Hidden Engineers and Service Providers: Earthworms in Agricultural Land-Use Types of South Tyrol, Italy



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Field Sampling
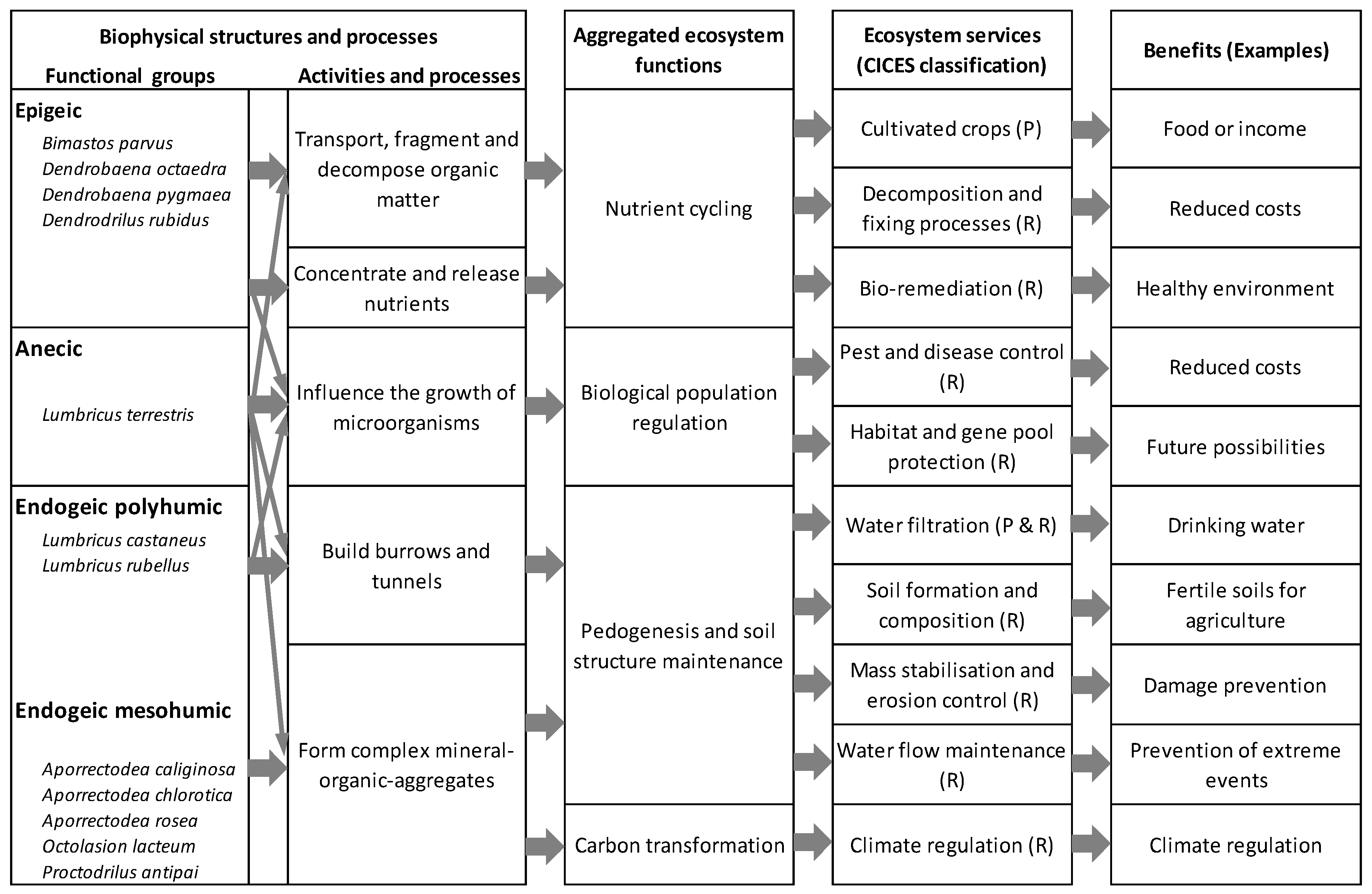
2.2. Functional Earthworm Groups and Ecosystem Services
2.3. Economic Value of Earthworm and Related Ecosystem Services
2.4. Statistical and Spatial Analysis
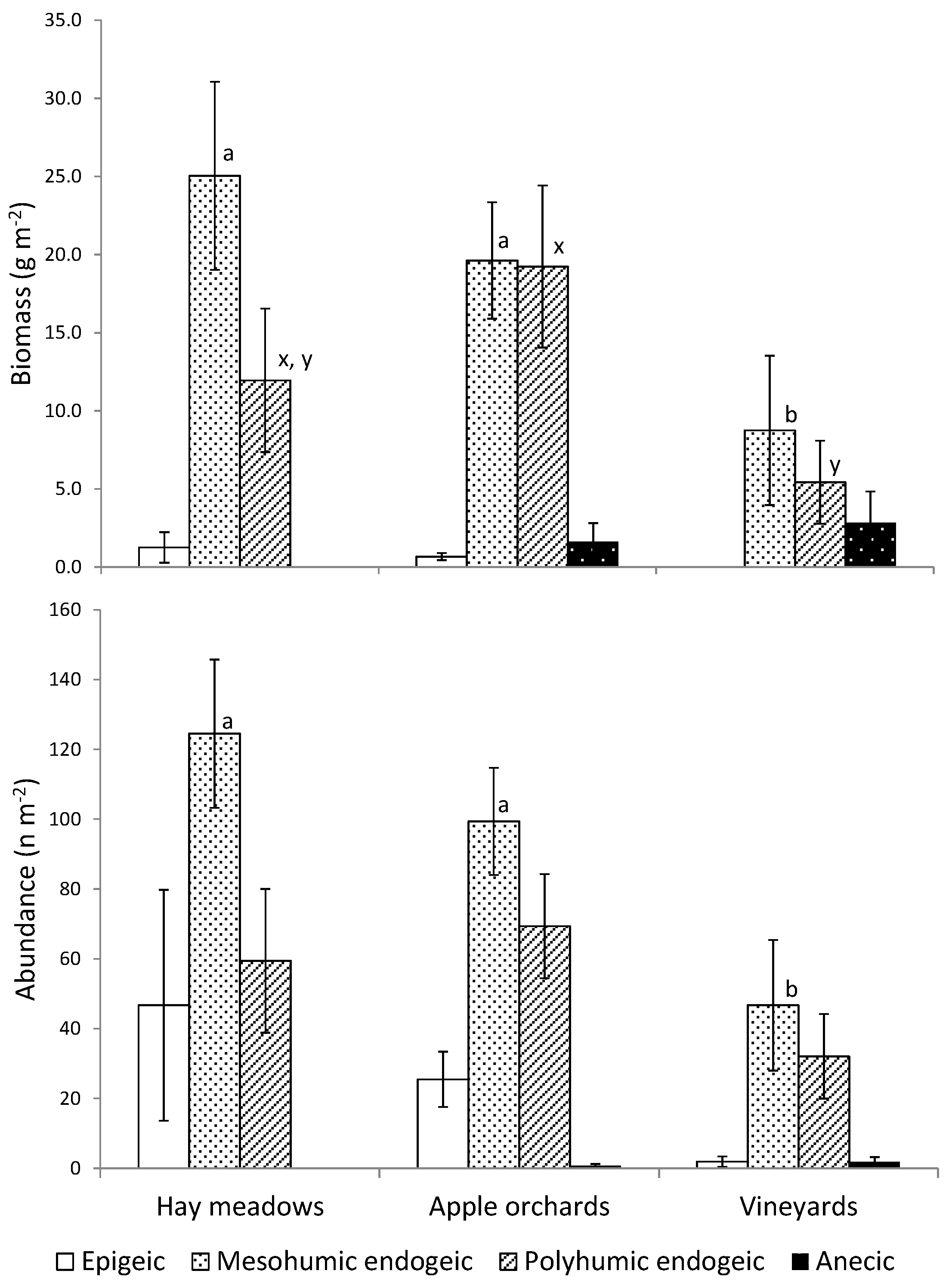
3. Results
4. Discussion
4.1. Land-Use and Earthworm Communities
4.2. Earthworm as Indicator for Sustainable Land Use
4.3. ES and Economic Valuation
4.4. Conclusion and Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darwin, C. The Formation of Vegetable Mould, Through the Action of Worms with Observations on Their Habits; John Murray: London, UK, 1881. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as Ecosystem Engineers. OIKOS 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Bartlett, M.D.; Briones, M.J.I.; Neilson, R.; Schmidt, O.; Spurgeon, D.; Creamer, R.E. A critical review of current methods in earthworm ecology: From individuals to populations. Eur. J. Soil Biol. 2010, 46, 67–73. [Google Scholar] [CrossRef]
- Blouin, M.; Sery, N.; Cluzeau, D.; Brun, J.-J.; Bédécarrats, A. Balkanized Research in Ecological Engineering Revealed by a Bibliometric Analysis of Earthworms and Ecosystem Services. Environ. Manag. 2013, 52, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusaro, S.; Gavinelli, F.; Lazzarini, F.; Paoletti, M.G. Soil Biological Quality Index based on earthworms (QBS-e). A new way to use earthworms as bioindicators in agroecosystems. Ecol. Indic. 2018, 93, 1276–1292. [Google Scholar] [CrossRef]
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: Managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Kibblewhite, M.G.; Ritz, K.; Swift, M.J. Soil health in agricultural systems. Philos. Trans. R. Soc. B 2008, 363, 685–701. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, M.G. The role of earthworms for assessment of sustainability and as bioindicators. Agric. Ecosyst. Environ. 1999, 74, 137–155. [Google Scholar] [CrossRef]
- Bispo, A.; Cluzeau, D.; Creamer, R.; Dombos, M.; Graefe, U.; Krogh, P.H.; Sousa, J.P.; Peres, G.; Rutgers, M.; Winding, A.; et al. Indicators for Monitoring Soil Biodiversity. Integr. Environ. Assess. Manag. 2009, 5, 717–719. [Google Scholar] [CrossRef] [Green Version]
- Turbé, A.; De Toni, A.; Benito, P.; Lavelle, P.; Ruiz, N.; Van der Putten, W.H.; Labouze, E.; Mudgal, S. Soil Biodiversity: Functions, Threats and Tools for Policy Makers; Report for European Commission; DG Environment: Brussels, Belgium, 2010. [Google Scholar]
- Van Groenigen, J.W.; Lubbers, I.M.; Vos, H.M.J.; Brown, G.G.; de Deyn, G.B.; van Groenigen, K.J. Earthworms increase plant production: A meta-analysis. Sci. Rep. 2014, 4, 6365. [Google Scholar] [CrossRef] [Green Version]
- Boag, B.; Palmer, L.F.; Neilson, R.; Legg, R.; Chambers, S.J. Distribution, prevalence and intensity of earthworm populations in arable land and grassland in Scotland. Ann. Appl. Biol. 1997, 130, 153–165. [Google Scholar] [CrossRef]
- Paoletti, M.G.; Sommaggio, D.; Favretto, M.R.; Petruzzelli, G.; Pezzarossa, B.; Barbafieri, M. Earthworms as useful bioindicators of agroecosystem sustainability in orchards and vineyards with different inputs. Appl. Soil Ecol. 1998, 10, 137–150. [Google Scholar] [CrossRef]
- Phillips, H.R.P.; Guerra, C.A.; Bartz, M.L.C.; Briones, M.J.I.; Brown, G.; Crowther, T.W.; Ferlian, O.; Gongalsky, K.B.; van den Hoogen, J.; Krebs, J.; et al. Global distribution of earthworm diversity. Science 2019, 366, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.C.; Harrison, P.A.; Pérez Soba, M.; Archaux, F.; Blicharska, M.; Egoh, B.N.; Erős, T.; Fabrega Domenech, N.; György, Á.; Haines-Young, R.; et al. How natural capital delivers ecosystem services: A typology derived from a systematic review. Ecosyst. Serv. 2017, 26, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.G.; McSwiney, C.P.; Grandy, A.S.; Suwanwaree, P.; Snider, R.M.; Robertson, G.P. Diversity and abundance of earthworms across an agricultural land-use intensity gradient. Soil Tillage Res. 2008, 100, 83–88. [Google Scholar] [CrossRef]
- Rutgers, M.; van Wijnen, H.J.; Schouten, A.J.; Mulder, C.; Kuiten, A.M.P.; Brussaard, L.; Breure, A.M. A method to assess ecosystem services developed from soil attributes with stakeholders and data of four arable farms. Sci. Total Environ. 2012, 415, 39–48. [Google Scholar] [CrossRef]
- Spurgeon, D.; Keith, A.; Schmidt, O.; Lammertsma, D.; Faber, J. Land-use and land-management change: Relationships with earthworm and fungi communities and soil structural properties. BMC Ecol. 2013, 13, 46. [Google Scholar] [CrossRef] [Green Version]
- Keith, A.M.; Robinson, D.A. Earthworms as Natural Capital: Ecosystem Service Providers in Agricultural Soils. Economol. J. 2012, 2, 91–99. [Google Scholar]
- Dominati, E.; Patterson, M.; Mackay, A. A framework for classifying and quantifying the natural capital and ecosystem services of soils. Ecol. Econ. 2010, 69, 1858–1868. [Google Scholar] [CrossRef]
- Haida, C.; Rüdisser, J.; Tappeiner, U. Ecosystem services in mountain regions: Experts’ perceptions and research intensity. Reg. Environ. Chang. 2015, 1–16. [Google Scholar] [CrossRef]
- Robinson, D.A.; Hockley, N.; Dominati, E.; Lebron, I.; Scow, K.M.; Reynolds, B.; Emmett, B.A.; Keith, A.M.; de Jonge, L.W.; Schjønning, P.; et al. Natural Capital, Ecosystem Services, and Soil Change: Why Soil Science Must Embrace an Ecosystems Approach. Vadose Zone J. 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; Van der Putten, W.H.; Wall, D.H. Ecological Linkages Between Aboveground and Belowground Biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Shipitalo, M.; Le Bayon, R.-C. Quantifying the Effects of Earthworms on Soil Aggregation and Porosity. In Earthworm Ecology, 2nd ed.; Edwards, C.A., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 183–200. ISBN 9780849318191. [Google Scholar]
- Johnston, A.S.; Sibly, R.M.; Hodson, M.E.; Alvarez, T.; Thorbek, P. Effects of agricultural management practices on earthworm populations and crop yield: Validation and application of a mechanistic modelling approach. J. Appl. Ecol. 2015, 52, 1334–1342. [Google Scholar] [CrossRef] [Green Version]
- Brussaard, L. Ecosystem Services Provided by the Soil Biota. In Soil Ecology and Ecosystem Services, 1st ed.; Wall, D.H., Ed.; Oxford University Press: Oxford, UK, 2012; pp. 45–58. ISBN 9780199575923. [Google Scholar]
- Luck, G.W.; Harrington, R.; Harrison, P.A.; Kremen, C.; Berry, P.M.; Bugter, R.; Dawson, T.R.; de Bello, F.; Díaz, S.; Feld, C.K.; et al. Quantifying the Contribution of Organisms to the Provision of Ecosystem Services. BioScience 2009, 59, 223–235. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S., III; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of Biodiversity on Ecosystem Functioning: A Consensus of Current Knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Blouin, M.; Hodson, M.E.; Delgado, E.A.; Baker, G.; Brussaard, L.; Butt, K.R.; Dai, J.; Dendooven, L.; Peres, G.; Tondoh, J.E.; et al. A review of earthworm impact on soil function and ecosystem services. Eur. J. Soil Sci. 2013, 64, 161–182. [Google Scholar] [CrossRef]
- Barrios, E. Soil biota, ecosystem services and land productivity. Ecol. Econ. 2007, 64, 269–285. [Google Scholar] [CrossRef]
- ASTAT. Statistisches Jahrbuch für Südtirol. Annuario Statistico della Provincia di Bolzano; ASTAT: Bolzano, Italy, 2014. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014; ISBN 978-92-5-108370-3. [Google Scholar]
- Meyer, E. Bodenzoologische Untersuchungen im Rahmen des Projektes “International Cooperative Programme on Assessment and Monitoring of Air Pollution Effects on Forests” im Auftrag des Forstwirtschaftsinspektorats Bozen und des I’Istituto Agrario di San Michele all’Adige (Settore Foreste Ambiente). Bericht über Arbeiten und Ergebnisse aus dem Jahr 1993; University of Innsbruck: Innsbruck, Austria, 1994. [Google Scholar]
- Rüdisser, J.; Tasser, E.; Peham, T.; Meyer, E.; Tappeiner, U. The dark side of biodiversity: Spatial application of the biological soil quality indicator (BSQ). Ecol. Indic. 2015, 53, 240–246. [Google Scholar] [CrossRef]
- Haines-Young, R.H.; Potschin, M.B. Methodologies for Defining and Assessing Ecosystem Services. Final Report; Project Code C08-0170-0062; University of Nottingham: Nottingham, UK, 2009. [Google Scholar]
- De Groot, R.S.; Alkemade, R.; Braat, L.; Hein, L.; Willemen, L. Challenges in integrating the concept of ecosystem services and values in landscape planning, management and decision making. Ecol. Complex. 2010, 7, 260–272. [Google Scholar] [CrossRef]
- Boyd, J.; Banzhaf, S. What are ecosystem services? The need for standardized environmental accounting units. Ecol. Econ. 2007, 63, 616–626. [Google Scholar] [CrossRef] [Green Version]
- Fisher, B.; Turner, R. Ecosystem services: Classification for valuation. Biol. Conserv. 2008, 141, 1167–1169. [Google Scholar] [CrossRef]
- Burkhard, B.; Kroll, F.; Nedkov, S.; Müller, F. Mapping ecosystem service supply, demand and budgets. Ecosyst. Serv. 2012, 21, 17–29. [Google Scholar] [CrossRef]
- Crossman, N.D.; Burkhard, B.; Nedkov, S.; Willemen, L.; Petz, K.; Palomo, I.; Drakou, E.G.; Martín-Lopez, B.; McPhearson, T.; Boyanova, K.; et al. A blueprint for mapping and modelling ecosystem services. Ecosyst. Serv. 2013, 4, 4–14. [Google Scholar] [CrossRef]
- Bouché, M.B. Strategies lombriciennes. Ecol. Bull. 1977, 122–132. [Google Scholar] [CrossRef]
- Curry, J.P. Functional interactions between earthworms, microorganisms, organic matter, and plants. In Earthworm Ecology, 2nd ed.; Edwards, C.A., Ed.; CRC Press: Boca Raton, FL, USA, 2004; ISBN 9780849318191. [Google Scholar]
- Webb, C.T.; Hoeting, J.A.; Ames, G.M.; Pyne, M.I.; LeRoy Poff, N. A structured and dynamic framework to advance traits-based theory and prediction in ecology. Ecol. Lett. 2010, 13, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Briones, M.J. Soil fauna and soil functions: A jigsaw puzzle. Front. Environ. Sci. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.E. Earthworms. Their Ecology and Relationships with Soils and Land Use; Academic Press: Orlando, FL, USA, 1985; ISBN 0124408605. [Google Scholar]
- Bernard, L.; Chapuis-Lardy, L.; Razafimbelo, T.; Razafindrakoto, M.; Pablo, A.-L.; Legname, E.; Poulain, J.; Bruls, T.; O’Donohue, M.; Brauman, A.; et al. Endogeic earthworms shape bacterial functional communities and affect organic matter mineralization in a tropical soil. ISME J. 2012, 6, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.A.; Bohlen, P.J. Biology and Ecology of Earthworms, 3rd ed.; Chapman & Hall: London, UK, 1996; ISBN 0412561603. [Google Scholar]
- Lavelle, P.; Spain, A. Soil Ecology. Springer: Dordrecht, The Netherlands, 2005; ISBN 978-0-7923-7123-6. [Google Scholar]
- Römbke, J.; Jansch, S.; Didden, W. The use of earthworms in ecological soil classification and assessment concepts. Ecotoxicol. Environ. Saf. 2005, 62, 249–265. [Google Scholar] [CrossRef]
- Brown, G.G.; Edwards, C.A.; Brussaard, L. How Earthworms Affect Plant Growth: Burrowing into the Mechanism. In Earthworm Ecology, 2nd ed.; Edwards, C.A., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 13–42. ISBN 9780849318191. [Google Scholar]
- Fischer, C.; Roscher, C.; Jensen, B.; Eisenhauer, N.; Baade, J.; Attinger, S.; Scheu, S.; Weisser, W.W.; Schumacher, J.; Hildebrandt, A. How Do Earthworms, Soil Texture and Plant Composition Affect Infiltration along an Experimental Plant Diversity Gradient in Grassland? PLoS ONE 2014, 9, e98987. [Google Scholar] [CrossRef]
- Van Schaik, L.; Palm, J.; Klaus, J.; Zehe, E.; Schröder, B. Linking spatial earthworm distribution to macropore numbers and hydrological effectiveness. Ecohydrology 2014, 7, 401–408. [Google Scholar] [CrossRef]
- Andriuzzi, W.; Pulleman, M.; Schmidt, O.; Faber, J.; Brussaard, L. Anecic earthworms (Lumbricus terrestris) alleviate negative effects of extreme rainfall events on soil and plants in field mesocosms. Plant Soil 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Porter, J.; Costanza, R.; Sandhu, H.; Sigsgaard, L.; Wratten, S. The Value of Producing Food, Energy, and Ecosystem Services within an Agro-Ecosystem. Ambio 2009, 38, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, H.S.; Wratten, S.D.; Cullen, R.; Case, B. The future of farming: The value of ecosystem services in conventional and organic arable land. An experimental approach. Ecol. Econ. 2008, 64, 835–848. [Google Scholar] [CrossRef]
- Simonsen, J.; Posner, J.; Rosemeyer, M.; Baldock, J. Endogeic and anecic earthworm abundance in six Midwestern cropping systems. Appl. Soil Ecol. 2010, 44, 147–155. [Google Scholar] [CrossRef]
- Van Zwieten, L.; Rust, J.; Kingston, T.; Merrington, G.; Morris, S. Influence of copper fungicide residues on occurrence of earthworms in avocado orchard soils. Sci. Total Environ. 2004, 329, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Komárek, M.; Čadková, E.; Chrastný, V.; Bordas, F.; Bollinger, J.-C. Contamination of vineyard soils with fungicides: A review of environmental and toxicological aspects. Environ. Int. 2010, 36, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.; Bauer, T.; Strauss, P.; Kratschmer, S.; Paredes, D.; Popescu, D.; Landa, B.; Guzmán, G.; Gómez, J.A.; Guernion, M.; et al. Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: A meta-analysis. J. Appl. Ecol. 2018, 55, 2484–2495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchholz, J.; Querner, P.; Paredes, D.; Bauer, T.; Strauss, P.; Guernion, M.; Scimia, J.; Cluzeau, D.; Burel, F.; Kratschmer, S.; et al. Soil biota in vineyards are more influenced by plants and soil quality than by tillage intensity or the surrounding landscape. Sci. Rep. 2017, 7, 17445. [Google Scholar] [CrossRef] [Green Version]
- Paiola, A.; Assandri, G.; Brambilla, M.; Zottini, M.; Pedrini, P.; Nascimbene, J. Exploring the potential of vineyards for biodiversity conservation and delivery of biodiversity-mediated ecosystem services: A global-scale systematic review. Ecosyst. Funct. Ecosyst. Serv. Biodivers. Ecol. Risk Assess. 2020, 706, 135839. [Google Scholar] [CrossRef]
- Stellin, F.; Gavinelli, F.; Stevanato, P.; Concheri, G.; Squartini, A.; Paoletti, M.G. Effects of different concentrations of glyphosate (Roundup 360®) on earthworms (Octodrilus complanatus, Lumbricus terrestris and Aporrectodea caliginosa) in vineyards in the North-East of Italy. Appl. Soil Ecol. 2018, 123, 802–808. [Google Scholar] [CrossRef]
- Zaller, J.G.; Heigl, F.; Ruess, L.; Grabmaier, A. Glyphosate herbicide affects belowground interactions between earthworms and symbiotic mycorrhizal fungi in a model ecosystem. Sci. Rep. 2014, 4, 5634. [Google Scholar] [CrossRef] [Green Version]
- Gaupp-Berghausen, M.; Hofer, M.; Rewald, B.; Zaller, J.G. Glyphosate-based herbicides reduce the activity and reproduction of earthworms and lead to increased soil nutrient concentrations. Sci. Rep. 2015, 5, 12886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavelle, P.; Rodríguez, N.; Arguello, O.; Bernal, J.; Botero, C.; Chaparro, P.; Gómez, Y.; Gutiérrez, A.; Hurtado, M.d.P.; Loaiza, S.; et al. Soil ecosystem services and land use in the rapidly changing Orinoco River Basin of Colombia. Agric. Ecosyst. Environ. 2014, 185, 106–117. [Google Scholar] [CrossRef]
- Palm, J.; van Schaik, N.L.M.B.; Schröder, B. Modelling distribution patterns of anecic, epigeic and endogeic earthworms at catchment-scale in agro-ecosystems. Pedobiologia 2013, 56, 23–31. [Google Scholar] [CrossRef]
- Birkhofer, K.; Schöning, I.; Alt, F.; Herold, N.; Klarner, B.; Maraun, M.; Marhan, S.; Oelmann, Y.; Wubet, T.; Yurkov, A.; et al. General Relationships between Abiotic Soil Properties and Soil Biota across Spatial Scales and Different Land-Use Types. PLoS ONE 2012, 7, e43292. [Google Scholar] [CrossRef]
- Jaensch, S.; Steffens, L.; Hoefer, H.; Horak, F.; Ross-Nickoll, M.; Russell, D.; Toschki, A.; Roembke, J. State of knowledge of earthworm communities in German soils as a basis for biological soil quality assessment. Soil Org. 2013, 85, 215–233. [Google Scholar]
- Tsiafouli, M.A.; Thébault, E.; Sgardelis, S.P.; de Ruiter, P.C.; Van der Putten, W.H.; Birkhofer, K.; Hemerik, L.; de Vries, F.T.; Bardgett, R.D.; Brady, M.V.; et al. Intensive agriculture reduces soil biodiversity across Europe. Glob. Chang. Biol. 2015, 21, 973–985. [Google Scholar] [CrossRef]
- De Vries, F.T.; Thébault, E.; Liiri, M.; Birkhofer, K.; Tsiafouli, M.A.; Bjørnlund, L.; Bracht Jørgensen, H.; Brady, M.V.; Christensen, S.; de Ruiter, P.C.; et al. Soil food web properties explain ecosystem services across European land use systems. Proc. Natl. Acad. Sci. USA 2013, 110, 14296–14301. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, B.; Faber, J.; Bloem, J. Applying Soil Health Indicators to Encourage Sustainable Soil Use: The Transition from Scientific Study to Practical Application. Sustainability 2018, 10, 3021. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga, M.C.; Feijoo, M.A.; Quintero, H.; Aldana, N.J.; Carvajal, A.F. Farmers’ perceptions of earthworms and their role in soil. Appl. Soil Ecol. 2013, 69, 61–68. [Google Scholar] [CrossRef]
- Velasquez, E.; Lavelle, P. Soil macrofauna as an indicator for evaluating soil based ecosystem services in agricultural landscapes. Acta Oecol. 2019, 100, 103446. [Google Scholar] [CrossRef]
- Fründ, H.-C.; Graefe, U.; Tischer, S. Earthworms as Bioindicators of Soil Quality. In Biology of Earthworms; Karaca, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 261–278. ISBN 978-3-642-14635-0. [Google Scholar]
- Pulleman, M.; Creamer, R.; Hamer, U.; Helder, J.; Pelosi, C.; Pérès, G.; Rutgers, M. Soil biodiversity, biological indicators and soil ecosystem services—An overview of European approaches. Curr. Opin. Environ. Sustain. 2012, 4, 529–538. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Römbke, J.; Schmelz, R.M.; Scheffczyk, A.; Faber, J.H.; Bloem, J.; Pérès, G.; Cluzeau, D.; Chabbi, A.; Suhadolc, M.; et al. Selecting cost effective and policy-relevant biological indicators for European monitoring of soil biodiversity and ecosystem function. Ecol. Indic. 2016, 69, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Stone, D.; Ritz, K.; Griffiths, B.G.; Orgiazzi, A.; Creamer, R.E. Selection of biological indicators appropriate for European soil monitoring. Appl. Soil Ecol. 2016, 97, 12–22. [Google Scholar] [CrossRef]
- Rutgers, M.; Orgiazzi, A.; Gardi, C.; Römbke, J.; Jänsch, S.; Keith, A.M.; Neilson, R.; Boag, B.; Schmidt, O.; Murchie, A.K.; et al. Mapping earthworm communities in Europe. Appl. Soil Ecol. 2016, 97, 98–111. [Google Scholar] [CrossRef]
- Schon, N.L.; Mackay, A.D.; Gray, R.A.; van Koten, C.; Dodd, M.B. Influence of earthworm abundance and diversity on soil structure and the implications for soil services throughout the season. Pedobiologia 2017, 62, 41–47. [Google Scholar] [CrossRef]
- Schon, N.L.; Mackay, A.D.; Gray, R.A.; Dodd, M.B.; van Koten, C. Quantifying dung carbon incorporation by earthworms in pasture soils. Eur. J. Soil Sci. 2015, 66, 348–358. [Google Scholar] [CrossRef]
- Pascual, U.; Termansen, M.; Hedlund, K.; Brussaard, L.; Faber, J.H.; Foudi, S.; Lemanceau, P.; Jørgensen, S.L. On the value of soil biodiversity and ecosystem services. Ecosyst. Serv. 2015, 15, 11–18. [Google Scholar] [CrossRef]
- Jones, L.; Norton, L.; Austin, Z.; Browne, A.L.; Donovan, D.; Emmett, B.A.; Grabowski, Z.; Howard, D.C.; Jones, J.; Kenter, J.; et al. Stocks and flows of natural and human-derived capital in ecosystem services. Land Use Policy 2016, 52, 151–162. [Google Scholar] [CrossRef]
- Adhikari, K.; Hartemink, A.E. Linking soils to ecosystem services—A global review. Geoderma 2016, 262, 101–111. [Google Scholar] [CrossRef]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G.A. An Underground Revolution: Biodiversity and Soil Ecological Engineering for Agricultural Sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Robinson, D.A.; Fraser, I.; Dominati, E.J.; Davíðsdóttir, B.; Jónsson, J.O.G.; Jones, L.; Jones, S.B.; Tuller, M.; Lebron, I.; Bristow, K.L.; et al. On the Value of Soil Resources in the Context of Natural Capital and Ecosystem Service Delivery. Soil Sci. Soc. Am. J. 2014, 78, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Dominati, E.; Mackay, A.; Green, S.; Patterson, M. A soil change-based methodology for the quantification and valuation of ecosystem services from agro-ecosystems: A case study of pastoral agriculture in New Zealand. Ecol. Econ. 2014, 100, 119–129. [Google Scholar] [CrossRef]
- De Groot, R. Function-analysis and valuation as a tool to assess land use conflicts in planning for sustainable, multi-functional landscapes: Landscapes and sustainability. Landsc. Urban Plan. 2006, 75, 175–186. [Google Scholar] [CrossRef]
- Spangenberg, J.H.; Settele, J. Precisely incorrect? Monetising the value of ecosystem services: Ecosystem Services—Bridging Ecology, Economy and Social Sciences. Ecol. Complex. 2010, 7, 327–337. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land-Use Type | n | Abundance (n m−2) | Biomass (g m−2) | Species Richness | H’ |
|---|---|---|---|---|---|
| Hay meadows | 15 | 239 ± 185 a | 40.2 ± 31.5 a | 3.3 ± 1.4 a | 1.2 a |
| Apple orchards | 20 | 208 ± 108 a | 43.1 ± 28.7 a | 3.6 ± 1.3 a | 1.2 a |
| Vineyards | 15 | 83 ± 118 b | 17.3 ± 34.5 b | 1.2 ± 1.3 b | 0.4 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rüdisser, J.; Tasser, E.; Peham, T.; Meyer, E.; Tappeiner, U. Hidden Engineers and Service Providers: Earthworms in Agricultural Land-Use Types of South Tyrol, Italy. Sustainability 2021, 13, 312. https://0-doi-org.brum.beds.ac.uk/10.3390/su13010312
Rüdisser J, Tasser E, Peham T, Meyer E, Tappeiner U. Hidden Engineers and Service Providers: Earthworms in Agricultural Land-Use Types of South Tyrol, Italy. Sustainability. 2021; 13(1):312. https://0-doi-org.brum.beds.ac.uk/10.3390/su13010312
Chicago/Turabian StyleRüdisser, Johannes, Erich Tasser, Thomas Peham, Erwin Meyer, and Ulrike Tappeiner. 2021. "Hidden Engineers and Service Providers: Earthworms in Agricultural Land-Use Types of South Tyrol, Italy" Sustainability 13, no. 1: 312. https://0-doi-org.brum.beds.ac.uk/10.3390/su13010312