The Role of Feral Goats in Maintaining Firebreaks by Using Attractants


1
Small Ruminant Research Group, Department of Animal and Food Sciences, Universitat Autònoma de Barcelona, CP 08193 Bellaterra, Barcelona, Spain
2
Department of Biology-LINCC, Universidad de las Islas Baleares, CP 07122 Palma, Mallorca, Spain
3
Forestry Engineer, Ministry of Environment and Territory, Balearic Islands Government, C/del Gremi de Corredors, 10, CP 07009 Palma, Mallorca, Spain
*
Author to whom correspondence should be addressed.
Sustainability 2020, 12(17), 7144; https://0-doi-org.brum.beds.ac.uk/10.3390/su12177144
Submission received: 30 July 2020
/
Revised: 27 August 2020
/
Accepted: 29 August 2020
/
Published: 1 September 2020
(This article belongs to the Special Issue Pastoral Goat Husbandry and Environment)
Abstract
:The threat of large forest fires is increasing, and the main causes are the depopulation of rural areas, along with the effects of climate change. To counter this threat in recent decades, there have been numerous proposals and actions aimed at promoting grazing in the forest as a tool for controlling biomass fuel. However, the continued disappearance of traditional herds makes this activity difficult. Rural depopulation has also meant that domestic species become feral, being habitual in the case of goats. Currently, little is known about the role that feral goats can play in the fight against forest fires. In this work, an analysis is made on the effect of feral goats on the control of the vegetation in firebreak areas. Furthermore, the effect of attractants, such as water, salt, or food, on goat behavior is also studied. The study was carried out on the island of Mallorca, where a population of feral goats occupies the mountain areas, and where it is common for them to graze on the network of firebreaks. The results showed that these areas in themselves exert an attractive effect with respect to the neighboring forest, and that the herbaceous biomass is reduced. This effect was enhanced with the implementation of water and salt points, although only in certain periods of the year. In general, it was possible to reduce the phytovolume of many species without affecting biodiversity in the short or medium term. Therefore, strategic management of feral animals, aimed at firebreak areas, could contribute not only to reducing the risk of fires and, consequently, to the mitigation of climate change, but also to attracting these animals to the forests, thus avoiding their dispersion to conflictive places such as roads, residences, agricultural fields, and gardens.

1. Introduction
In recent decades, Europe, North America and parts of Asia have experienced an increase in their forest area [1]. This increase corresponds to the abandonment of agricultural land in remote and poorly productive areas, coinciding with the increase in urban area [2]. The decrease in agricultural, livestock, and forestry activity leads to the loss of the mosaic structure of the landscape, reducing its heterogeneity [3]. One of the effects of this change in land use is colonization by woody species of abandoned land [4]. In Mediterranean forests, this transformation leads to increasingly dense undergrowth, with a greater accumulation of biomass that, in turn, increases the risk of forest fires [5]. The increase in fuel continuity at landscape scale has led to the generation of large fires in the last decade, as has occurred in the Mediterranean area [6], including the Balearic Islands [7]. Furthermore, the existence of an understory composed of highly flammable sclerophyllous species, or large tufts of grasses, aids the spread of fire from the understory to the canopy, thus increasing its devastating effect [8].
On the other hand, global change also includes the effect of climate change, for which forecasts in the Mediterranean basin indicate an increase in periods of drought, torrential rain, and lightning storms [9], which are great hazards in the generation of forest fires. Higher risk is expected in areas where the forest cover is already very high, for example, in the Alps, the Pyrenees, or the Balkans [10,11]. The increase in man-made fires, either intentionally or accidentally [12], must also be accounted for.
One action in the prevention of forest fires is the opening of firebreak areas, which, in practice, involves clearing or cutting forest strips to contain the spread of fire [4]. The maintenance of these strips can be carried out by grazing, thus reducing the costs of mechanical clearing [13,14]. This practice constitutes a good example of the multifunctionality of the livestock sector and allows the local population to be involved in fire prevention [15]. Despite this, the availability of domestic livestock for the maintenance of these areas is limited due to the increasing abandonment of extensive livestock farming [16,17].
Parallel to the decrease in the livestock herd, an increase has been observed in the populations of feral herbivores and unmanaged feral livestock, with the highest densities for many decades being recorded [13]. These populations have also started to be perceived as possible tools in the prevention of forest fires, while guaranteeing the conservation of biodiversity [18]. Along these same lines, some authors have shown that it is possible to influence the behavior of wild herbivores through the use of attractants, such as water, food, or salt [13,19,20], thus causing changes in the structure and composition of the vegetation. However, the use of attractants for wild animals, even for a management purpose, could have an impact on their behavior and population dynamics [20], and if this were the case, it could be perceived by conservation sectors as an interference in ecological processes.
Feral livestock could be a tool for clime mitigation by the prevention of forest fires since, as they are not wild populations, their management is more justified. Furthermore, these animals, especially feral goats (Capra hircus L.), often cause damage to crops and gardens, forcing administrations to devote a lot of effort to their control or eradication [21,22]. So far, there are no known works that analyze the role that feral goats can play in the maintenance of firebreaks. The island of Mallorca, in the western Mediterranean, enables us to address this issue, since it has a network of firebreak areas and a population of feral goats, with serious management problems [22,23].
For this reason, the objective of this work is to analyze the effect of feral goats on the vegetation of the firebreak areas in pine forests. An analysis will also be performed on the use of attractants (water, food, and salt) to increase their presence in these areas and thus increase their impact on vegetation. The initial hypothesis is that the firebreak areas exert an attractive effect on feral goats, and this effect can be accentuated with the use of other attractants, thus helping to control plant biomass, and therefore the functionality of the firebreaks, for a longer time.
2. Materials and Methods
2.1. Study Area
The work was carried out in the Sierra de Na Burguesa (39°35′11″ N 2°33′55″ E) located in the municipality of Calvià in the southwestern part of the island of Mallorca (Balearic Islands, Spain). The altitude of the study area ranges between 400 and 503 m above sea level (asl). The climate is dry supra-Mediterranean, with a mean temperature of 17 °C, with January being the coldest month and August being the warmest. Annual rainfall is irregular, distributed mainly in spring and autumn, with annual lows of 313 mm and highs of 863 mm. The mean annual temperature was 18 °C for the two years of study, and the annual rainfall was 528 and 326 mm for the first and second year, respectively [24].
The vegetation is characterized by the dominance of Aleppo pine (Pinus halepensis M.) forests with an understory of Mediterranean scrubs, where species such as Erica multiflora L., Pistacia lentiscus L., Phillyrea angustifolia L., Cistus albidus L., Cistus monspeliensis L., and Anthyllis cytisoides L. are common. Perennial grasses predominate in the herbaceous layer, including Ampelodesmos mauritanica (Poiret) and Brachypodium retusum (Pers.), as well as other annual species.
The study area is part of the network of fire prevention firebreak areas on the island of Mallorca. The study area was located in a firebreak strip of variable width ranging between 30 and 50 m and a length of 2 km. Maintenance is the responsibility of the Council of Agriculture, Environment, and Territory of the Balearic Islands. It is carried out by means of selective mechanical or manual brush clearing, in addition to thinning and the occasional pruning of trees, with a frequency of 5–6 years, stipulated in the IV General Plan of defense against forest fires of the Balearic Islands [25], with the last clearing being carried out in early spring 2017. According to reports from the Ministry of Agriculture, Fisheries and Food of Spain, the last forest fire in the study area was in 1993, with a total surface of 492 hectares affected.
The area is frequented by feral goats (Capra hircus), weighing between 30–50 kg, generally forming small groups that do not exceed 50 animals, with one or more males. Their behavior is territorial, with foraging areas of between 45 and 170 hectares [26]. These goat populations probably come from the gradual abandoning of livestock from old traditional extensive farms, and as they currently lack management, they graze all over the Na Burguesa mountains.
2.2. Data Collection
To determine the effect of goats on plant species in the firebreak areas, 12 plots of 5 × 5 m were marked out. Plots were distributed in 6 pairs (one fenced plot and its respective unfenced plot). An attempt was made for each pair of plots to have the same ecological conditions (same inclination, orientation, and height), and they were less than 5 m apart. Fencing of the plots was performed immediately after the clearing maintenance of the firebreak areas, using a galvanized metal mesh 25 mm thick and 1.80 m high, thus guaranteeing the complete exclusion of goats. Each pair of plots was randomly distributed in the firebreak areas. The phytovolume of the bushes (including A. mauritanica) and the biodiversity of the vegetation in these plots were estimated before the clearing and at 2 years after clearing. In order to calculate the phytovolume (V) of each individual, the height (h), largest diameter (D), and smallest diameter (d) were measured, and then the inverted cone formula was used:
V = π·D·d·h/3
To determine the biodiversity, a variant of the point interception method was used [27], which consisted of tracing 2 linear transects, following the two diagonals of the plot. Every 10 cm, a 1 m rod was driven into the ground and all species that came into contact with it were noted.
The biomass of the herbaceous group was determined after two years of the clearing. The herbaceous biomass was obtained by collecting samples from the herbaceous stratum in 5 quadrants of 50 × 50 cm distributed randomly in each plot at the end of the study period. The samples were dried to constant weight at 103 °C.
The effect of attractants on the habitat used by feral goats was studied from fecal group counts, assuming that the greater the use of a territory, the greater the amount of deposited feces [28,29]. A total of 32 strip transects of 20 × 1 m were established in two areas, 16 in each, separated by a distance of 490 m. In each zone, eight transects were distributed in parallel in the firebreak area and another eight in the adjacent pine forest. These transects were cleared of excrement at the start of the study, and the accumulated excrement was counted and removed periodically. Six fecal pellets together were established as a minimum fecal group [30], with a similar appearance in color, consistency, and humidity [31]. The sampling period was two consecutive years (from July 2017 to July 2019). During the first year, no type of attractant was used, and for the second year, two water points were installed in aluminum drinking troughs, 16 × 14.5 × 6 cm with outlet regulators, connected to a 210-L water container and attached to a stake 60 cm above the ground. The containers were filled every 15 days on average, guaranteeing the availability of water. Also, from the second year, a 10 kg block of salt was placed about 3 m from each trough, at a height of approximately 80 cm. In each water replacement, 1 kg of corn grain was spread around each water container. The 16 transects closest to the attractants were placed at an average distance of 70 m, and the 16 farthest were placed 490 m away.
2.3. Statistical Data Processing
To evaluate habitat use, the accumulation of fecal groups in transects was analyzed as an indirect measurement of the number of animals present. Two study periods were established to analyze the effect of the attractants: “before”, referring to the first year prior to the use of attractants, and “after”, referring to the second year after placement. The total fecal group count was divided by the total days of accumulation and multiplied by 30 to obtain a mean value of fecal groups per month. The mean monthly accumulation of fecal groups was analyzed by means of a generalized linear mixed model (GLMM), using the transect as a random factor and using the following as fixed factors: the period, referring to before and after the placement of attractants; the season, referring to the four seasons of the year; the area, split between near and far with respect to the distance towards the attractants; and the habitat, differentiating between the firebreaks area and the forest.
The effect of the exclusion of goats on the accumulation of dry biomass of grasses was analyzed by means of a mixed general model with the treatment (inside referring to the fenced plot, and outside referring to the unfenced plot), and the area (near and far with respect to the distance from the attractant) as fixed factors and plot as a random factor.
As expected, the height, cover, and phytovolume of the vegetation were highly correlated (Spearman ρ > 0.77 p < 0.0001), so only the phytovolume was analyzed. This was normalized by a logarithmic transformation. To analyze the effect of the exclusion of goats on the phytovolume of woody species (including A. mauritanica), a general mixed model was performed, with plot and species as random factors and treatment, time (before clearing and two years after clearing), and zone as fixed factors, including interactions when they were significant. For the analysis, data from species that did not have a minimum of 10 individuals were excluded.
Similar analyses were carried out separately for the species groups according to the flammability index, since this index evaluates the ease of igniting and producing a flame when subjected to constant heat energy [32]. Table 1 shows the flammability index of the main species in the study area. Statistical analyses were adjusted with mixed models, with the plot as a random factor and treatment, period, and area as fixed factors. In some cases, it was only possible to test some of the interactions since there were not enough replicates at the different levels of each factor. For the most abundant species, this same model was re-adjusted considering only the individuals of the species.
All the aforementioned mixed models were adjusted using the lmer function of the lme4 package [35] by R, R Core Team [36]. The best model was selected following the Akaike information criterion (AIC) (models with lower AIC value were selected), and the residual distribution graphs were reviewed to ensure the adequacy of the models [37].
For the analysis of the effect of exclusion on diversity, the Shannon diversity index was calculated for each of the plots. The effect of the treatment and area as fixed factors was analyzed using a two-way ANOVA with the avo function by R, R Core Team [36].
3. Results
3.1. Effect of Habitat and Attractants on the Accumulation of Fecal Groups
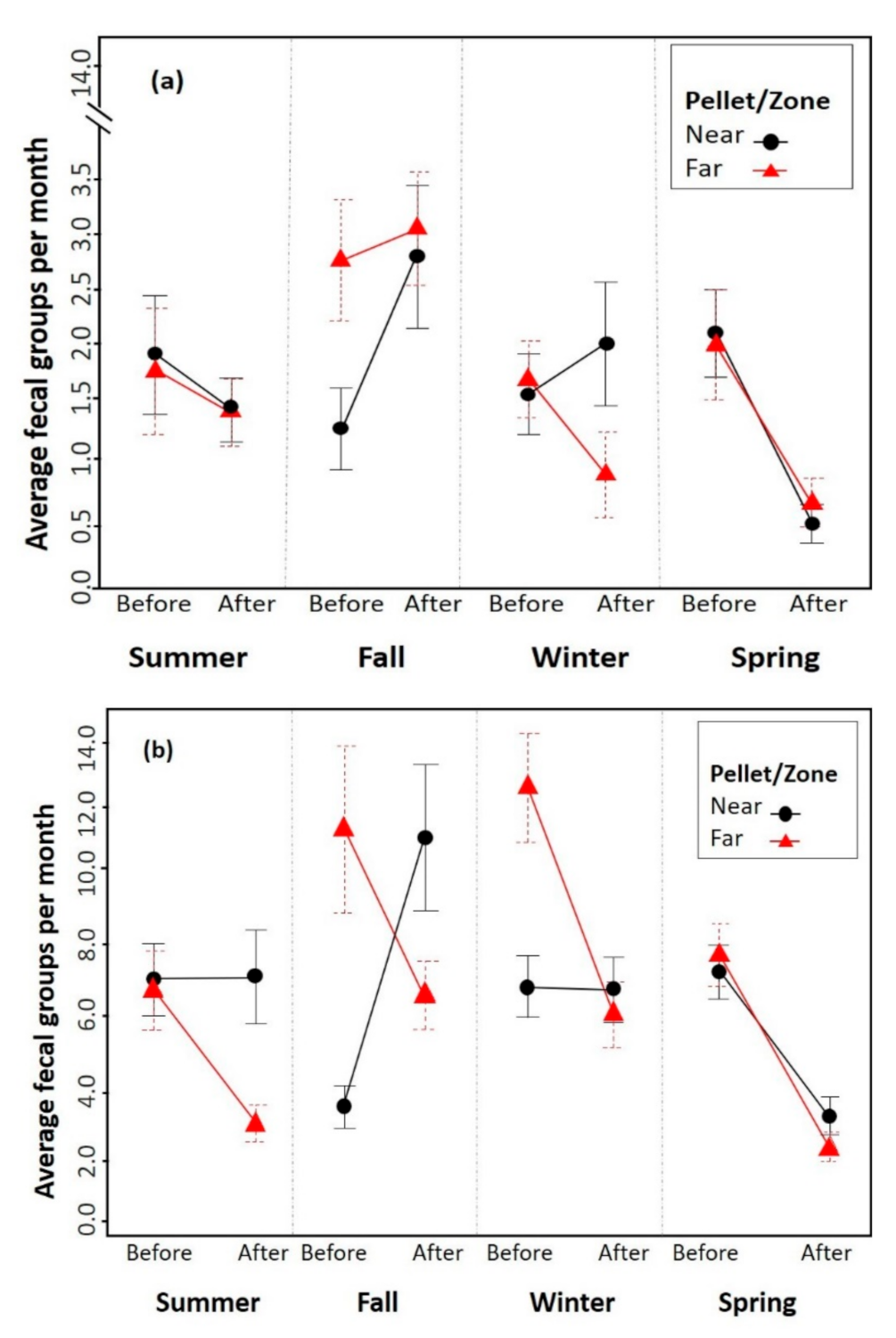
Statistically significant differences were found in the accumulation of fecal groups according to the period (p = 0.0005 GLMM), the season (p = 0.0002 GLMM), and the habitat (p < 0.0001 GLMM). The interactions between the period and habitat (p = 0.002 GLMM) and period and zone (p < 0.0001 GLMM) were also significant. Figure 1 illustrates the habitat effect, showing the number of fecal groups in the firebreaks area being 70% higher in relation to the forest (note the difference in scale between Figure 1a,b). Before the attractants were placed, the presence of fecal groups in the area furthest from attractants versus those closest was the same in most seasons of the year, except in autumn in the forest, and autumn and winter in the firebreaks, where it was greater in the furthest (Figure 1). After the attractants were placed, there was a general decrease in the accumulation of fecal groups in the forest habitat, except in autumn and winter in the nearby area, which increased. On the other hand, within the firebreak areas, after the placement of the attractants, fecal groups always decreased in the distant zone, whereas in the near zone they only decreased in spring, remaining stable in summer and winter and increasing in autumn.
3.2. Effect of the Exclusion of Goats on the Evolution of the Vegetation in the Firebreaks
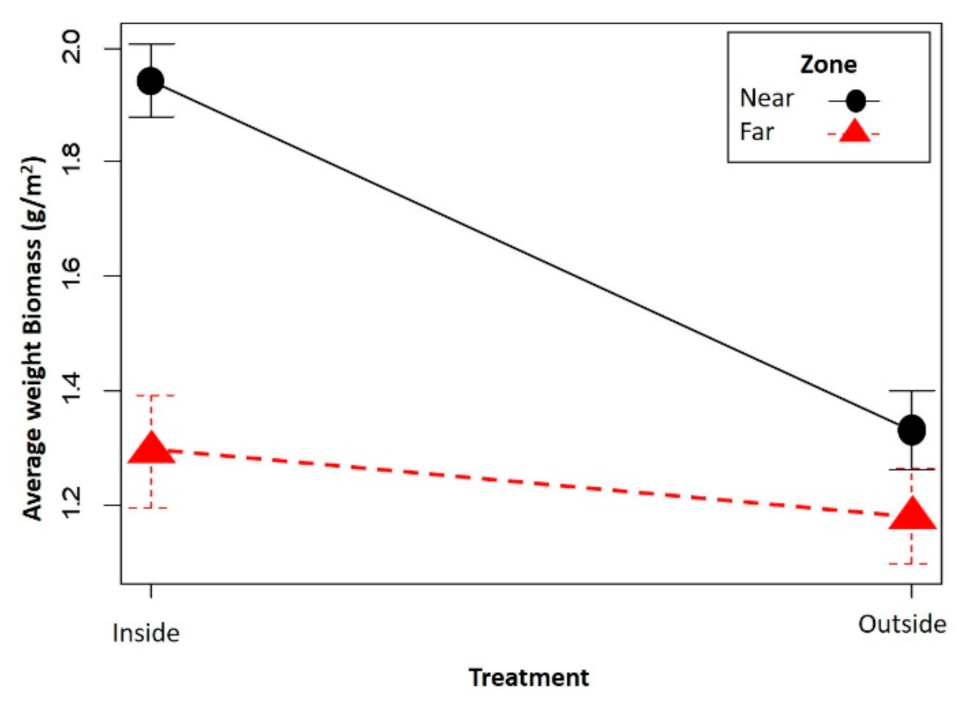
Significant differences were observed in the biomass of the herbaceous stratum, both between treatments (p = 0.02 GLMM) and between areas (p = 0.01 GLMM). The biomass of grasses in the fenced plot is 53% higher compared with the unfenced plot (Figure 2). This difference is more noticeable in plots near the attractant where it reaches 76%, while in the fenced plots far from the attractant, the biomass difference was only 37%.
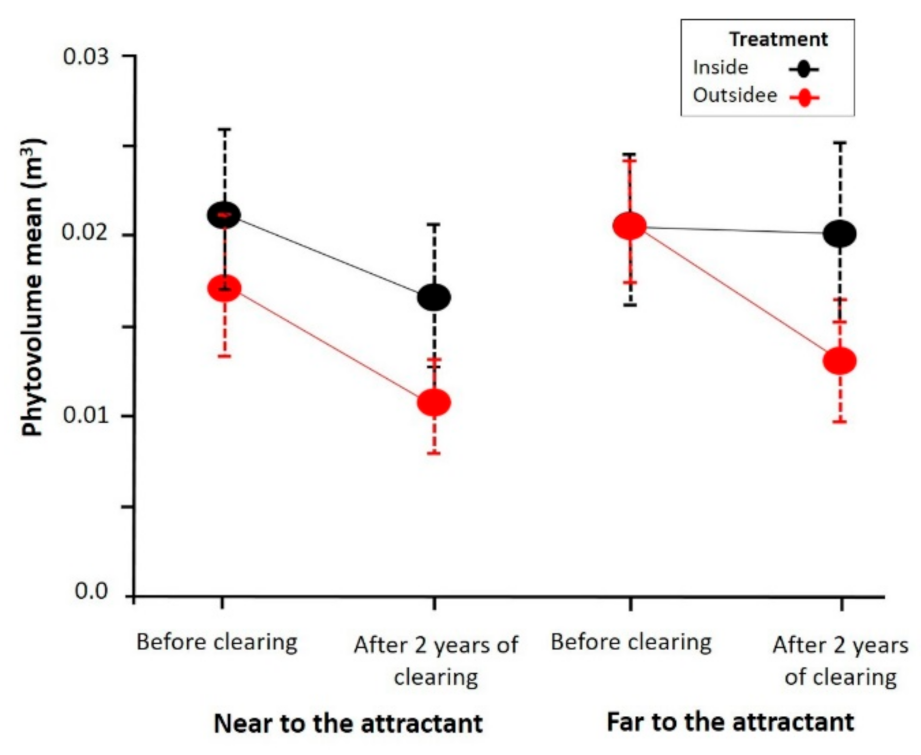
The results of the analysis of the variation of the phytovolume of the bushes and scrubs, including A. mauritanica and excluding species with less than 10 individuals in the study plots, are shown in Figure 3. There are significant differences regarding the treatment (p = 0.01 GLMM), with the phytovolume being greater within the fenced plots. There were also differences regarding time (p < 0.0001 GLMM), with greater phytovolume before the clearing. In contrast, no differences were found with respect to the area (p > 0.05 GLMM). A significant effect (p = 0.0002 GLMM) was observed in the interaction between the time and zone, showing that, after two years, the phytovolume within the plots far from the attractant was similar to the phytovolume before the clearing, whereas this effect was not noticeable in fenced plots near the attractant.
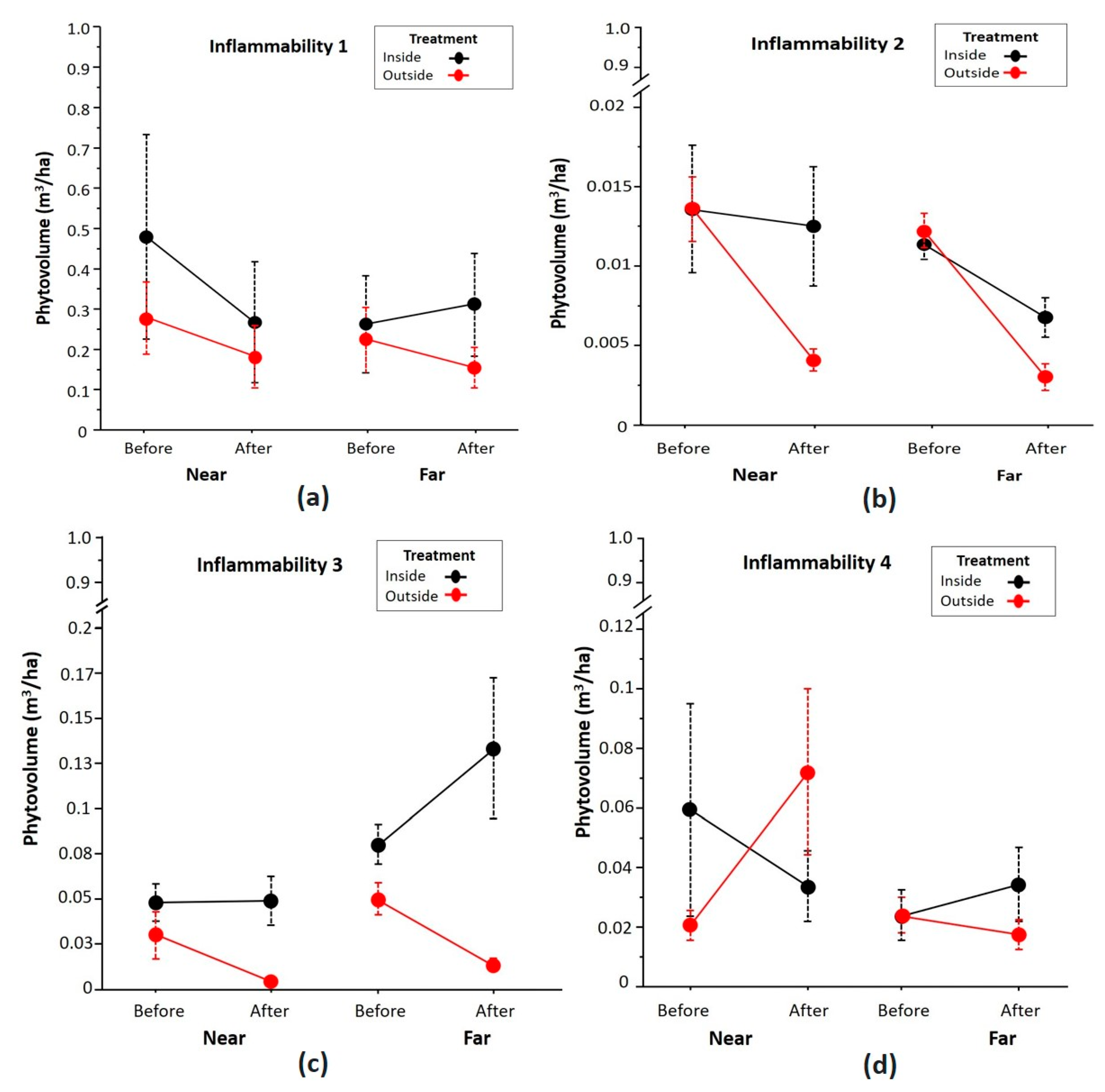
Figure 4 shows variations of the plant volume grouped according to the flammability index in the different bushes and shrubs, including A. mauritanica. For the species with flammability index 1, no significant differences (p > 0.05 GLMM) were detected for the treatment, area, and time factors. The species with flammability index 2 did not show significant differences with respect to treatment and area (p > 0.05 GLMM), while the time and treatment*time interaction showed significant differences (p = 0.001 GLMM) and (p = 0.002 GLMM), respectively, indicating that variation of the phytovolume is less within fenced plots after clearing. The species with flammability index 3 showed significant differences for the three factors: treatment (p = 0.005 GLMM), zone (p = 0.003 GLMM), and time (p = 0.02 GLMM), indicating differences in phytovolume inside and outside the fenced plots, as well as the effect of the distance to the attractant and differences after two years of clearing. In this case, the interaction between factors could not be calculated due to lack of replicates. For species with flammability index 4, no significant differences were found with any of the three factors (p > 0.05 GLMM). However, significant differences were observed for the treatment and time interaction (p < 0.05 GLMM). In this case, a significant increase in the phytovolume of these species was observed after two years, although only outside the fenced plots of the nearby area.
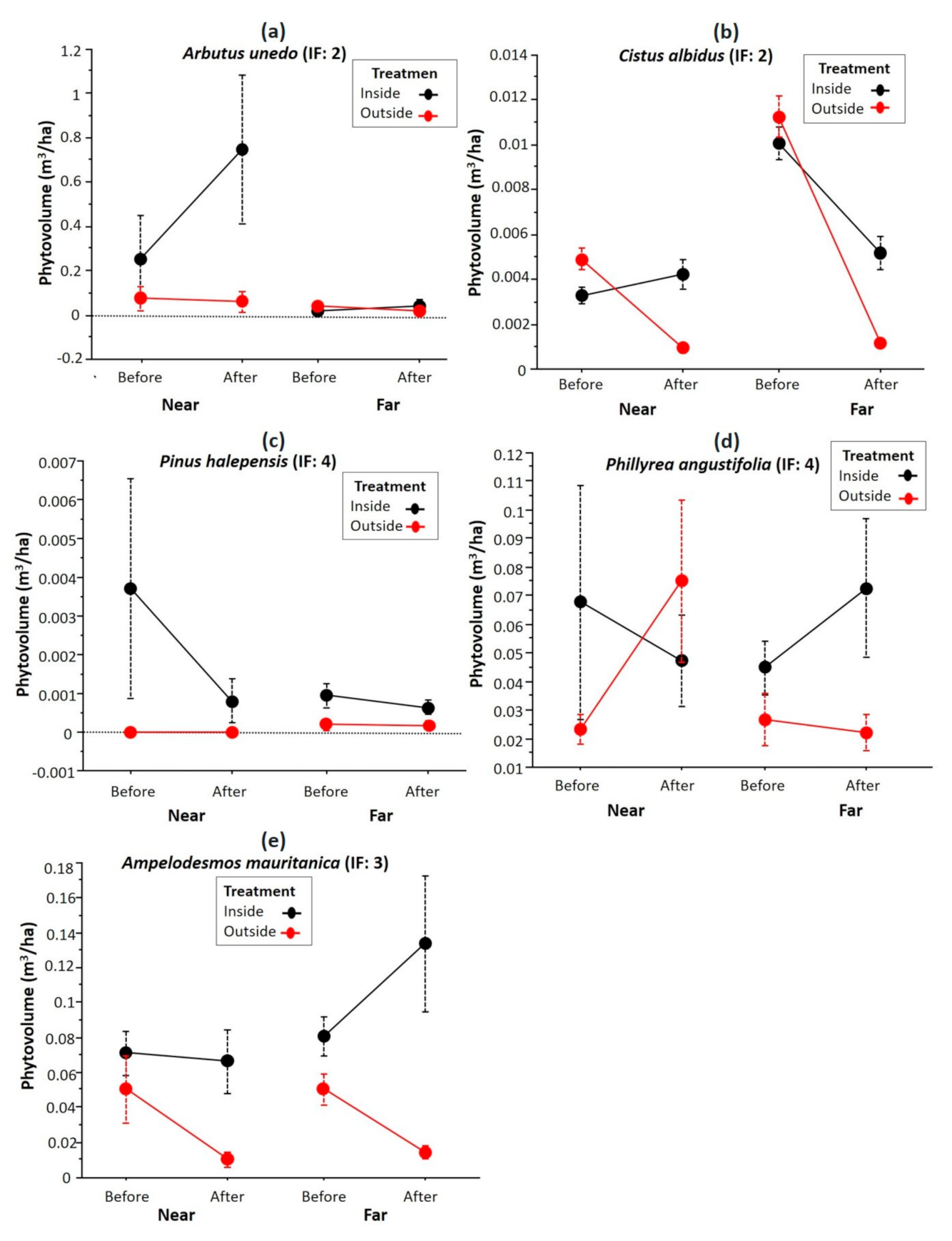
Figure 5 shows the plant volume of some of the most abundant species in the study area. In general, in the nearby plots a lower recovery of the phytovolume is observed in the unfenced plots (outside) compared with the fenced ones (inside). The phytovolume in some fenced plots, whether near or far from the attractant, showed a phytovolume higher than the initial record (two previous years), such as A. unedo (near), A. mauritanica (far), and P. angustifolia (far). Observing the phytovolume from outside the fenced plots, differences were observed both near and far from the attractant, except in P. angustifolia, with phytovolume around zero.
The Shannon biodiversity index (H) did not show significant differences between inside and outside the exclusions, nor between study areas (p > 0.05 ANOVA). H in the area near the attractants within the exclusions was 1.98 ± 0.04, while outside it was 1.85 ± 0.38. In the case of the exclusions of the area far from the attractants, within exclusion H was 1.99 ± 0.16 and outside was 1.91 ± 0.05.
4. Discussion
The present study shows the important role that feral goats play in the maintenance of firebreak areas and how the use of attractants increases these effects. The results show that feral goats use the firebreak area more than the adjacent forest, putting significant pressure on its vegetation. This is in agreement with other results in which it was observed that firebreak areas have herbaceous covers of higher quality and abundance for feral ungulates and extensive livestock, compared with woody formations [18,34]. For many ungulates, modified linear areas such as firebreaks and roads act as attractants, either because of their current biodiversity, the quantity or quality of existing food, or because they are corridors for movement [13,18,38]. In the case of goats, it should be added that, being prey animals, they usually prefer open spaces where they have a wide visual field to monitor possible predators [39].
Other authors also conclude that it is possible to reduce phytovolume and biomass by different species of domestic livestock, such as cows, sheep, and goats, and highlight the potentiating effect of the attractants [40,41]. The results obtained also show the effect on the distribution of feral goats in the autumn and winter seasons, with a greater effect on vegetation near the attractants.
It is worth noting the scarce difference detected in the number of fecal groups in the summer before the use of attractants and after their placement, when water shortage could have intensified its effect. This could be due to hunting carried out 2 km from the firebreak between the months of April and June, when 257 feral goats were killed, as well as the hunting in September, when another 67 goats were killed. This would explain the general decrease in excrement accumulation in the second year of study compared with the first. Across the island, feral goat population management systems are highly heterogeneous, from administration controlled areas to completely abandoned areas, including hunting areas [18]. This makes population management very difficult since there are significant fluctuations in populations at a local level. Public administration will have to make a coordinated effort to harness the potential of these herds in maintaining firewall areas [21].
The behavior of feral goats, in regard to the range areas, was modified after placing attractants during the summer, winter, and autumn seasons, as the use of the area increased compared with before their placement. This phenomenon has also been observed in elk management, where lumps of salt were distributed at strategic points in their territories to avoid collisions with vehicles on the roads [42,43]. These results are of special relevance for the management of the territory, since extensive livestock farming is in decline, and the populations of feral animals and the forest biomass are increasing [18].
In this work, the differences in the biomass of herbaceous plants within the fenced plots compared with the biomass of those outside them were particularly notable, indicating that grazing decreases their biomass, especially in areas near water and salt supply points. Similar results have been obtained with equine cattle in Galicia (Spain), where it was evident that the continuous and rotational management of the grazing system modifies the productivity of herbaceous plants and the evolution of understory biodiversity [44]. This has also been observed with sheep in Andalusia (Spain), where a decrease in biomass of between 53% and 72% was observed [45]. In the case of domestic goats in mountain areas where grazed and ungrazed areas were compared, a decrease in understory biomass was also observed [46].
It should be noted that the herbaceous layer of the study area is dominated by Brachypodium retusum, which is a common grass in the Mediterranean basin with flammable characteristics and early post-fire regeneration [47]. On the other hand, this grass constitutes the fine fuel for the initiation of many forest fires [48], so its reduction must be considered a success in the management of feral goats. From the results obtained, it also follows that the use of supplementary attractants, such as water and salt, increase the clearing effect of animals within a discrete radius of the supply point, in the order of tens of meters in this case, a distance similar to that obtained in other studies [13,49].
Key data in firebreaks management refer to the time it takes for shrub vegetation to recover after mechanical cutting. These data will determine the frequency of clearing and, therefore, the efficiency of prevention tasks. In this work, it has been observed that part of the parcels in exclusion recover the phytovolume that existed before clearing in two years, while others require more time. It is also observed that the non-fenced plots (with free grazing), adjacent to those that recover the phytovolume in two years, have not recovered it in this period. This indicates that feral goats could delay the frequency of clearing, thereby saving management resources for fuel treatment. This has already been observed using domestic livestock, especially goats [50,51,52]. However, in the case of feral goats, this effect does not occur in all the plots, possibly due to lack of management that forces them to remain in the area. In the case of this study, reduction in plant volume depends on the area and the shrub species.
The results of the analysis of phytovolume variations as a function of flammability showed that, in general, outside the exclusion plots there is a reduction in phytovolume for all classes of flammability, although in the group of non-flammable species (1) this reduction is not significant. However, in the case of the highly flammable group of species (4), it was observed that in plots near the supplementary attractants the phenomenon was reversed, with a significant increase in the phytovolume of this group of species outside the fences. Aleppo pine (P. halepensis), a dominant tree in the area, is found in this group and is not very appealing to animals due to its high content of secondary compounds [53]. In this particular case, the browsing effect is likely to reduce competition with other species and favor the growth of highly flammable and poorly palatable species, such as pine. Something similar could happen with P. angustifolia, which is also highly flammable but much more palatable.
When the variability of the phytovolume by species in the grazed plots is analyzed after the two years of treatment, a lower recovery is observed in most of them. The effectiveness of feral goats in reducing fuel in firebreaks is thus verified. In addition, the use of feral goats also offers the potential to reduce fuel used in mechanical brush clearing, with a likely reduction in greenhouse gas emission. However, as previously discussed for pine, in some species the effect seems to be the opposite. This would be the case for P. angustifolia, which increased the phytovolume in the ungrazed plots near the attractant supply points. When browsing, animals are likely to reduce the competition of other species, thus favoring their growth.
Regarding biodiversity, grazing treatment and supply of attractants had no effect in the short–medium term. After two years of clearing and one year with attractants, no differences were observed in the Shannon indices. The values were low, below 2, probably due to low floristic richness and the dominance of some species. Two years is likely to be too short a period for changes in vegetation biodiversity to occur, especially in perennial species [13].
5. Conclusions
From the results obtained, it can be concluded that firebreak areas are places of attraction for feral goats, and their browsing activity contributes to slowing down the recovery of vegetation after mechanical clearing. This effect can be enhanced by supplying attractants, such as water and salt, at certain times of year and is especially evident in the reduction of the biomass of fine fuel, mainly Brachypodium retusum. As for the control of woody species, the effect depends on each area and each species, although, in general, it is possible to reduce the phytovolume of many species without affecting biodiversity in the short or medium term. The effect is probably less forceful than that achieved by directed grazing with domesticated herds. In this sense, support and recovery policies for rural pastoral communities would be necessary. However, this livestock has been decreasing since giving territory to populations of feral animals is a management strategy to attract feral goat populations to firebreak areas. This would contribute not only to reducing the risk of fires and, consequently, to the mitigation of climate change but also to the distribution of these animals to less conflictive places that do not cause damage in particularly fragile areas.
Author Contributions
Conceptualization, E.B. and J.B.; Formal analysis, J.P. and E.B.; Funding acquisition, E.B. and J.B.; Investigation, J.P., E.B., M.I., O.D. and J.B.; Methodology, J.P., E.B. and J.B.; Project administration, J.B.; Resources, E.B. and J.B.; Supervision, J.B.; Visualization, J.P.; Writing—original draft, J.P., E.B. and J.B.; Writing—review & editing, J.B., E.B., J.P., M.I. and O.D. All authors have read and agreed to the published version of the manuscript.
Funding
Funding was provided by Ministry of Economy and Competitiveness (MINECO) of Spain through the project CGL2015-70449-R. Javier Pareja was supported by the National Fund for Scientific, Technological Development and Technological Innovation (FONDECYT), the funding branch of the National Council for Science, Technology and Technological Innovation (CONCYTEC) Peru (Grant contract number N° 236-2015-FONDECYT).
Acknowledgments
Thanks to David Alomar, Miquel Ribas, Gabriella Rizzardini, Txell Grau, Elizabeth Siddall Araceli Gort and David Bartolomé for their help in data field collection. To the owners of the area of study to facilitate us the work in their propriety.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of Global Forest Area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- García-Ruiz, J.M.; Lana-Renault, N. Hydrological and Erosive Consequences of Farmland Abandonment in Europe, with Special Reference to the Mediterranean Region—A Review. Agric. Ecosyst. Environ. 2011, 140, 317–338. [Google Scholar] [CrossRef]
- Navarro, L.M.; Pereira, H.M. Rewilding Abandoned Landscapes in Europe. In Rewilding European Landscapes; Springer: Cham, Switzerland, 2015; pp. 3–23. [Google Scholar] [CrossRef] [Green Version]
- Loepfe, L.; Martinez-Vilalta, J.; Oliveres, J.; Piñol, J.; Lloret, F. Feedbacks between Fuel Reduction and Landscape Homogenisation Determine Fire Regimes in Three Mediterranean Areas. For. Ecol. Manag. 2010, 259, 2366–2374. [Google Scholar] [CrossRef]
- Lasanta-Martínez, T.; Vicente-Serrano, S.M.; Cuadrat-Prats, J.M. Mountain Mediterranean Landscape Evolution Caused by the Abandonment of Traditional Primary Activities: A Study of the Spanish Central Pyrenees. Appl. Geogr. 2005, 25, 47–65. [Google Scholar] [CrossRef]
- San-Miguel-Ayanz, J.; Moreno, J.M.; Camia, A. Analysis of Large Fires in European Mediterranean Landscapes: Lessons Learned and Perspectives. For. Ecol. Manag. 2013, 294, 11–22. [Google Scholar] [CrossRef]
- Domenech, O. Incendis Forestals a Les Illes Balears: 25 Anys de Dades Estadístiques per a La Defensa Integrada i La Conservació Del Patrimoni Natural. Llibre Verd Protecció d’Espècies Balear. Soc. d’Història Nat. Balear. 2015, 20, 487–493. [Google Scholar]
- Pausas, J.G.; Paula, S. Fuel Shapes the Fire-Climate Relationship: Evidence from Mediterranean Ecosystems. Glob. Ecol. Biogeogr. 2012, 21, 1074–1082. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate Change Impacts, Adaptive Capacity, and Vulnerability of European Forest Ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Fernández-Olalla, M.; Muñoz-Igualada, J.; Martínez-Jauregui, M.; Rodríguez-Vigal, C.; San Miguel-Ayanz, A. Selección de Especies y Efecto Del Ciervo (Cervus elaphus L.) Sobre Arbustedos y Matorrales de Los Montes de Toledo, España Central. Investig. Agrar. Sist. Recur. For. 2006, 15, 329–338. [Google Scholar] [CrossRef]
- Moreira, F.; Viedma, O.; Arianoutsou, M.; Curt, T.; Koutsias, N.; Rigolot, E.; Barbati, A.; Corona, P.; Vaz, P.; Xanthopoulos, G.; et al. Landscape—Wildfire Interactions in Southern Europe: Implications for Landscape Management. J. Environ. Manag. 2011, 92, 2389–2402. [Google Scholar] [CrossRef] [Green Version]
- Radeloff, V.C.; Helmers, D.P.; Anu Kramer, H.; Mockrin, M.H.; Alexandre, P.M.; Bar-Massada, A.; Butsic, V.; Hawbaker, T.J.; Martinuzzi, S.; Syphard, A.D.; et al. Rapid Growth of the US Wildland-Urban Interface Raises Wildfire Risk. Proc. Natl. Acad. Sci. USA 2018, 115, 3314–3319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velamazán, M.; San Miguel, A.; Escribano, R.; Perea, R. Use of Firebreaks and Artificial Supply Points by Wild Ungulates: Effects on Fuel Load and Woody Vegetation along a Distance Gradient. For. Ecol. Manag. 2018, 427, 114–123. [Google Scholar] [CrossRef]
- Lovreglio, R.; Meddour-Sahar, O.; Leone, V. Goat Grazing as a Wildfire Prevention Tool: A Basic Review. iForest Biogeosciences For. 2014, 7, 260. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Mirazo, J.; Robles, B.A.; Jiménez, R.; Martínez-Moya, J.L.; López-Quintanilla, J.; González-Rebollar, L. La Prevención de Incendios Forestales Mediante Pastoreo Controlado: El Estado del Arte en Andalucía. In Proceedings of the 4th International Wildland Fire Conference, Sevilla, Spain, 13–17 May 2007. [Google Scholar]
- Lasanta, T.; Nadal-Romero, E.; Arnáez, J. Managing Abandoned Farmland to Control the Impact of Re-Vegetation on the Environment. The State of the Art in Europe. Environ. Sci. Policy 2015, 52, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, K.; Bouchard, A.; Domon, G. Managing Abandoned Farmland: The Need to Link Biological and Sociological Aspects. Environ. Manag. 2008, 42, 603–619. [Google Scholar] [CrossRef]
- Ruiz-Mirazo, J.; Robles, A.B.; González-Rebollar, J.L. Two-Year Evaluation of Fuelbreaks Grazed by Livestock in the Wildfire Prevention Program in Andalusia (Spain). Agric. Ecosyst. Environ. 2011, 141, 13–22. [Google Scholar] [CrossRef]
- Bailey, D.W. Identification and Creation of Optimum Habitat Conditions for Livestock. Rangel. Ecol. Manag. 2005, 58, 109–118. [Google Scholar] [CrossRef]
- Sahlsten, J.; Bunnefeld, N.; Månsson, J.; Ericsson, G.; Bergström, R.; Dettki, H. Can Supplementary Feeding Be Used to Redistribute Moose Alces Alces? Wildlife Biol. 2010, 16, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Roque, M.À. Entre la Ecología Ambiental y el Turismo Cinegético. Cabras en la Sierra de Tramuntana. Quad. Mediterrània 2015, 25, 282–293. [Google Scholar]
- Vives, J.A.; Baraza, Y.E. La Cabra Doméstica Asilvestrada (Capra hircus) en Mallorca ¿Una Especie a Erradicar? Galemys 2010, 22, 193–205. [Google Scholar]
- Mayol, J.; Alcover, J.A.; Domenech, O.; Moragues, E.; Rita, J. La Cabra, Espècie Invasora a les Balears; Lleonard Muntaner Editor: Illes Balears, Spain, 2017; pp. 1–96. [Google Scholar]
- Agencia Estatal de Meteorología-AEMET. Gobierno de España. Available online: http://www.aemet.es/es/eltiempo/prediccion/municipios/calvia-id07011 (accessed on 8 May 2019).
- Gobierno de las Islas Baleares. IV Plan General de Defensa Contra Incendios Forestales de las Islas Baleares. Available online: http://www.caib.es/sites/xarxaforestal/es/iv_plan_general-66858/ (accessed on 8 May 2020).
- Ibáñez, M.; Gort, A.; Baraza, E.; Serrano, E.; Bartolome, J. Pattern of Territory Use by Feral Goats on Insular Mediterranean Habitats. In Proceedings of the II Movement Ecology Brazil Meeting, Campo Grande, MS, Brasil, 24 June–5 July 2019; p. 13. [Google Scholar]
- Elzinga, C.L.; Salzer, D.W.; Willoughby, J.W. Measuring & Monitering Plant Populations; US Department of the Interior, Bureau of Land Management: Denver, CO, USA, 1998; pp. 1–1730.
- Fattorini, L.; Ferretti, F.; Pisani, C.; Sforzi, A. Two-Stage Estimation of Ungulate Abundance in Mediterranean Areas Using Pellet Group Count. Environ. Ecol. Stat. 2011, 18, 291–314. [Google Scholar] [CrossRef]
- Härkönen, A.; Harkonen, S.; Heikkila, R. Use of Pellet Group Counts in Determining Density and Habitat Use of Moose Alces Alces in Finland. Wildlife Biol. 2018, 5, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, P.; Ferreres, J.; Jaroso, R.; Durán, M.; Escudero, M.A.; Marco, J.; Gortázar, C. Estimating Roe Deer Abundance from Pellet Group Counts in Spain: An Assessment of Methods Suitable for Mediterranean Woodlands. Ecol. Indic. 2010, 10, 1226–1230. [Google Scholar] [CrossRef]
- Sutherland, W.J. Ecological Census Techniques: A Handbook; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Valette, J.C. Inflammabilities of Mediterranean Species. In Forest Fire Risk and Management; Balabanis, P., Eftichidis, G., Fantechi, R., Eds.; Porto Carras: Chalkidiki, Greece, 1992; pp. 51–64. [Google Scholar] [CrossRef]
- Henaoui, S.E.-A.; Bouazza, M.; Hassiba, S.-M. Inflammabilité et Teneur en eau des Communautés Végétales à Cistus Dans la Région de Tlemcen (Nord-OuestAlgérien). Eur. Sci. J. 2013, 9, 1857–7881. [Google Scholar]
- Mancilla-Leytón, J.M.; Pino Mejías, R.; Martín Vicente, A. Do Goats Preserve the Forest? Evaluating the Effects of Grazing Goats on Combustible Mediterranean Scrub. Appl. Veg. Sci. 2013, 16, 63–73. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. lme4 (Version 1.1–8). Available online: http://cran.r-project.org/package=lme4 (accessed on 12 January 2019).
- R Development Core Tea. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Royle, A.; York, G.M.S.N. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Suárez-Esteban, A.; Delibes, M.; Fedriani, J.M. Barriers or Corridors? The Overlooked Role of Unpaved Roads in Endozoochorous Seed Dispersal. J. Appl. Ecol. 2013, 50, 767–774. [Google Scholar] [CrossRef] [Green Version]
- Wittmer, H.U. Mountain Goats: Ecology, Behavior, and Conservation of an Alpine Ungulate. Ecology 2008, 89, 2666–2667. [Google Scholar] [CrossRef]
- Ruiz-Mirazo, J.; Robles, A.B.; González-Rebollar, J.L. Efecto de la Colocación de Terrones de sal sobre el Ramoneo del Estrato Arbustivo en Áreas Pasto-Cortafuegos de Andalucía; SEEP: Zamora, Spain, 2010; pp. 487–492. [Google Scholar]
- Mancilla-Leytón, J.M.; Vicente, A.M. Effect of Agro-Industrial by-Products on Browsing of Rosmarinus Officinalis by Goats. J. Arid Environ. 2014, 110, 8–11. [Google Scholar] [CrossRef]
- Grosman, P.D.; Jaeger, J.A.G.; Biron, P.M.; Dussault, C.; Ouellet, J.-P. Trade-off between Road Avoidance and Attraction by Roadside Salt Pools in Moose: An Agent-Based Model to Assess Measures for Reducing Moose-Vehicle Collisions. Ecol. Modell. 2011, 222, 1423–1435. [Google Scholar] [CrossRef]
- Laurian, C.; Dussault, C.; Ouellet, J.-P.; Courtois, R.; Poulin, M.; Breton, L. Behavioral Adaptations of Moose to Roadside Salt Pools. J. Wildl. Manag. 2008, 72, 1094–1100. [Google Scholar] [CrossRef]
- Rigueiro-Rodríguez, A.; Mouhbi, R.; Santiago-Freijanes, J.J.; del Pilar González-Hernández, M.; Mosquera-Losada, M.R. Horse Grazing Systems: Understory Biomass and Plant Biodiversity of a Pinus radiata Stand. Sci. Agric. 2012, 69, 38–46. [Google Scholar] [CrossRef]
- Robles, A.B.; Ruiz, J.; González-Rebollar, J.L. Sheep Grazing in a Firebreak. Effects on the Herbaceous Fuel Load. In La Multifuncionalidad de los Pastos: Producción Ganadera Sostenible y Gestión de los Ecosistemas. XLVIII Reunión Científica de la Sociedad Española para el Estudio de los Pastos; Sociedad Española para el Estudio de los Pastos (SEEP): Huesca, Spain, 2009; pp. 657–662. [Google Scholar]
- Marques, D.; Fachada, M.; Viana, H. Synergies between Goat Grazing and Shrub Biomass in Mountain Areas. Sustain. Goat Prod. Advers. Environ. 2017, 1, 155–175. [Google Scholar] [CrossRef]
- Santana, V.M.; Baeza, M.J.; Marrs, R.H. Response of Woody and Herbaceous Fuel to Repeated Fires in Mediterranean Gorse Shrublands. Int. J. Wildl. Fire 2013, 22, 508–514. [Google Scholar] [CrossRef]
- Vilà-Cabrera, A.; Saura-Mas, S.; Lloret, F. Effects of Fire Frequency on Species Composition in a Mediterranean Shrubland. Écoscience 2008, 15, 519–528. [Google Scholar] [CrossRef]
- Månsson, J.; Roberge, J.-M.; Edenius, L.; Bergström, R.; Nilsson, L.; Lidberg, M.; Komstedt, K.; Ericsson, G. Food Plots as a Habitat Management Tool: Forage Production and Ungulate Browsing in Adjacent Forest. Wildlife Biol. 2015, 21, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Bowie, D.D.; Kumi, A.S.; Min, B.R.; Smith, R.C.; Davis, R.J.; Elliott, A.W.; Gurung, N.K. Preliminary Observations on Effects of Using Different Stocking Rates of Meat Goats to Control Understory Vegetation in Longleaf Pine Stands. Agrofor. Syst. 2016, 90, 747–761. [Google Scholar] [CrossRef]
- Jáuregui, B.M.; Celaya, R.; García, U.; Osoro, K. Vegetation Dynamics in Burnt Heather-Gorse Shrublands under Different Grazing Management with Sheep and Goats. Agrofor. Syst. 2007, 70, 103–111. [Google Scholar] [CrossRef]
- Torrano, L.; Valderrábano, J. Grazing Ability of European Black Pine Understorey Vegetation by Goats. Small Rumin. Res. 2005, 58, 253–263. [Google Scholar] [CrossRef]
- Burney, O.T.; Jacobs, D.F. Ungulate Herbivory of Boreal and Temperate Forest Regeneration in Relation to Seedling Mineral Nutrition and Secondary Metabolites. New For. 2013, 44, 753–768. [Google Scholar] [CrossRef]
Figure 1.
Number of fecal groups in the forest (a) and firebreaks (b) throughout the seasons, depending on the period of installation of attractants (Before or After) and the distance to them. Circles and triangles represent the mean, and vertical lines represent the standard error.
Figure 1.
Number of fecal groups in the forest (a) and firebreaks (b) throughout the seasons, depending on the period of installation of attractants (Before or After) and the distance to them. Circles and triangles represent the mean, and vertical lines represent the standard error.

Figure 2.
Variation of the weight of the herbaceous biomass in the firebreaks, depending on the fenced and unfenced plots (inside and outside) and the distance to the area of attractants (near and far). Circles and triangles represent the mean, and vertical lines represent their standard error.
Figure 2.
Variation of the weight of the herbaceous biomass in the firebreaks, depending on the fenced and unfenced plots (inside and outside) and the distance to the area of attractants (near and far). Circles and triangles represent the mean, and vertical lines represent their standard error.

Figure 3.
Variation of the phytovolume of the firebreaks bushes according to the treatment, location inside and outside of the plots, zone, location near and far with respect to the distance far from the attractants, and time before the clearing and after two years. The circle represents the mean, and vertical lines represent its standard error.
Figure 3.
Variation of the phytovolume of the firebreaks bushes according to the treatment, location inside and outside of the plots, zone, location near and far with respect to the distance far from the attractants, and time before the clearing and after two years. The circle represents the mean, and vertical lines represent its standard error.

Figure 4.
Variation of the phytovolume of the bushes grouped by the flammability index on (a) scale 1, (b) scale 2, (c) scale 3, and (d) scale 4 with respect to the treatment factors (inside and outside the fenced plots), time (before and two years after clearance), and area (near and far from the attractant). The circles represent the means and the vertical lines their standard error.
Figure 4.
Variation of the phytovolume of the bushes grouped by the flammability index on (a) scale 1, (b) scale 2, (c) scale 3, and (d) scale 4 with respect to the treatment factors (inside and outside the fenced plots), time (before and two years after clearance), and area (near and far from the attractant). The circles represent the means and the vertical lines their standard error.

Figure 5.
Variation of the phytovolume of the most abundant shrub species in the firebreaks (a) Arbutus unedu, (b) Cistus albidus, (c) Pinus halepensis, (d) Phyllirea angustifolia, and (e) Ampelodesmos mauritanica depending on the treatment factors (fenced plots: inside, and unfenced plots: outside), time (before clearing and two years after clearing), and zone (near and far regarding the attractant). The circle represents the mean, the vertical lines represent its standard error, and the IF represents the flammability index.
Figure 5.
Variation of the phytovolume of the most abundant shrub species in the firebreaks (a) Arbutus unedu, (b) Cistus albidus, (c) Pinus halepensis, (d) Phyllirea angustifolia, and (e) Ampelodesmos mauritanica depending on the treatment factors (fenced plots: inside, and unfenced plots: outside), time (before clearing and two years after clearing), and zone (near and far regarding the attractant). The circle represents the mean, the vertical lines represent its standard error, and the IF represents the flammability index.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Flammability index of the main species in the Sierra de Na Burguesa, Mallorca, Spain.
| Species | Stratum | Flammability Index 1 |
|---|---|---|
| Ampelodesmos mauritanica | Herbaceous | 3 |
| Anthyllis cytisoides | Shrubby | 2 |
| Arbutus unedo | Arboreous | 2 |
| Asparagus acutifolius | Shrubby | 3 |
| Ceratonia siliqua | Arboreous | 3 |
| Cistus albidus | Shrubby | 2 |
| Cistus monspeliensis | Shrubby | 2 |
| Erica multiflora | Shrubby | 2 |
| Euphorbia characias | Shrubby | 3 |
| Globularia alypum | Shrubby | 3 |
| Lonicera sp. | Arboreous | 1 |
| Olea europaea | Arboreous | 2 |
| Phagnalon saxatile | Shrubby | 3 |
| Phillyrea angustifolia | Shrubby | 4 |
| Pinus halepensis | Arboreous | 4 |
| Pistacia lentiscus | Shrubby | 1 |
| Quercus coccifera | Arboreous | 3 |
| Quercus ilex | Arboreous | 4 |
| Rhamnus oleoides | Shrubby | 2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pareja, J.; Baraza, E.; Ibáñez, M.; Domenech, O.; Bartolomé, J. The Role of Feral Goats in Maintaining Firebreaks by Using Attractants. Sustainability 2020, 12, 7144. https://0-doi-org.brum.beds.ac.uk/10.3390/su12177144
AMA Style
Pareja J, Baraza E, Ibáñez M, Domenech O, Bartolomé J. The Role of Feral Goats in Maintaining Firebreaks by Using Attractants. Sustainability. 2020; 12(17):7144. https://0-doi-org.brum.beds.ac.uk/10.3390/su12177144
Chicago/Turabian StylePareja, Javier, Elena Baraza, Miguel Ibáñez, Oriol Domenech, and Jordi Bartolomé. 2020. "The Role of Feral Goats in Maintaining Firebreaks by Using Attractants" Sustainability 12, no. 17: 7144. https://0-doi-org.brum.beds.ac.uk/10.3390/su12177144
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.