Sustainable Weed Control and Enhancing Nutrient Use Efficiency in Crops through Brassica (Brassica compestris L.) Allelopathy

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
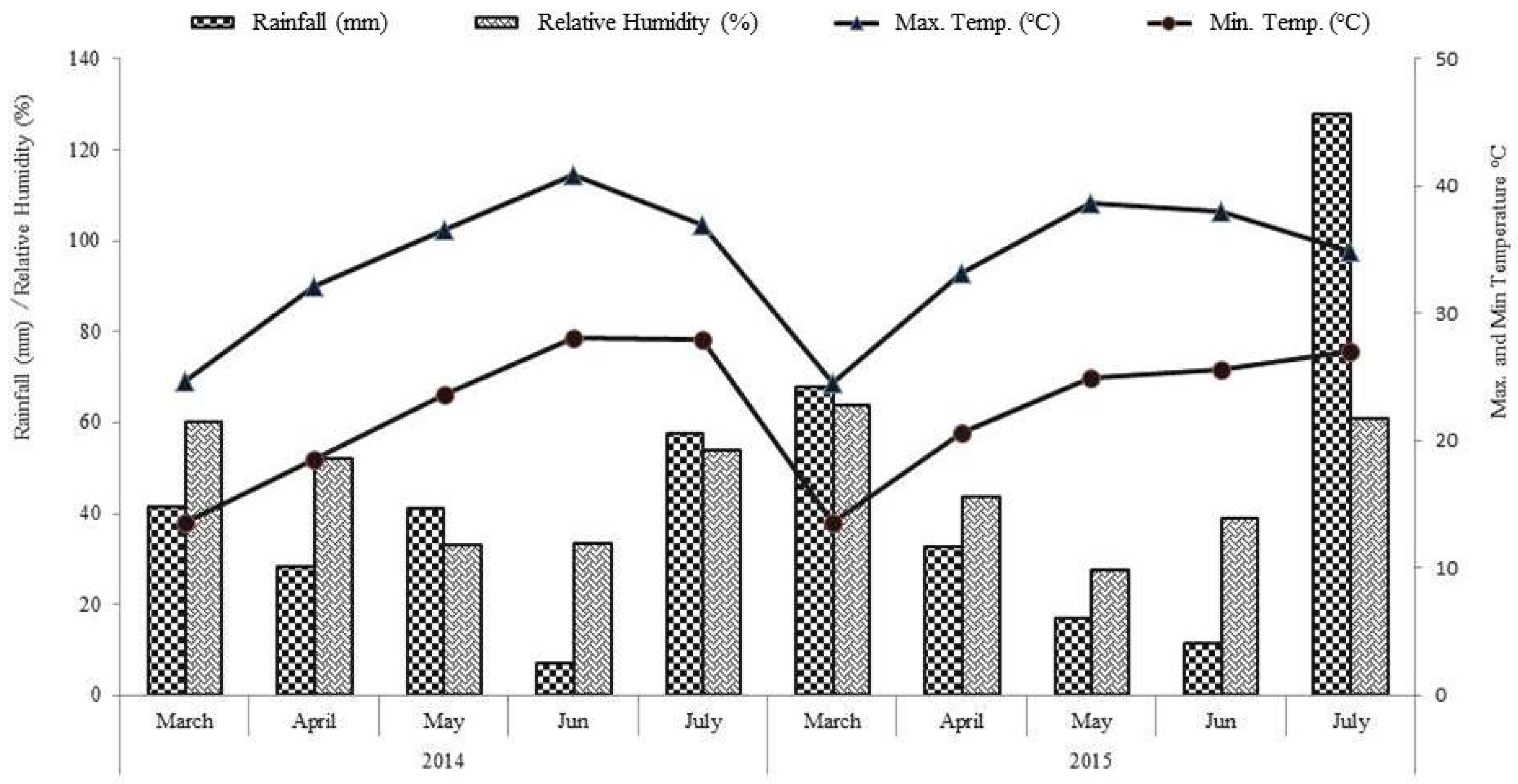
2.1. Soil, Site, and Climate
2.2. Experimental Design
2.3. Land Preparation and Crop Management
2.4. Brassica Crop Water Extract Preparation
2.5. Brassica Crop Residue Preparation
2.6. Soil Sampling
2.7. Observations, Measurements, and Data Analysis
3. Results
3.1. Physical and Chemical Indicators of Soil Health
3.2. Microbiological and Biochemical Indicators of Soil Health
3.3. Weed Dynamics
3.4. Yield Components
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tanentzap, A.J.; Lamb, A.; Walker, S.; Farmer, A. Resolving conflicts between agriculture and the natural environment. PLoS Biol. 2015, 13, 23–45. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Rauf, H.A.; Shah, A.N.; Shahzad, B.; Bukhari, M.A. Allelopathic effects of rose wood, guava, eucalyptus, sacred fig and jaman leaf litter on growth and yield of wheat (Triticum aestivum L.) in a wheat-based agroforestry system. Planta Daninha 2017, 35, e017166992. [Google Scholar] [CrossRef] [Green Version]
- Gnanavel, I. Eco-Friendly Weed Control Options for Sustainable Agriculture. Sci. Int. 2015, 3, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Zahra, S.T.; Ahmad, M.; Shah, A.N.; Hassan, W. Herbicidal potential of dryland plants on growth and tuber sproutingin purple nutsedge (Cyperus rotundus). Planta Daninha 2018, 36, e018170606. [Google Scholar] [CrossRef]
- Duke, S.O.; Scheffler, B.E.; Dayan, F.E. Allelochemicals as herbicides. In Physiological Aspects of Allelopathy, Proceedings of the 1st European OECD Allelopathy Symposium, Vigo, Spain, 21–23 June 2001; Bonjoch, N.P., Reigosa, M.J., Eds.; Gamesal: Vigo, Spain, 2001; pp. 47–59. [Google Scholar]
- Fickett, N.D.; Boerboom, C.M.; Stoltenberg, D.E. Predicted corn yield loss due to weed competition prior to postemergence herbicide application on Wisconsin farms. Weed Technol. 2013, 27, 54–62. [Google Scholar] [CrossRef]
- Farooq, N.; Abbas, T.; Tanveer, A.; Jabran, K. Allelopathy for weed management. In Co-Evolution of Secondary Metabolites; Springer: Cham, Switzerland, 2020; pp. 505–519. [Google Scholar]
- Parween, T.; Jan, S.; Mahmooduzzafar, S.; Fatma, T.; Siddiqui, Z.H. Selective effect of pesticides on plant—A review. Crit. Rev. Food Sci. Nut. 2016, 56, 160–179. [Google Scholar] [CrossRef] [PubMed]
- The Express Tribune. India Fails to Substantiate Claim of Pakistan’s Involvement in Uri Attack. 2017. Available online: https://tribune.com.pk/story/1339596/india-fails-substantiate-claim-pakistans-involvement-uri-attack (accessed on 26 February 2017).
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Shah, A.D.; Iqbal, J.; Ullah, A.; Yang, G.; Yousaf, M.; Fahad, S.; Tanveer, M.; Hassan, W.; Tung, S.A.; Wang, L.; et al. Allelopathic potential of oil seed crops in production of crops: A review. Environ. Sci. Pollut. Res. 2016, 23, 14854–14867. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.D.; Iqbal, J.; Fahad, S.; Tanveer, M.; Yang, G.; Khan, E.A.; Shahzad, B.; Yousaf, M.; Hassan, W.; Ullah, A.; et al. Allelopathic influence of sesame and green gram intercrops on cotton in a replacement series. Clean Soil Air Water 2017, 45, e1500469. [Google Scholar] [CrossRef]
- Weston, L.A.; Duke, S.O. Weed and crop allelopathy. Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Alsaadawi, I.S.; Dayan, F.E. Potentials and prospects of sorghum allelopathy in agro ecosystems. Allelop. J. 2009, 24, 255–270. [Google Scholar]
- Takeshita, V.; Mendes, K.F.; Alonso, F.G.; Tornisielo, V.L. Effect of Organic Matter on the Behavior and Control Effectiveness of Herbicides in Soil. Planta Daninha 2019, 37, e019214401. [Google Scholar] [CrossRef] [Green Version]
- Alsaadawi, I.S.; Al-Ekeelie, M.H.S.; Al-Hamzawi, M.K. Differential allelopathic potential of grain sorghum genotypes to weeds. Allelop. J. 2007, 19, 153–160. [Google Scholar]
- Weston, L.A.; Alsaadawi, I.S.; Bearson, S.R. Sorghum allelopathy from ecosystem to molecule. J. Chem. Ecol. 2013, 8, 34–45. [Google Scholar] [CrossRef]
- Siemens, D.H.; Garner, S.H.; Mitchell-Olds, T.; Callaway, R.M. Cost of defense in the context of plant competition: Brassica rapa may grow and defend. Ecology 2002, 83, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Bajwa, A.A.; Cheema, S.A.; Cheema, Z.A. Application of Allelopathy in Crop Production. Int. J. Agric. Biol. 2013, 6, 1367–1378. [Google Scholar]
- Turk, M.A.; Tawaha, A.M. Inhibitory effects of aqueous extracts of black mustard on germination and growth of lentil. Pak. J. Agron. 2002, 1, 28–30. [Google Scholar]
- Turk, M.A.; Tawaha, A.M. Allelopathy effect of black mustard (Brassica nigra. L.) on germination and growth of wild oat. Crop. Prot. 2003, 22, 673–677. [Google Scholar] [CrossRef]
- Turk, M.A.; Lee, K.D.; Tawaha, A.M. Inhibitory effects of aqueous extracts of black mustard on germination and growth of radish. Res. J. Agric. Biol. Sci. 2005, 1, 227–231. [Google Scholar]
- Smith, A.N.; Reberg-Horton, S.C.; Place, G.T.; Meijer, A.D.; Arellano, C.; Mueller, J.P. Rolled rye mulch for weed suppression in organic no-tillage soybeans. Weed Sci. 2011, 59, 224–231. [Google Scholar] [CrossRef]
- Narwal, S.S.; Sati, S.C.; Palaniraj, R. Allelopathic weed suppression of brassica accessions against major winter weeds in north India. In Proceedings of the Second European Allelopathy Symposium “Allelopathy—From Understanding to Application”, Pulawy, Poland, 3–5 June 2004. [Google Scholar]
- Vyvyan, J.R. Allelochemicals as leads for new herbicides and agrochemicals. Tetrahedron 2002, 58, 1631–1646. [Google Scholar] [CrossRef]
- Younesabadi, M. Study of allelopathic interference of rapeseed (Brassica napus) Var. Belinda on germination and growth of cotton (Gossypium hirsutum) and itsdominant weeds. In Proceedings of the Forth World Congress on Allelopathy, “Establishing the Scientific Base”, Wagga Wagga, New South Wales, Australia, 21–26 August 2005; Charles Sturt University: Bathurst, New South Wales, Australia; pp. 283–286. [Google Scholar]
- Wu, F.; Wang, X.; Xue, C. Effect of cinnamic acid on soil microbial characteristics in the cucumber rhizosphere. Eur. J. Soil Biol. 2009, 45, 356–362. [Google Scholar] [CrossRef]
- Kong, C.H.; Wang, P.; Zhao, H.; Xu, X.H.; Zhu, Y.D. Impact of allelochemical exuded from allelopathic rice on soil microbial community. Soil Biol. Biochem. 2008, 40, 1862–1869. [Google Scholar] [CrossRef]
- USDA-NRCS. Soil Health Assessment, Soil Survey Staff; NRCS: Washington, DC, USA. Available online: http://www.nrcs.usda.gov/wps/portal/nrcs/main/soils/health/ (accessed on 27 November 2014).
- Bertin, C.; Yang, X.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant. Soil. 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Geng, G.D.; Zhang, S.Q.; Cheng, Z.H. Effects of different allelochemicals on mineral elements absorption of tomato root. China Veget. 2009, 4, 48–51. [Google Scholar]
- Jónsson, J.Ö.G. Soil: Ecosystem Services, Economic Valuation and Sustainability Indicators. Ph.D. Thesis, Faculty of Life and Environmental Sciences, University of Iceland, Reykjavik, Island, 2019. [Google Scholar]
- Singh, B.; Shan, Y.H.; Johnson-beeebout, S.E.; Singh, Y.; Buresh, R.J. Crop residue management for lowland rice-based cropping systems in Asia. Adv. Agron. 2008, 98, 118–199. [Google Scholar]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant. Soil. 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Jabran, K.; Cheema, Z.A.; Farooq, M.; Hussain, M. Lower doses of Pendimethalin mixed with allelopathic crop water extracts for weed management in canola (Brassica napus). Int. J. Agric. Bio. 2010, 12, 335–340. [Google Scholar]
- Blake, G.R.; Hartge, K.H. Bulk density. In Methods of Soil Analysis: Part 1. Physical and Mineroalogical Methods. Agronomy Monograph No. 9, 2nd ed.; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; pp. 363–375. [Google Scholar]
- Vomocil, J.A. Porosity. In Methods of Soil Analysis; Blake, C.A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 299–314. [Google Scholar]
- Ryan, J.; Estefan, G.; Rashid, A. Soil and Plant Analysis: Laboratory Manual; International Center for Agricultural Research in Dry Areas (ICARDA): Aleppo, Syria; Nacional Agricultural Research Centre: Islamabad, Pakistan, 2001; p. 172.
- U.S. Salinity Laboratory Staff. Diagnosis and Improvement of Saline and Alkali Soils. USDA Agriculture Handbook, 60; Richards, L.A., Ed.; U.S. Government Printing Office: Washington, DC, USA, 1954.
- Bremner, J.M.; Mulvaney, C.S. Total nitrogen. In Methods of Soil Analysis; Page, A.L., Miller, R.H., Keeny, D.R., Eds.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1982; pp. 1119–1123. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Olsen, S.O.; Sommers, I.E. Phosphorus. In Methods of Soil Analysis. Chemical and Microbial Properties: Part 2, 2nd ed.; Page, A.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Helmke, P.A.; Sparks, D.L. Lithium, sodium and potassium, rubidium and cesium. In Methods of Soil Analysis; American Society of Agronomy: Madison, WI, USA, 1996; pp. 551–575. [Google Scholar]
- Aslam, Z.; Yasir, M.; Jeon, C.O.; Chung, Y.R. Lysobacter oryzae sp. nov., isolated from the rhizosphere of rice (Oryza sativa L.) managed under no-tillage practice. Int. J. Syst. Evol. Microbiol. 2008, 59, 675–680. [Google Scholar] [CrossRef]
- Janssen, P.H.; Yates, P.S.; Grinton, B.E.; Taylor, P.M.; Sait, M. Improved culturability of soil bacteria and isolation in pure culture of novel members of the divisions Acidobacteria, Actinobacteria, Proteobacteria, and Verrucomicrobia. Appl. Environ. Microbiol. 2002, 68, 2391–2396. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.X.; Ye, Q.F.; Min, H.; Duan, X.J.; Jin, W.M. Bt-transgenic rice straw affects the culturable micro biota and dehydrogenase and phosphatase activities in a flooded paddy soil. Soil Biol. Biochem. 2004, 36, 289–295. [Google Scholar]
- Martin, J.P. Use of acid, rose bengal and streptomycin in the plate method for enumerating soil fungi. Soil Sci. 1950, 69, 215–232. [Google Scholar] [CrossRef]
- Min, H.; Ye, Y.F.; Chen, Z.Y.; Wu, W.X.; Du, Y.F. Effects of butachlor on microbial populations and enzyme activities in paddy soil. J. Environ. Sci. Health 2001, 36, 581–595. [Google Scholar] [CrossRef] [PubMed]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- International Maize and Wheat Improvement Center (CIMMYT). From Agronomic Data to Farmers Recommendations: An Economics Training Manual. No. 27; CIMMYT: Texcoco, Mexico, 1988. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biometric Approach, 3rd ed.; McGraw Hill Book Company: New York, NY, USA, 1996. [Google Scholar]
- Petersen, J.R.; Belz, F.W.; Hurle, K. Weed suppression by release of isothiocyanates from turnip-rape mulch. Agron. J. 2001, 93, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Cheema, Z.A.; Khaliq, A.; Jabran, K.; Mushtaq, M.N. Weed control in wheat through combined application of two allelopathic crop water extracts with lower dose of atlantis. In Proceedings of the 8th National Weed Science Conference, Government College University, Lahore, Pakistan, 25–27 June 2007. [Google Scholar]
- Scavo, A.; Mauromicale, G. Integrated Weed Management in Herbaceous Field Crops. Agronomy 2020, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Jafariehyazdi, E.; Javidfar, F. Comparison of allelopathic effects of some brassica species in two growth stages on germination and growth of sunflower. Plant. Soil Envrion. 2011, 57, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Norsworthy, J.K.; Malik, M.S.; Jha, P.; Oliveira, M.J. Effects of isothiocyanates on purple (Cyperus rotundus L.) and yellow nutsedge (Cyperus esculentus L.). Weed Biol. Manag. 2006, 6, 131–138. [Google Scholar] [CrossRef]
- Arsalan, M.; Uremis, I.; Uludag, A. Determining bio-herbicidal potential of rapeseed, radish and turnip extracts on germination inhibition of cut leaf ground-cherry (Physalis angulata L.) seeds. J. Agron. 2005, 4, 134–137. [Google Scholar]
- Cutillo, F.; D’Abrosca, B.; Della Greca, M.; Di Marino, C.; Golino, A.; Previtera, L.; Zarrelli, A. Cinnamic acid amides from Chenopodium album: Effects on seed germination and plant growth. Phytochemistry 2003, 64, 1381–1387. [Google Scholar] [CrossRef]
- Narwal, S.S.; Sati, S.C.; Palaniraj-Singh, H.R.; Rawat, L.S. Weed Management Potential of Brassica Accessions on Major Winter Weeds. In Proceedings of the Abstracts, III World Congress on Allelopathy Challenge for the New Millennium, Tsukuba, Japan, 26–30 August 2002; Fujii, Y., Hiradata, S., Araya, H., Eds.; Japanese Weed Science Society: Tsukuba, Japan; p. 146. [Google Scholar]
- Norsworthy, J.K.; Brandenberger, L.; Burgos, N.R.; Riley, M. Weed suppression in Vigna unguiculata with a spring-seeded brassicaceae green manure. Crop. Protec. 2005, 24, 441–447. [Google Scholar] [CrossRef]
- Shaver, T. Crop residue and soil physical properties. In Proceedings of the 22nd Annual Central Plains Irrigation Conference, Kearney, NE, USA, 24–25 February 2010. [Google Scholar]
- Scavo, A.; Carlos, R.; José, M.G.M.; Rosa, M.V.; Giovanni, M.; Francisco, A.M. The extraction procedure improves the allelopathic activity of cardoon (Cynara cardunculus var. altilis) leaf allelochemicals. Ind. Crop. Prod. 2019, 128, 479–487. [Google Scholar] [CrossRef]
- Sidhu, B.S.; Beri, V. Experience with managing rice residue in intensive rice-wheat cropping system in Punjab. In Conservation Agriculture-Status and Prospects; Abrol, I.P., Gupta, R.K., Malik, R.K., Eds.; Centre for Advancement of Sustainable Agriculture (CASA): New Delhi, India, 2005; pp. 55–63. [Google Scholar]
- Verhulst, N.; Nelissen, V.; Jespers, N.; Haven, H.; Sayre, K.D.; Raes, D.; Deckers, J.; Govaerts, B. Soil water content, maize yield and its stability as affected by tillage and crop residue management in rainfed semi-arid highlands. Plant. Soil. 2011, 124, 347–356. [Google Scholar] [CrossRef]
- Jin, Y.Q.; Du, B.J.; Gao, H.J.; Chang, J.; Zhang, L.G. Effects of maize straw returning on water dynamics and water use efficiency of winter wheat in lime concretion black soil. J. Triticeae Crops 2013, 33, 1–7. [Google Scholar]
- Zhou, J.; Xu, D.; Xue, C. Study of comprehensive utilization efficiency of returning rice straw to field. Chin. Agri. Sci. Bulli. 2002, 4, 3. [Google Scholar]
- Jabran, K.; Farooq, M.; Aziz, T.; Siddique, K.H.M. Allelopathy and crop nutrition. In Allelopathy: Current Trends and Future Applications; Cheema, Z.A., Farooq, M., Wahid, A., Eds.; Springer: Berlin, Germany, 2013; pp. 337–348. [Google Scholar]
- Beres, I.; Kazinczi, G. Allelopathic effects of shoot extracts and residues of weeds on field crops. Allelop. J. 2000, 7, 93–98. [Google Scholar]
- Sharma, M.P.; Bali, S.V.; Gupta, D.K. Crop yield and properties of Inceptisol as influenced by residue management under rice-wheat system. J. Indian Soc. Soil Sci. 2000, 48, 506–509. [Google Scholar]
- Gong, Z.; Li, P.; Wilke, B.M.; Alef, K. Effects of vegetable oil residue after soil extraction on physical-chemical properties of sandy soil and plant growth. J. Environ. Sci. 2008, 20, 1458–1462. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Pandher, J.K.; Arora, V.; Kohli, R.K. Phytotoxic effect of parthenium residues on the selected soil properties and growth of chickpea and radish. Weed Biol. Manag. 2002, 2, 73–78. [Google Scholar] [CrossRef]
- Batish, D.R.; Tung, P.; Singh, H.P.; Kohli, R.K. Phytotoxicity of sunflower residues against some summer season crops. J. Agron. Crop. Sci. 2002, 188, 19–24. [Google Scholar] [CrossRef]
- Dick, R.P. Soil Enzymatic Activities as in Indicator of Soil Quality, in Deinding Soil Quality for a Sustainable Development; Doran, J.W., Coleman, D.C., Bezdicek, D.F., Stewart, B.A., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 107–124. [Google Scholar]
- Doran, J.W. Microbial changes associated with residue management with reduce management with reduced tillage. Soil Sci. Soc. Am. J. 1980, 44, 518–524. [Google Scholar] [CrossRef]
- Dick, W.A.; Juma, N.G.; Tabatabai, M.A. Effects of soils on acid phosphatase and inorganic pyro phosphatase of corn roots (Zea mays). Soil Sci. 1983, 136, 19–25. [Google Scholar] [CrossRef]
- Doornbos, R.F.; van Loon, L.C.; Bakker, P.A.H.M. Impact of root exudates and plant defense signalling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 2012, 32, 227–243. [Google Scholar] [CrossRef]
- Bohme, L.; Langer, U.; Bohme, F. Microbial biomass, enzyme activities and microbial community structure in two European long-term field experiments. Agric. Ecosys. Environ. 2005, 109, 141–152. [Google Scholar] [CrossRef]
- Brzezinska, M.; Stepniewska, Z.; Stepniewski, W. Soil oxygen status and dehydrogenase activity. Soil Biol. Biochem. 1998, 30, 1783–1790. [Google Scholar] [CrossRef]
- Rial, C.; Gómez, E.; Varela, R.M.; Molinillo, J.M.G.; Macías, F.A. Ecological relevance of the major allelochemicals in Lycopersicon esculentum roots and exudates. J. Agric. Food Chem. 2018, 66, 4638–4644. [Google Scholar] [CrossRef]
- Ravankar, H.N.; Patil, R.; Puranik, R.B. Decomposition of different organic residues in soil. PKV Res. J. 2000, 24, 23–25. [Google Scholar]
- Farooq, M.; Jabran, K.; Rehman, H.; Hussain, M. Allelopathic effects of rice on seedling development in wheat, oat, barley and berseem. Allelop. J. 2008, 22, 385–390. [Google Scholar]
- Kruidhof, H.; Bastiaans, L.; Kropff, M. Ecological weed management by cover cropping: Effects on weed growth in autumn and weed establishment in spring. Weed Res. 2008, 48, 492–502. [Google Scholar] [CrossRef]
- Stagnari, F.; Galieni, A.; Speca, S.; Cafiero, G.; Pisante, M. Effects of straw mulch on growth and yield of durum wheat during transition to conservation agriculture in Mediterranean environment. Field Crop. Res. 2014, 167, 51–63. [Google Scholar] [CrossRef]
- Khaliq, A.; Matloob, A.; Hussain, A.; Hussain, S.; Aslam, F.; Zamir, S.I.; Chattha, M.U. Wheat residue management options affect productivity, weed growth and soil properties in direct-seeded fine aromatic rice. Clean Soil Air Water 2015, 43, 1259–1265. [Google Scholar] [CrossRef]
- Zaji, B.; Majd, A. Allelopathic potential of canola (Brassica napus L.) residues on weed suppression and yield response of maize (Zea mays L.). In Proceedings of the International Conference on Chemical, Ecology and Environmental Sciences (IICCEES), Pattaya, Thailand, 17–18 December 2011; pp. 457–460. [Google Scholar]
- Alam, M.K.; Islam, M.M.; Salahin, N.; Hasanuzzaman, M. Effect of tillage practices on soil properties and crop productivity in wheat mung bean rice cropping system under subtropical climatic conditions. Sci. World J. 2014, 1, 1–15. [Google Scholar]
- Adugna, A.; Abegaz, A. Effects of land use changes on the dynamics of selected soil properties in northeast Wellega, Ethiopia. Soil 2016, 2, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, A.; Lal, R.; Shrestha, R.K.; Farooq, M. Mulching affects soil properties and greenhouse gases emissions under long term no-till and plough till systems in Alfisol of central Ohio. Land Dev. Degrad. 2016, 34, 45–56. [Google Scholar]
- Kamara, A.; Akobundu, I.; Chikoye, D.; Jutzi, S. Selective control of weeds in an arable crop by mulches from some multipurpose trees in south western Nigeria. Agrofor. Syst. 2000, 50, 17–26. [Google Scholar] [CrossRef]
- Khaliq, A.; Hussain, S.; Matloob, A.; Tanveer, A.; Aslam, F. Swine cress (Cronopus didymus L. Sm.) residues inhibit rice emergence and early seedling growth. Phillips. Agric. Sci. 2014, 96, 419–425. [Google Scholar]


{kind=link}
| Soil Properties | 2014 | 2015 |
|---|---|---|
| Soil bulk density (g/cm3) | 1.48 | 1.45 |
| Total soil porosity (%) | 43.10 | 44.30 |
| Soil pH | 7.85 | 7.79 |
| Electrical conductivity (dS/m) | 1.11 | 1.19 |
| Total soil organic matter (%) | 0.53 | 0.61 |
| Available P2O5 (mg/kg) | 6.74 | 6.95 |
| Exchangeable K2O (mg/kg) | 123.00 | 131.00 |
| Total nitrogen (soil) (g/kg) | 0.24 | 0.29 |
| Bacteria (cfu/g×105) | 35.00 | 45.00 |
| Fungi (cfu/g×104) | 5.00 | 8.00 |
| Activity of alkaline phosphatase (μg NP/g soil/ha) | 135.00 | 143.00 |
| Activity of dehydrogenase (μg TPF/g soil/ha) | 21.00 | 25.00 |
| Treatment | 2014 | 2015 | Mean Treatment (T) | 2014 | 2015 | Mean Treatment (T) |
|---|---|---|---|---|---|---|
| Soil bulk density (g/cm3) | Total soil porosity (%) | |||||
| (a)Control | 1.48 | 1.47 | 1.47 A | 42.82 | 44.06 | 43.44 C |
| (b)BWE 10 L/ha | 1.47 | 1.47 | 1.47 A | 43.52 | 44.10 | 43.81 C |
| BWE 20 L/ha | 1.48 | 1.47 | 1.47 A | 43.83 | 44.12 | 43.97 C |
| (c)BR 4 t/ha | 1.45 | 1.34 | 1.41 B | 45.40 | 46.99 | 46.19 B |
| BR 6 t/ha | 1.42 | 1.28 | 1.34 C | 45.96 | 48.51 | 47.23 A |
| Mean Year (Y) | 1.46 A | 1.40 B | 44.30 B | 45.56 A | ||
| LSD (p ≤ 0.05) | T = 0.06; Y = 0.04 | T = 1.02; Y = 0.77 | ||||
| Soil pH | Soil (d)EC (dS/m) | |||||
| Control | 7.77 | 7.74 | 7.76 A | 1.07 | 1.10 | 1.09 C |
| BWE 10 L/ha | 7.77 | 7.73 | 7.75 A | 1.09 | 1.13 | 1.11 C |
| BWE 20 L/ha | 7.77 | 7.73 | 7.75 A | 1.11 | 1.14 | 1.12 C |
| BR 4 t/ha | 7.49 | 7.46 | 7.48 B | 1.19 | 1.24 | 1.21 B |
| BR 6 t/ha | 7.46 | 7.26 | 7.36 C | 1.29 | 1.32 | 1.30 A |
| Mean (Y) | 7.65 A | 7.58 B | 1.15 B | 1.19 A | ||
| LSD (p ≤ 0.05) | T = 0.09; Y = 0.06 | T = 0.05; Y = 0.03 | ||||
| Total soil organic matter (%) | Total soil nitrogen (g/kg) | |||||
| Control | 0.68 d | 0.69 d | 0.68 C | 0.19 | 0.20 | 0.19 C |
| BWE 10 L/ha | 0.67 d | 0.69 d | 0.68 C | 0.20 | 0.20 | 0.20 C |
| BWE 20 L/ha | 0.68 d | 0.70 d | 0.69 C | 0.20 | 0.20 | 0.20 C |
| BR 4 t/ha | 0.89 c | 1.18 ab | 1.03 B | 0.28 | 0.33 | 0.30 B |
| BR 6 t/ha | 1.04 b | 1.26 a | 1.15 A | 0.30 | 0.38 | 0.34 A |
| Mean (Y) | 0.80 B | 0.91 A | 0.23 | 0.26 | ||
| LSD (p ≤ 0.05) | T = 0.11; Y = 0.07; T×Y = 0.15 | T = 0.03 | ||||
| Available potassium (mg/kg) | Available phosphorous (mg/kg) | |||||
| Control | 118.38 | 121.95 | 119.59 C | 6.72 d | 6.75 d | 6.73 C |
| BWE 10 L/ha | 119.55 | 120.66 | 120.66 C | 6.75 d | 6.76 d | 6.76 C |
| BWE 20 L/ha | 119.52 | 121.00 | 120.76 C | 6.77 d | 6.78 d | 6.77 C |
| BR 4 t/ha | 170.35 | 181.47 | 175.95 B | 7.93 c | 9.09 b | 8.51 B |
| BR 6 t/ha | 183.33 | 198.33 | 190.85 A | 9.09 b | 10.12 a | 9.60 A |
| Mean (Y) | 142.22 | 148.48 | 7.45 B | 7.89 A | ||
| LSD (p ≤ 0.05) | T = 12.44 | T = 0.39; Y = 0.25; T×Y = 0.56 | ||||
| Treatment | 2014 | 2015 | Mean Treatment (T) | 2014 | 2015 | Mean Treatment (T) |
|---|---|---|---|---|---|---|
| Bacteria (cfu/g × 105) 20 (d)DAS | Fungi (cfu/g × 104) 20 DAS | |||||
| (a)Control | 42 | 43 | 43 C | 7 d | 8 d | 7 C |
| (b)BWE 10 L/ha | 42 | 44 | 43 C | 7 d | 8 d | 8 C |
| BWE 20 L/ha | 44 | 45 | 45 C | 8 d | 9 cd | 8 C |
| (c)BR 4 t/ha | 55 | 65 | 60 B | 11 c | 15 b | 13 B |
| BR 6 t/ha | 67 | 78 | 71 A | 16 b | 20 a | 18 A |
| Mean Year (Y) | 50 B | 55 A | 10 B | 12 A | ||
| LSD (p ≤ 0.05) | T = 9.05; Y = 3.95 | T = 1.88; Y = 1.19; T × Y = 2.65 | ||||
| Bacteria (cfu/g × 105) (e)AH | Fungi (cfu/g × 104) AH | |||||
| Control | 19 | 20 | 20 C | 5 e | 6 de | 5 D |
| BWE 10 L/ha | 21 | 21 | 21 C | 6 de | 6 de | 6 CD |
| BWE 20 L/ha | 21 | 21 | 21 C | 6 de | 8 de | 7 C |
| BR 4 t/ha | 30 | 32 | 31 B | 8 d | 13 b | 10 B |
| BR 6 t/ha | 33 | 37 | 35 A | 10 c | 16 a | 13 A |
| Mean (Y) | 25 B | 27 A | 7 B | 10 A | ||
| LSD (p ≤ 0.05) | T = 3.21; Y = 1.65 | T = 1.47; Y = 0.93; T×Y = 2.08 | ||||
| Microbial activity (mg CO2-C kg−1 d−1) 20 DAS | Microbial activity (mg CO2-C kg−1 d−1) AH | |||||
| Control | 3.58 | 3.70 | 3.64 C | 2.97 | 3.12 | 3.04 C |
| BWE 10 L/ha | 3.61 | 3.75 | 3.68 C | 3.07 | 3.15 | 3.11 C |
| BWE 20 L/ha | 3.65 | 3.78 | 3.72 C | 3.09 | 3.18 | 3.14 C |
| BR 4 t/ha | 4.48 | 4.75 | 4.62 B | 3.69 | 3.85 | 3.77 B |
| BR 6 t/ha | 4.96 | 5.22 | 5.09 A | 4.08 | 4.28 | 4.18 A |
| Mean (Y) | 4.06 B | 4.24 A | 3.38 B | 3.51 A | ||
| LSD (p ≤ 0.05) | T = 0.45; Y = 0.15 | T = 0.31; Y = 0.11 | ||||
| Alkaline phosphatase (μg NP/g soil/h) | Dehydrogenase (μg TPF/g soil/h) | |||||
| Control | 134.77 | 134.80 | 134.78 C | 20.65 d | 22.35 d | 21.51 C |
| BWE 10 L/ha | 134.82 | 134.91 | 134.86 C | 22.16 d | 22.67 d | 22.42 C |
| BWE 20 L/ha | 135.12 | 135.18 | 135.15 C | 22.86 d | 23.25 d | 22.56 C |
| BR 4 t/ha | 159.22 | 163.35 | 161.28 B | 27.33 c | 33.35 b | 30.33 B |
| BR 6 t/ha | 175.48 | 186.44 | 180.96 A | 33.67 b | 38.33 a | 35.00 A |
| Mean (Y) | 147.88 B | 150.94 A | 24.74 B | 27.99 A | ||
| LSD (p ≤ 0.05) | T = 6.51; Y = 2.99 | T = 2.61; Y = 1.65; T×Y = 3.69 | ||||
| Treatment | 2014 | 2015 | Mean Treatment (T) | 2014 | 2015 | Mean (T) |
|---|---|---|---|---|---|---|
| Trianthema portulacastrum density (m2) | T. portulacastrum dry weight (g/m2) | |||||
| (a)Control | 164 | 164 | 164 A | 196 | 192 | 194 A |
| (b)BWE 10 L/ha | 160 | 156 | 158 A | 184 | 184 | 184 A |
| BWE 20 L/ha | 144 | 140 | 142 B | 164 | 164 | 164 B |
| (c)BR 4 t/ha | 108 | 80 | 94 C | 124 | 96 | 110 C |
| BR 6 t/ha | 80 | 64 | 72 D | 88 | 64 | 76 D |
| Mean Year (Y) | 131 A | 120 B | 38 A | 35 B | ||
| LSD (p ≤ 0.05) | T= 14.96; Y= 9.48 | T= 19.44; Y= 9.40 | ||||
| Cyperus rotundus density (m2) | C. rotundus dry weight (g/m2) | |||||
| Control | 40 | 40 | 40 A | 12 | 12 | 12 |
| BWE 10 L/ha | 40 | 40 | 40 A | 8 | 8 | 8 |
| BWE 20 L/ha | 32 | 32 | 32 B | 8 | 8 | 8 |
| BR 4 t/ha | 24 | 28 | 26 C | 4 | 4 | 4 |
| BR 6 t/ha | 20 | 24 | 22 D | 4 | 4 | 4 |
| Mean (Y) | 31 | 33 | 7 | 7 | ||
| LSD (p ≤ 0.05) | T= 3.36 | NS | ||||
| Total weeds density (m2) | Total weeds dry weight (g per 0.25 m2) | |||||
| Control | 229.25 a | 231.20 a | 230.23 A | 226.20 | 223.32 | 224.76 A |
| BWE 10 L/ha | 216.76 a | 215.72 a | 216.24 A | 211.00 | 209.40 | 210.20 B |
| BWE 20 L/ha | 183.20 b | 181.84 b | 182.52 B | 191.08 | 188.36 | 189.72 C |
| BR 4 t/ha | 161.20 b | 123.84 c | 142.52 C | 143.84 | 116.16 | 130.00 D |
| BR 6 t/ha | 121.40 c | 101.68 c | 111.54 D | 110.76 | 85.92 | 98.34 E |
| Mean (Y) | 182.36 A | 170.86 B | 176.58 A | 164.63 B | ||
| LSD (p ≤ 0.05) | T= 17.80; Y= 10.24; T×Y= 25.16 | T= 13.40; Y= 11.72 | ||||
| Treatment | 2014 | 2015 | Mean Treatment (T) | 2014 | 2015 | Mean Treatment (T) |
|---|---|---|---|---|---|---|
| Number of pods per plant | Number of seed per pod | |||||
| (a)Control | 14.03 | 15.17 | 14.60 E | 5.29 | 6.17 | 5.73 D |
| (b)BWE 10 L/ha | 16.92 | 18.18 | 17.55 D | 6.35 | 7.03 | 6.69 C |
| BWE 20 L/ha | 17.67 | 21.27 | 19.47 C | 6.55 | 7.18 | 6.87 BC |
| (c)BR 4 t/ha | 19.29 | 22.74 | 21.01 B | 7.04 | 7.68 | 7.36 B |
| BR 6 t/ha | 21.25 | 23.03 | 22.14 A | 9.14 | 9.56 | 9.35 A |
| Mean Year (Y) | 17.83 B | 20.08 A | 6.87 B | 7.53 A | ||
| LSD (p ≤ 0.05) | T= 1.08; Y= 0.68 | T= 0.51; Y= 0.32 | ||||
| Weight of 1000 seeds (g) | Yield (kg/ha) | |||||
| Control | 50.14 | 50.26 | 50.20 D | 743.2 f | 773.9 e | 758.5 E |
| BWE 10 L/ha | 52.28 | 53.19 | 52.74 C | 785.1 de | 795.9 d | 790.5 D |
| BWE 20 L/ha | 53.06 | 53.26 | 53.16 B | 842.0 c | 852.7 c | 847.4 C |
| BR 4 t/ha | 53.66 | 53.70 | 53.68 B | 923.6 b | 924.9 b | 924.3 B |
| BR 6 t/ha | 54.29 | 55.38 | 54.84 A | 1005.0 a | 1009.7 a | 1007.3 A |
| Mean (Y) | 52.69 | 53.16 | 859.78 B | 871.41 A | ||
| LSD (p ≤ 0.05) | T= 0.53 | T= 8.53; Y= 5.40; T×Y= 12.07 | ||||
| Treatments | Yield (kg/ha) | Adjusted Yield (kg/ha) | Gross Income (e)$/ha | Total Cost $/ha | Net Benefits $/ha | Benefit Cost Ratio |
|---|---|---|---|---|---|---|
| (a)Control | 759 | 683 | 765 | 615 | 150 | 0.24 |
| (b)BWE 10 L/ha | 791 | 712 | 797 | 624 | 173 | 0.28 |
| BWE 20 L/ha | 847 | 762 | 854 | 625 | 228 | 0.36 |
| (c)BR 4 t/ha | 924 | 832 | 931 | 653 | 278 | 0.43 |
| BR 6 t/ha | 1007 | 906 | 1015 | 668 | 347 | 0.52 |
| Remarks | $ 44.67/40 kg | 10% less than actual | 1$ = 98.5 (d)PKR |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, R.; Aslam, Z.; Maitah, M.; Zaman, Q.u.; Bashir, S.; Hassan, W.; Chen, Z. Sustainable Weed Control and Enhancing Nutrient Use Efficiency in Crops through Brassica (Brassica compestris L.) Allelopathy. Sustainability 2020, 12, 5763. https://0-doi-org.brum.beds.ac.uk/10.3390/su12145763
Ullah R, Aslam Z, Maitah M, Zaman Qu, Bashir S, Hassan W, Chen Z. Sustainable Weed Control and Enhancing Nutrient Use Efficiency in Crops through Brassica (Brassica compestris L.) Allelopathy. Sustainability. 2020; 12(14):5763. https://0-doi-org.brum.beds.ac.uk/10.3390/su12145763
Chicago/Turabian StyleUllah, Raza, Zubair Aslam, Mansoor Maitah, Qamar uz Zaman, Safdar Bashir, Waseem Hassan, and Zhongbing Chen. 2020. "Sustainable Weed Control and Enhancing Nutrient Use Efficiency in Crops through Brassica (Brassica compestris L.) Allelopathy" Sustainability 12, no. 14: 5763. https://0-doi-org.brum.beds.ac.uk/10.3390/su12145763