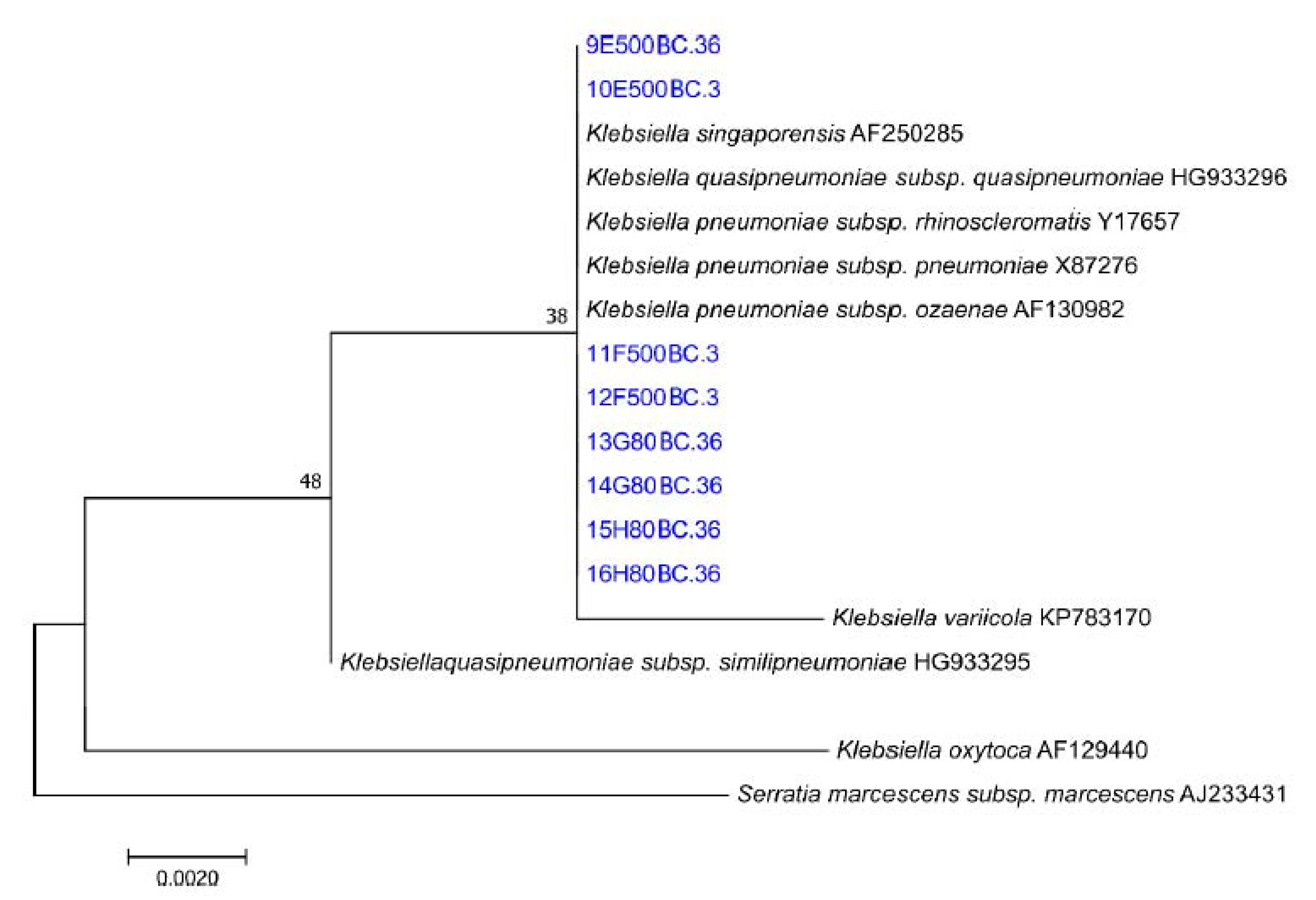
Four colonies were identified on each spread plate: two that represented the dominant color (dark brown/black) and shape of the overall colonies present and two that appeared to differ from the rest. Each selected colony was isolated and cultivated. The strain identification was based on the ±700 bp partial sequence of the 16S rRNA gene of the organisms. The sequences were compared against the GenBank of the National Center for Biotechnology in the United States of America with the aid of a basic BLAST (Basic Local Alignment Search Tool) search. The phylogenetic neighbors were obtained using the BLAST program against a database of type strains of valid published prokaryotic names (
http://www.ezbiocloud.net/) for each isolated sample.
Streak plate analysis involved trimming data and only using >q20 reads. Every read was BLASTED, and the data saved. The top hit for every BLAST result was recorded and noted of how many times each species appeared as a hit. The read count is the measurement of the number of reads that matched the relevant organism name.
2.9. SEM Analysis
The morphology of the precipitate was investigated using scanning electron microscopy (SEM) and X-ray photoelectron spectroscopy (XPS). The main advantages of these methods are that the precipitate can remain in its original form for analysis, and only a small amount of precipitate sample is required to complete the analysis.
Sample Preparation and Analysis
The appropriate samples were centrifuged at 9000 rpm and 20 °C for 10 min. The precipitate was centrifuged and washed 6 times. The samples were subsequently dried in anaerobic sterile glass jars with an anaerobic indicator (Oxoid, Thermo Scientific, Basingstoke, Hampshire, UK) and AneroGenTM sachet (Oxoid, Thermo Scientific, Basingstoke, Hampshire, UK) in the presence of silica gel. The jars were subsequently sealed and left to dry overnight. The samples were analyzed using a Zeiss Ultra PLUS FEG scanning electron microscope (SEM).
4. Discussion
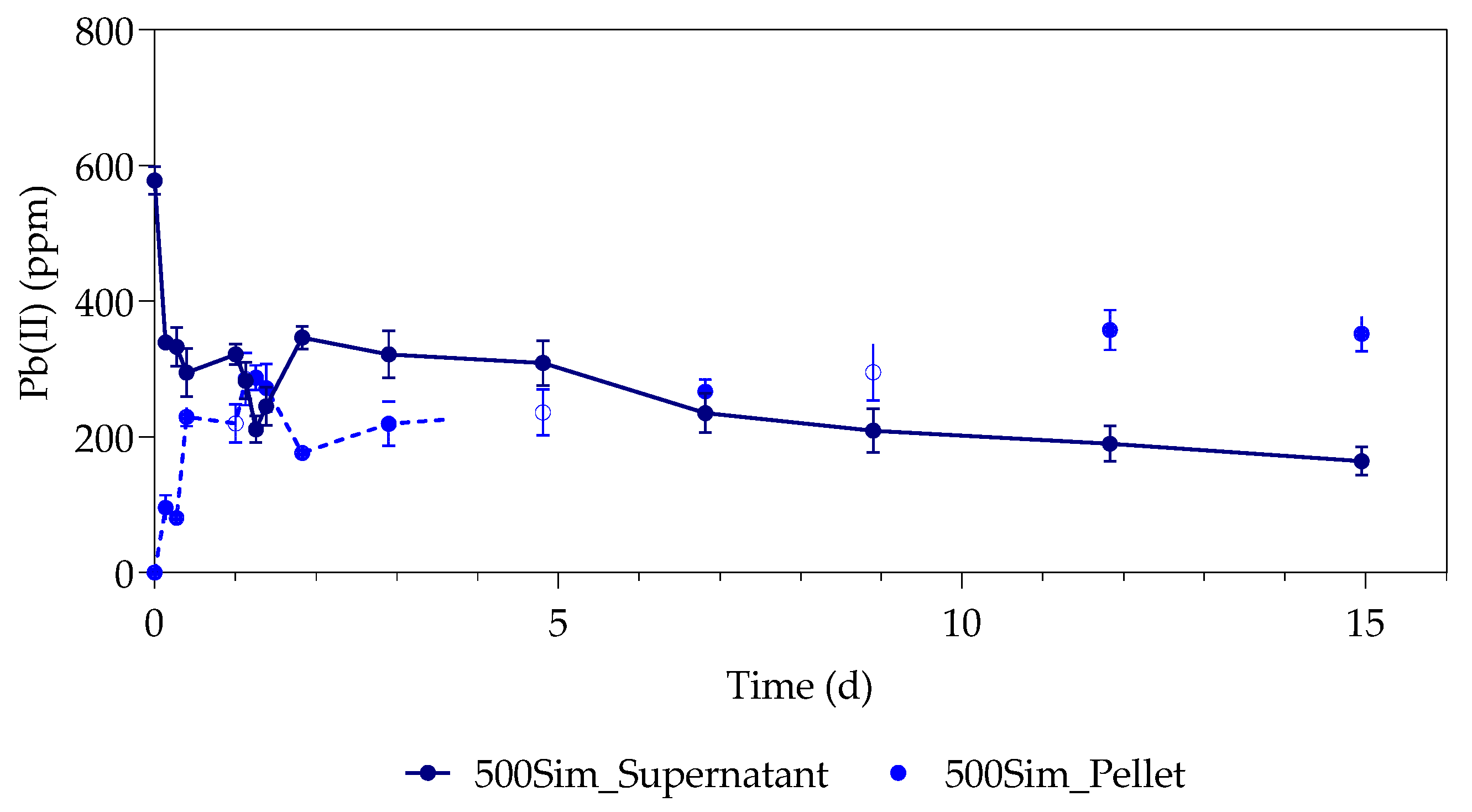
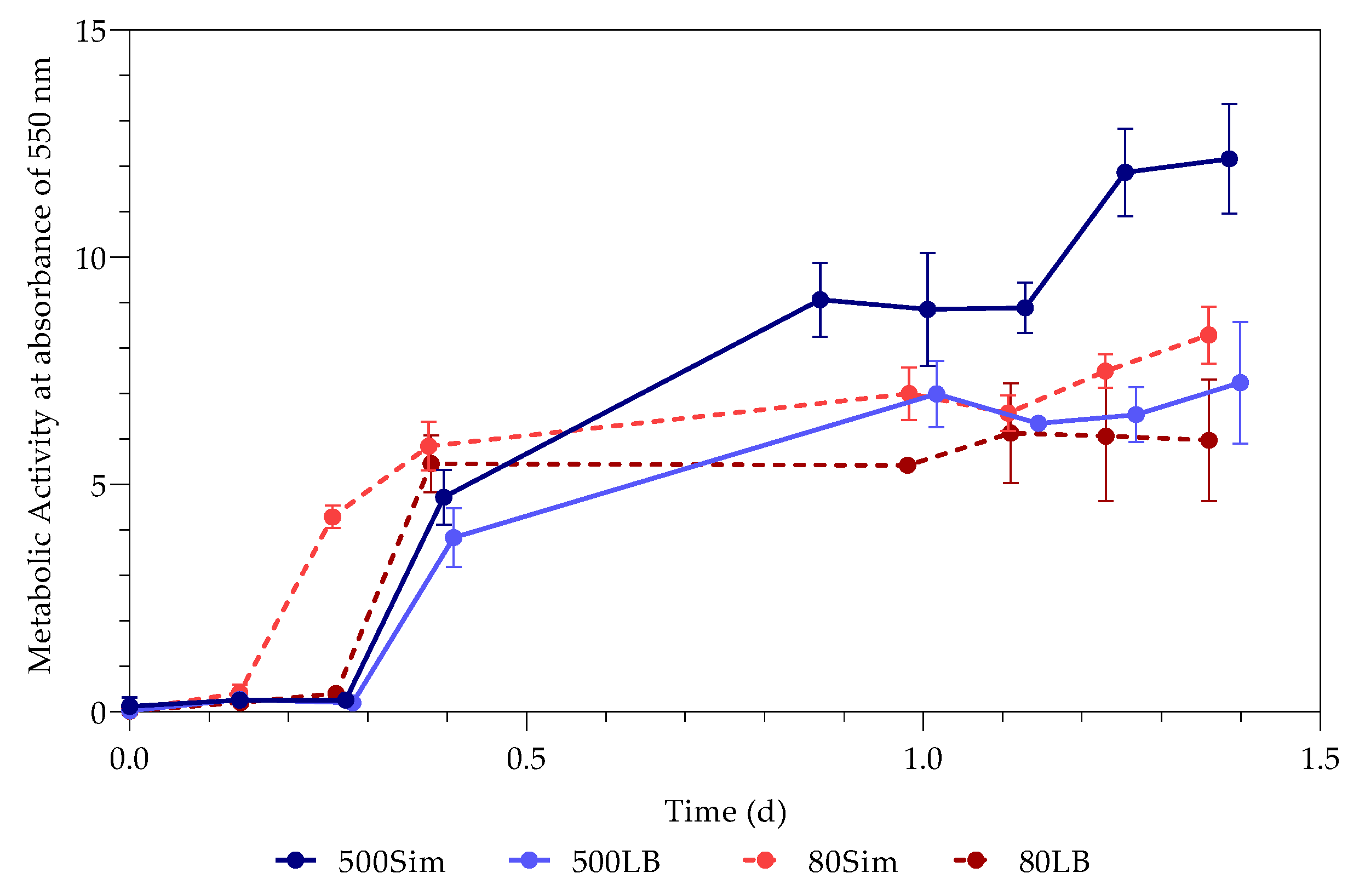
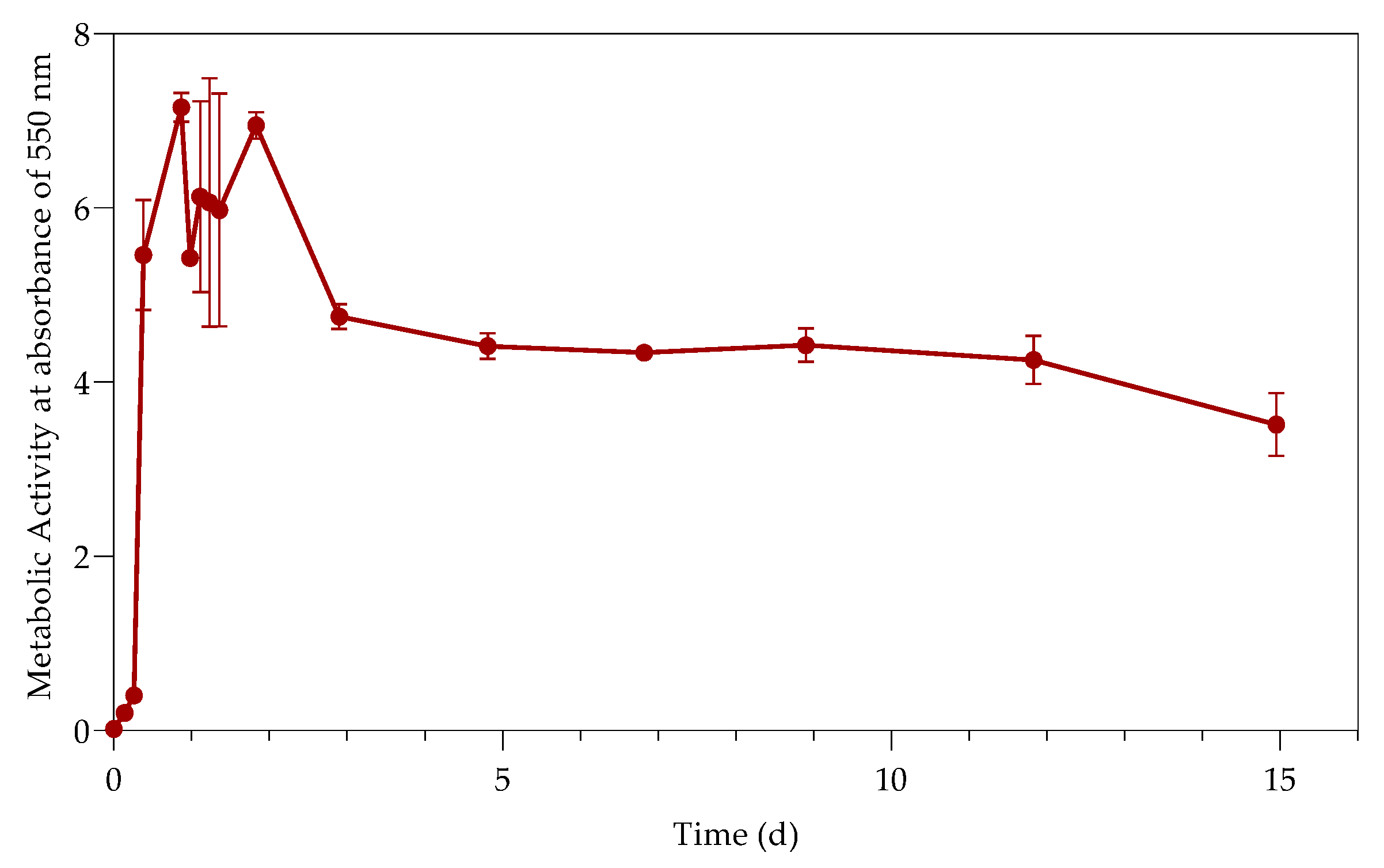
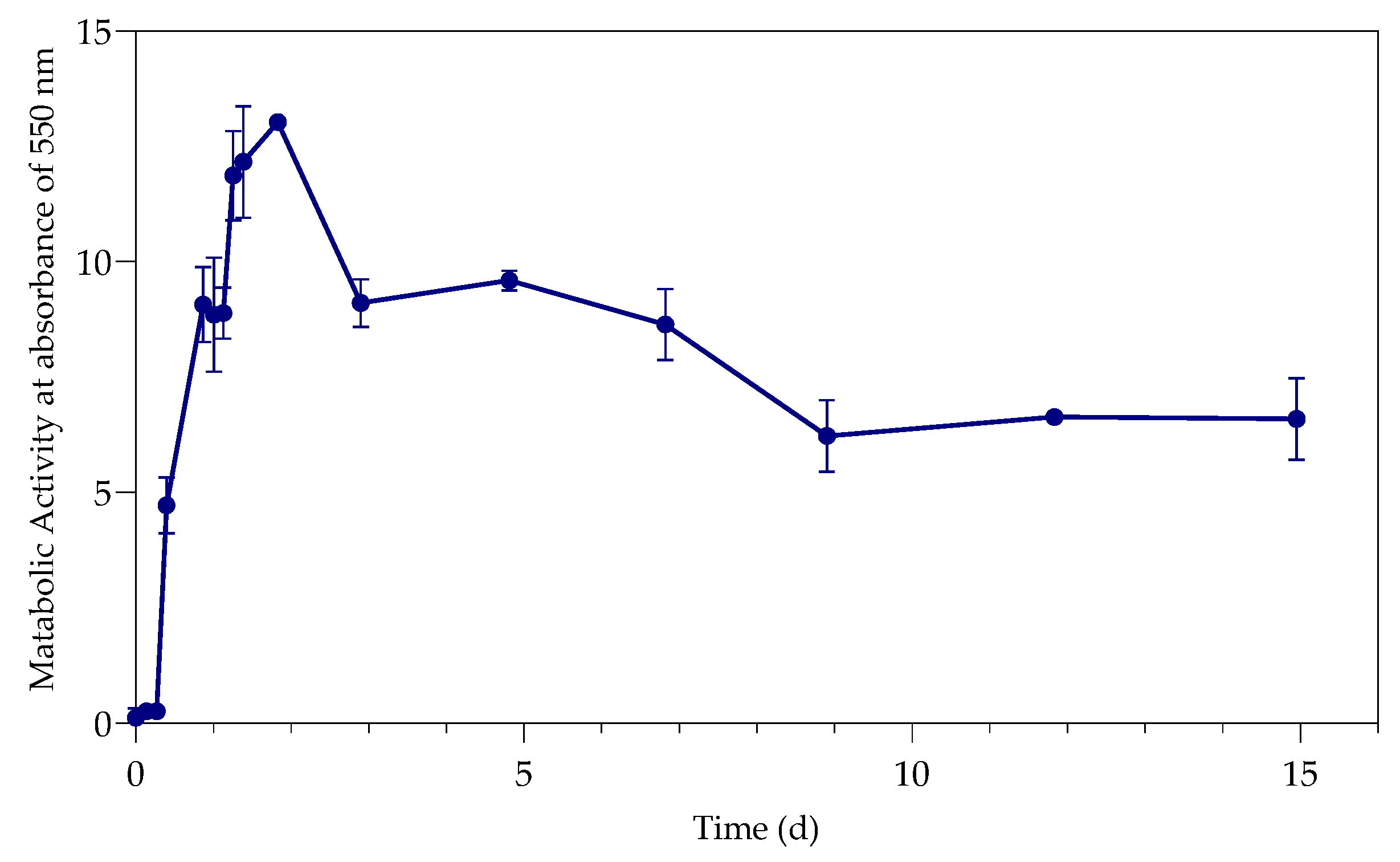
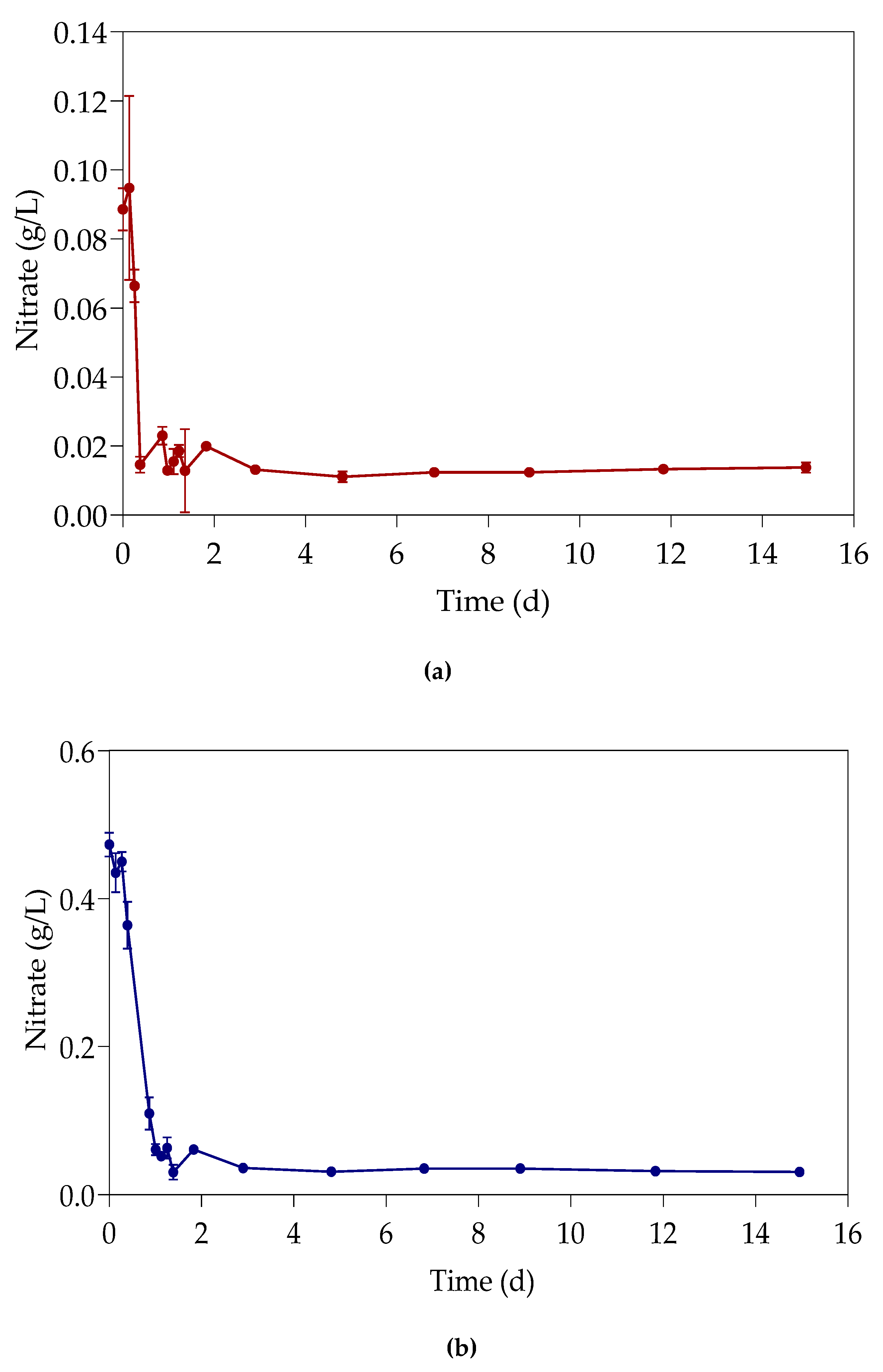
The study investigated the effect of Pb(II) and nutrient concentrations on Pb(II) bio-removal, microbial growth, active species, and population distribution of an industrially obtained Pb(II)-resistant consortium. The results demonstrated that the removal of Pb(II) from solution was independent of microbial metabolic activity and nutrient availability; the majority of Pb(II) removal (≈50%) took place during the first 3 h in the absence of observable changes in reactor medium, indicating a biosorption mechanism present. Metabolic activity was dependent on the amount of nitrate available, and a lag in initial growth may indicate a detoxification mechanism at work during the initial few hours of rapid Pb(II) removal. A small drop in total organic carbon indicates that substrate depletion was not a contributing factor to any growth changes.
The consumption of nitrates during initial exponential growth provides evidence for anaerobic respiration/denitrification present. Anaerobic respiration is the metabolic process during which microbes transfer electrons to terminal electron acceptors other than oxygen. It is crucial when the bacteria require the removal of surplus electrons to maintain an internal redox balance in the microbial cell. Compounds such as nitrate or metals (such as Pb(II)) have been used as electron acceptors for oxidizing NAD(P)H to NAD(P)
+ [
20]. It is proposed that nitrates are used as alternative electron acceptors in the absence of oxygen, during the first few hours of exponential growth, where the nitrate levels presented a decrease coupled with a growth increase. Sulphur is released at the same time during denitrification with the enzyme nitrate reductase from cysteine and methionine [
21], two amino acids present in the substrate, which in turns binds with Pb(II) to form PbS. When the nitrates are depleted a second alternative electron sink is required. It is highly likely that the remaining Pb(II) was then converted to Pb, as observed in a previous study conducted and published by this team [
9]. The exact mechanism should, however, be verified in future investigations as a conclusive answer could not yet be drawn.
The presence of various species in both the 80 and 500 ppm samples indicated a diverse consortium with Pb-resistant capabilities; however, the results from the spread-plate experiment evidence that Klebsiella pneumoniae was the dominant species responsible for Pb(II) precipitation. The dominance of the effective Pb biosorbent Ralstonia solanacearum species at 500 ppm Pb provides credence to the hypothesis that a biosorption mechanism is responsible for the initial Pb(II) removal observed in the initial 3 h. Preliminary unpublished experiments conducted by this team found that the consortium of bacteria performs better at Pb(II) removal as opposed to a refined single species, but conclusive results are required, and further studies will be implemented into the investigation of a synergistic co-existence between various microbial entities present during Pb(II) removal.
The specific growth rates were calculated under the various conditions with Pb(II) inhibition. It was found that the overall generation time for the consortium was comparatively quick. The rapid rate of generation was attributed to various microbial entities present in the sample, each with different generation times and growth rates.
It was concluded from the morphology studies that viable bacterial cells are embedded in abundance in the precipitate. This indicated a precipitation mechanism rather than an external excretion mechanism.
The findings from this study elucidate the possible mechanisms of Pb(II) removal and subsequent precipitation and will be implemented in conjunction with a proposed kinetic model to develop a design and implementation strategy for continuous bio removal and recovery of Pb(II) from industrial effluents.


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

























