
The Environmental Impact of Ecological Intensification in Soybean Cropping Systems in the U.S. Upper Midwest


 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cropping Sequences, Site Characteristics and Field Management
2.2. Life Cycle Assessment
2.3. Uncertainty Assessment
3. Results and Discussion
3.1. Land-Based Functional Unit (FU1)
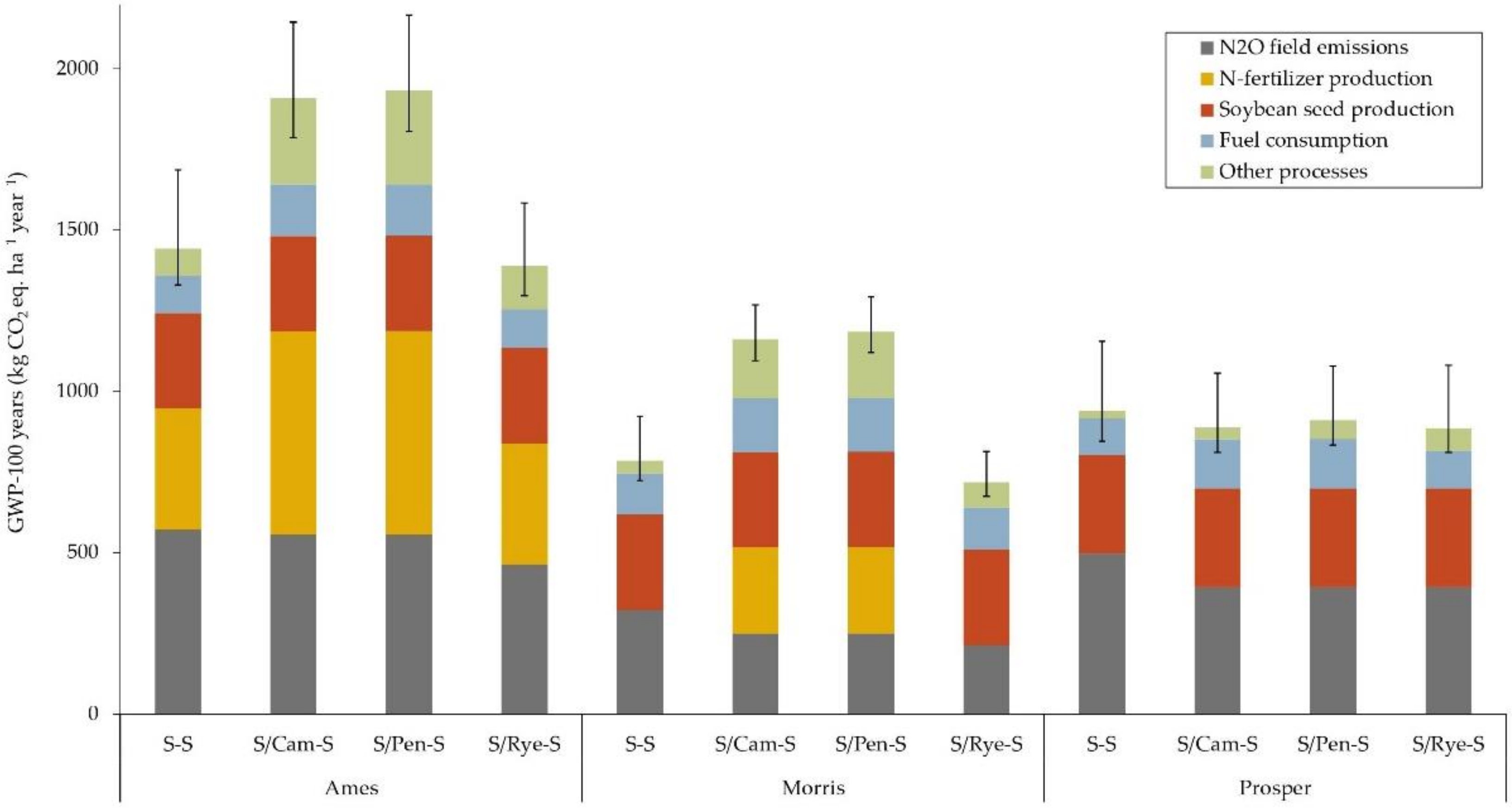
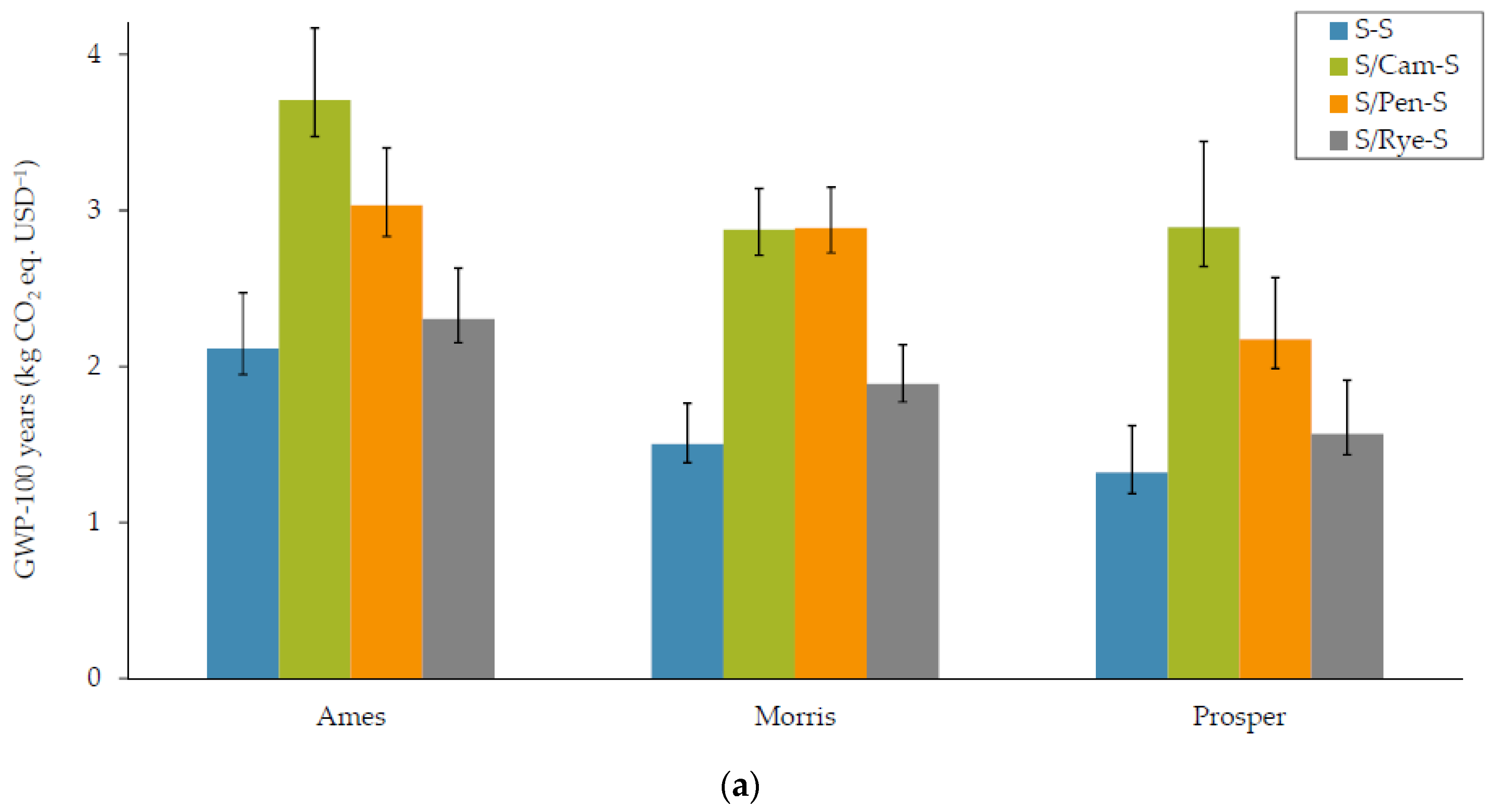
3.1.1. Global Warming Potential (GWP)
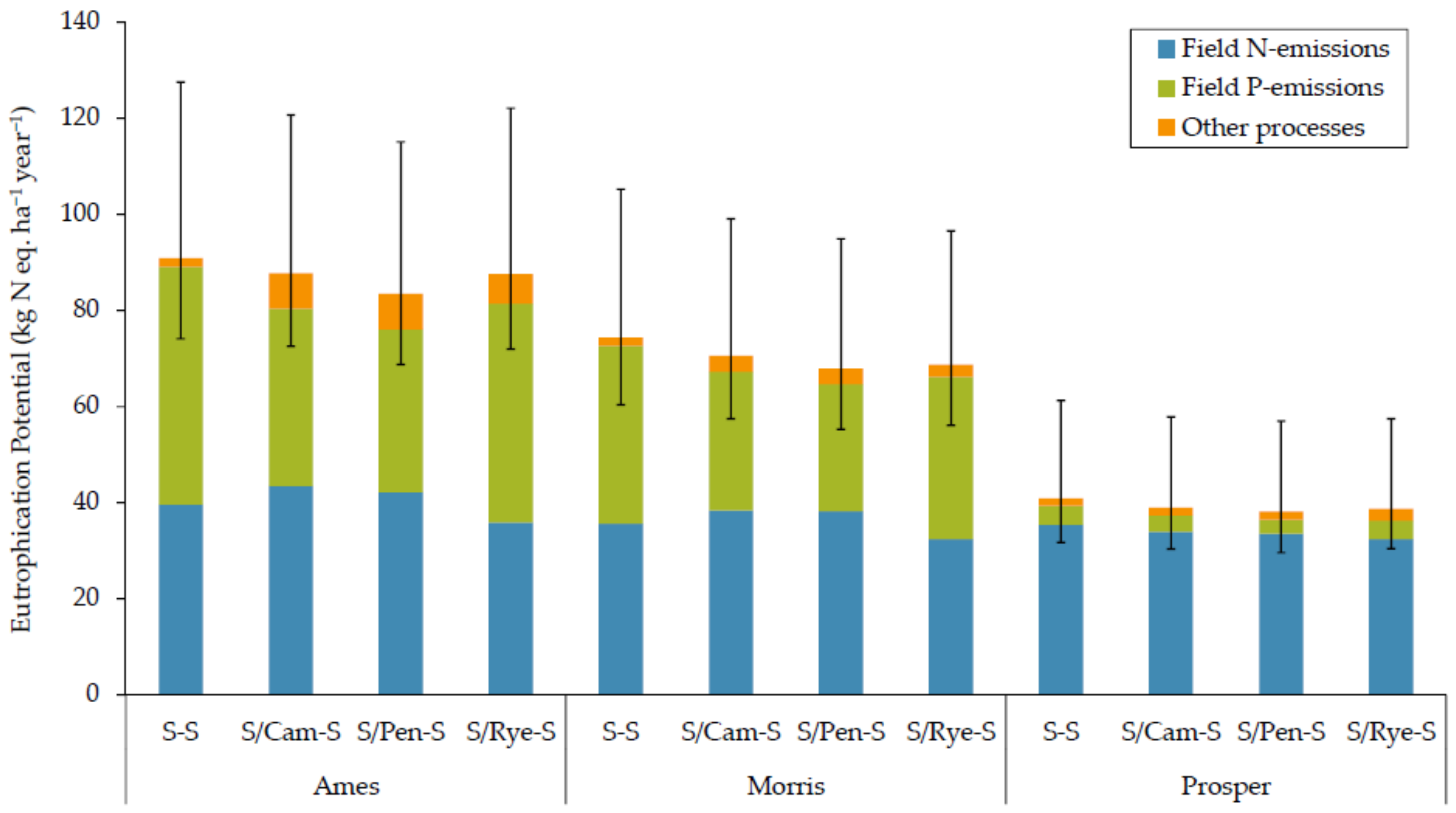
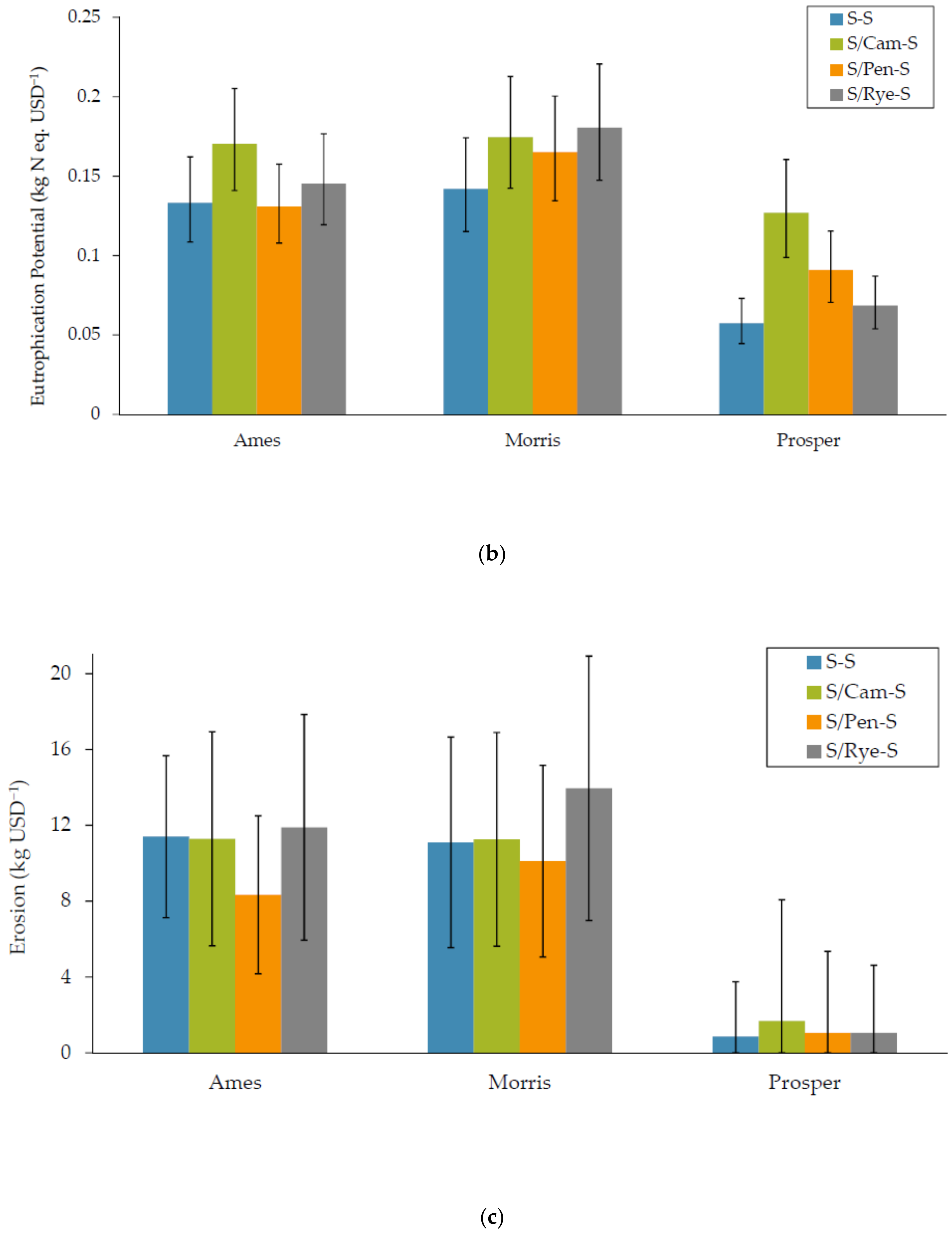
3.1.2. Eutrophication
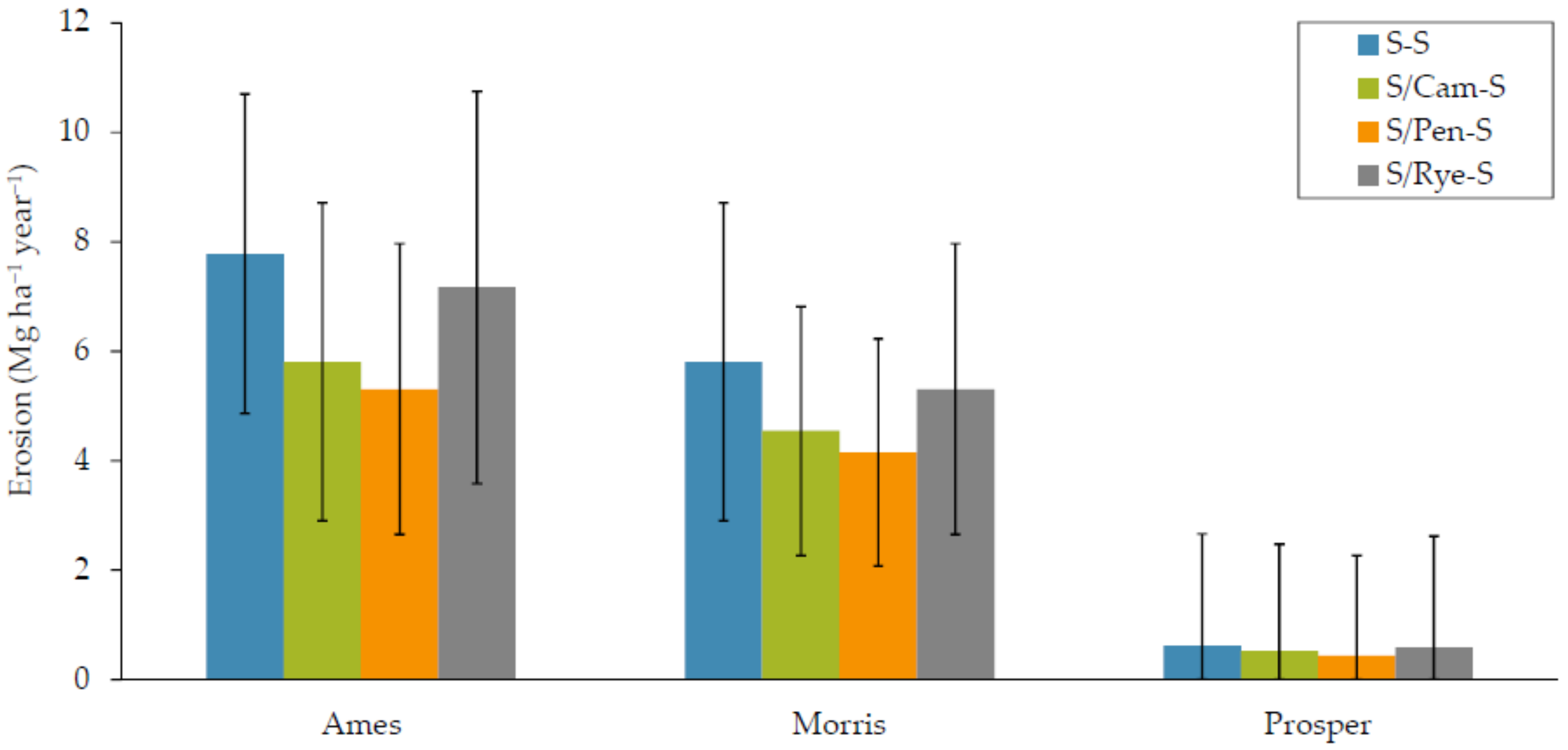
3.1.3. Soil Water Erosion
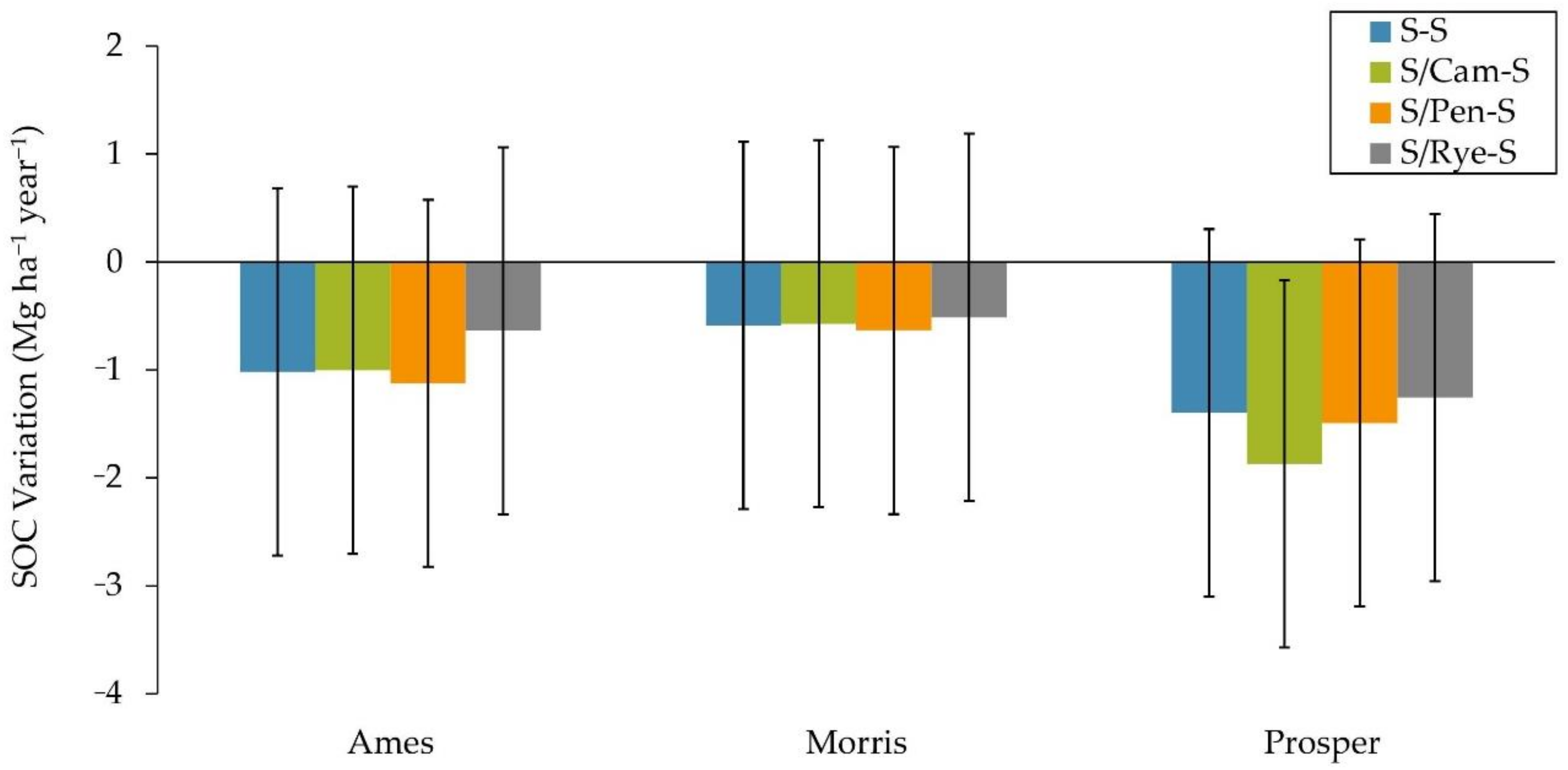
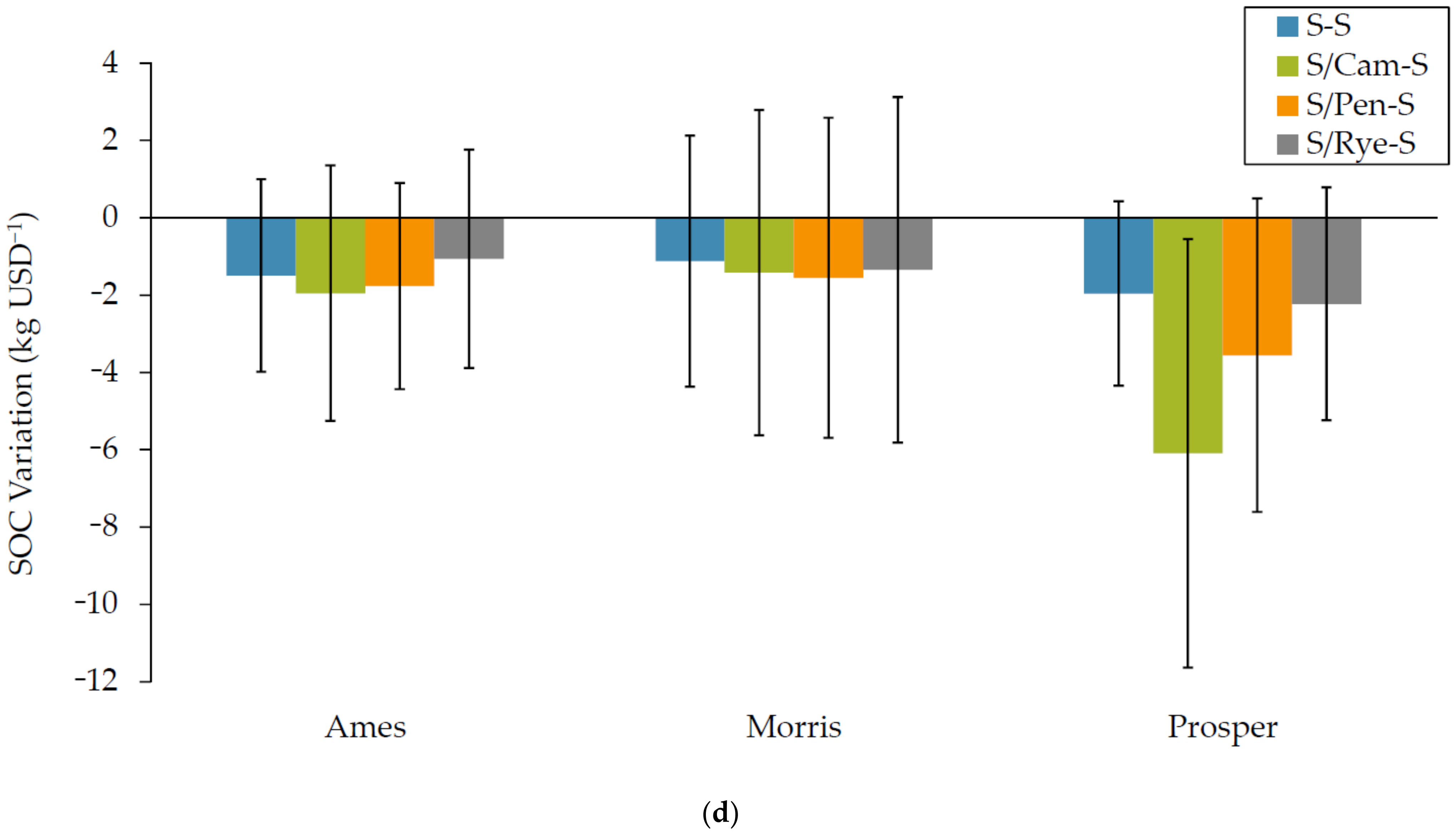
3.1.4. Soil Organic Carbon (SOC) Variation
3.2. Economic Functional Unit (FU2)
3.3. The Environmental Impact of the “Soybean N-Credit”
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cassman, K.G. Ecological intensification of cereal production systems: Yield potential, soil quality, and precision agriculture. Proc. Natl. Acad. Sci. USA 1999, 96, 5952–5959. [Google Scholar] [CrossRef] [Green Version]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Pervaiz, Z.H.; Iqbal, J.; Zhang, Q.; Chen, D.; Wei, H.; Saleem, M. Continuous cropping alters multiple biotic and abiotic indicators of soil health. Soil Syst. 2020, 4, 59. [Google Scholar] [CrossRef]
- Karlen, D.L.; Hurley, E.G.; Andrews, S.S.; Cambardella, C.A.; Meek, D.W.; Duffy, M.D.; Mallarino, A.P. Crop rotation effects on soil quality at three northern corn/soybean belt locations. Agron. J. 2006, 98, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.; Miao, S.; Li, Y.; Zhong, X. Chemical composition of soil organic carbon changed by long-term monoculture cropping system in Chinese black soil. Plant Soil Environ. 2018, 64, 557–563. [Google Scholar] [CrossRef] [Green Version]
- Greer, K.; Martins, C.; White, M.; Pittelkow, C.M. Assessment of high-input soybean management in the US Midwest: Balancing crop production with environmental performance. Agric. Ecosyst. Environ. 2020, 292, 106811. [Google Scholar] [CrossRef]
- Wittwer, R.A.; Dorn, B.; Jossi, W.; Van Der Heijden, M.G.A. Cover crops support ecological intensification of arable cropping systems. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G.A. An underground revolution: Biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Kleijn, D.; Bommarco, R.; Fijen, T.P.M.; Garibaldi, L.A.; Potts, S.G.; van der Putten, W.H. Ecological intensification: Bridging the gap between science and practice. Trends Ecol. Evol. 2019, 34, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Snapp, S.S.; Swinton, S.M.; Labarta, R.; Mutch, D.; Black, J.R.; Leep, R.; Nyiraneza, J.; O’Neil, K. Evaluating cover crops for benefits, costs and performance within cropping system niches. Agron. J. 2005, 97, 322–332. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Shaver, T.M.; Lindquist, J.L.; Shapiro, C.A.; Elmore, R.W.; Francis, C.A.; Hergert, G.W. Cover crops and ecosystem services: Insights from studies in temperate soils. Agron. J. 2015, 107, 2449–2474. [Google Scholar] [CrossRef] [Green Version]
- De Baets, S.; Poesen, J.; Meersmans, J.; Serlet, L. Cover crops and their erosion-reducing effects during concentrated flow erosion. Catena 2011, 85, 237–244. [Google Scholar] [CrossRef]
- Marques, M.J.; Ruiz-Colmenero, M.; Bienes, R.; García-Díaz, A.; Sastre, B. Effects of a permanent soil cover on water dynamics and wine characteristics in a steep vineyard in the central Spain. Air Soil Water Res. 2020, 13. [Google Scholar] [CrossRef]
- Rodrigo-Comino, J.; Terol, E.; Mora, G.; Giménez-Morera, A.; Cerdà, A. Vicia sativa Roth. can reduce soil and water losses in recently planted vineyards (Vitis vinifera L.). Earth Syst. Environ. 2020, 4, 827–842. [Google Scholar] [CrossRef]
- Matson, P.A.; Parton, W.J.; Power, A.G.; Swift, M.J. Agricultural intensification and ecosystem properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [Green Version]
- West, T.O.; Post, W.M. Soil organic carbon sequestration rates by tillage and crop rotation. Soil Sci. Soc. Am. J. 2002, 66, 1930–1946. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, M.D.; Tiemann, L.K.; Grandy, A.S. Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol. Appl. 2014, 24, 560–570. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Pervaiz, Z.H.; Contreras, J.; Lindenberger, J.H.; Hupp, B.M.; Chen, D.; Zhang, Q.; Wang, C.; Iqbal, J.; Twigg, P. Cover crop diversity improves multiple soil properties via altering root architectural traits. Rhizosphere 2020, 16, 100248. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kemanian, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J.; et al. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Eberle, C.A.; Thom, M.D.; Nemec, K.T.; Forcella, F.; Lundgren, J.G.; Gesch, R.W.; Riedell, W.E.; Papiernik, S.K.; Wagner, A.; Peterson, D.H.; et al. Using pennycress, camelina, and canola cash cover crops to provision pollinators. Ind. Crop. Prod. 2015, 75, 20–25. [Google Scholar] [CrossRef]
- Moser, B.R. Biodiesel from alternative oilseed feedstocks: Camelina and field pennycress. Biofuels 2012, 3, 193–209. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.; Kumar, A. Development of emission parameters and net energy ratio for renewable diesel from Canola and Camelina. Energy 2013, 58, 426–437. [Google Scholar] [CrossRef]
- Krohn, B.J.; Fripp, M. A life cycle assessment of biodiesel derived from the “niche filling” energy crop camelina in the USA. Appl. Energy 2012, 92, 92–98. [Google Scholar] [CrossRef]
- Fan, J.; Shonnard, D.R.; Kalnes, T.N.; Johnsen, P.B.; Rao, S. A life cycle assessment of pennycress (Thlaspi arvense L.) -derived jet fuel and diesel. Biomass Bioenergy 2013, 55, 87–100. [Google Scholar] [CrossRef]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crop. Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Obour, A.K. Oilseed Camelina (Camelina sativa L Crantz): Production systems, prospects and challenges in the USA Great Plains. Adv. Plants Agric. Res. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- McGinn, M.; Phippen, W.B.; Chopra, R.; Bansal, S.; Jarvis, B.A.; Phippen, M.E.; Dorn, K.M.; Esfahanian, M.; Nazarenus, T.J.; Cahoon, E.B.; et al. Molecular tools enabling pennycress (Thlaspi arvense) as a model plant and oilseed cash cover crop. Plant Biotechnol. J. 2019, 17, 776–788. [Google Scholar] [CrossRef] [Green Version]
- Bergtold, J.S.; Ramsey, S.; Maddy, L.; Williams, J.R. A review of economic considerations for cover crops as a conservation practice. Renew. Agric. Food Syst. 2019, 34, 62–76. [Google Scholar] [CrossRef]
- Dunn, M.; Ulrich-Schad, J.D.; Prokopy, L.S.; Myers, R.L.; Watts, C.R.; Scanlon, K. Perceptions and use of cover crops among early adopters: Findings from a national survey. J. Soil Water Conserv. 2016, 71, 29–40. [Google Scholar] [CrossRef]
- Roesch-Mcnally, G.E.; Basche, A.D.; Arbuckle, J.G.; Tyndall, J.C.; Miguez, F.E.; Bowman, T.; Clay, R. The trouble with cover crops: Farmers’ experiences with overcoming barriers to adoption. Renew. Agric. Food Syst. 2018, 33, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.G.; Gross, K.L.; Robertson, G.P. Effects of crop diversity on agroecosystem function: Crop yield response. Ecosystems 2008, 11, 355–366. [Google Scholar] [CrossRef]
- Gesch, R.W.; Archer, D.W.; Berti, M.T. Dual cropping winter camelina with soybean in the northern corn belt. Agron. J. 2014, 106, 1735–1745. [Google Scholar] [CrossRef]
- Berti, M.; Gesch, R.; Johnson, B.; Ji, Y.; Seames, W.; Aponte, A. Double- and relay-cropping of energy crops in the northern Great Plains, USA. Ind. Crop. Prod. 2015, 75, 26–34. [Google Scholar] [CrossRef]
- Mohammed, Y.A.; Matthees, H.L.; Gesch, R.W.; Patel, S.; Forcella, F.; Aasand, K.; Steffl, N.; Johnson, B.L.; Wells, M.S.; Lenssen, A.W. Establishing winter annual cover crops by interseeding into Maize and Soybean. Agron. J. 2020, 112, 719–732. [Google Scholar] [CrossRef] [Green Version]
- Cubins, J.A.; Wells, M.S.; Frels, K.; Ott, M.A.; Forcella, F.; Johnson, G.A.; Walia, M.K.; Becker, R.L.; Gesch, R.W. Management of pennycress as a winter annual cash cover crop. A review. Agron. Sustain. Dev. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Heaton, E.A.; Schulte, L.A.; Berti, M.; Langeveld, H.; Zegada-Lizarazu, W.; Parrish, D.; Monti, A. Managing a second-generation crop portfolio through sustainable intensifi cation: Examples from the USA and the EU. Biofuels Bioprod. Biorefin. 2013, 7, 702–714. [Google Scholar] [CrossRef]
- Mohammed, Y.A.; Patel, S.; Matthees, H.L.; Lenssen, A.W.; Johnson, B.L.; Scott Wells, M.; Forcella, F.; Berti, M.T.; Gesch, R.W. Soil nitrogen in response to interseeded cover crops in maize-soybean production systems. Agronomy 2020, 10, 1439. [Google Scholar] [CrossRef]
- Patel, S.; Lenssen, A.W.; Moore, K.J.; Mohammed, Y.A.; Gesch, R.W.; Wells, M.S.; Johnson, B.L.; Berti, M.T.; Matthees, H.L. Interseeded pennycress and camelina yield and their influence on row crops. Agron. J. 2021, in press. [Google Scholar]
- ISO. Environmental Management—Life Cycle Assessment—Principles and Framework; ISO 14040:2006; International Organization for Standardization (ISO): Geneva, Switzerland, 2006. [Google Scholar]
- Caffrey, K.R.; Veal, M.W. Conducting an agricultural life cycle assessment: Challenges and perspectives. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Nemecek, T.; Dubois, D.; Huguenin-Elie, O.; Gaillard, G. Life cycle assessment of Swiss farming systems: I. Integrated and organic farming. Agric. Syst. 2011, 104, 217–232. [Google Scholar] [CrossRef]
- Wernet, G.; Bauer, C.; Steubing, B.; Reinhard, J.; Moreno-Ruiz, E.; Weidema, B. The ecoinvent database version 3 (part I): Overview and methodology. Int. J. Life Cycle Assess. 2016, 21, 1218–1230. [Google Scholar] [CrossRef]
- Cecchin, A.; Pourhashem, G.; Gesch, R.W.; Lenssen, A.W.; Mohammed, Y.A.; Patel, S.; Berti, M.T. Environmental trade-offs of relay-cropping winter cover crops with soybean in a maize-soybean cropping system. Agric. Syst. 2021, 189, 103062. [Google Scholar] [CrossRef]
- Bouwman, A.F.; Boumans, L.J.M.; Batjes, N.H. Modeling global annual N2O and NO emissions from fertilized fields. Glob. Biogeochem. Cycles 2002, 16, 28-1–28-9. [Google Scholar] [CrossRef]
- Bouwman, A.F.; Boumans, L.J.M.; Batjes, N.H. Estimation of global NH3 volatilization loss from synthetic fertilizers and animal manure applied to arable lands and grasslands. Glob. Biogeochem. Cycles 2002, 16, 8-1–8-14. [Google Scholar] [CrossRef]
- IPCC Agriculture Forestry and Other Land Use. Chapter 11: N2O emissions from managed soils, and CO2 emissions from lime and urea application. In 2006 IPCC Guidelines for National Greenhouse Gas Inventories; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2006; Volume 4, pp. 1–54. [Google Scholar]
- United States Environmental Protection Agency. Emission Factors for Greenhouse Gas Inventories; US-EPA: Washington, DC, USA, 2018.
- Rosenbaum, R.K.; Anton, A.; Bengoa, X.; Bjørn, A.; Brain, R.; Bulle, C.; Cosme, N.; Dijkman, T.J.; Fantke, P.; Felix, M.; et al. The Glasgow consensus on the delineation between pesticide emission inventory and impact assessment for LCA. Int. J. Life Cycle Assess. 2015, 20, 765–776. [Google Scholar] [CrossRef] [Green Version]
- Xue, X.; Hawkins, T.R.; Ingwersen, W.W.; Smith, R.L. Demonstrating an approach for including pesticide use in life-cycle assessment: Estimating human and ecosystem toxicity of pesticide use in Midwest corn farming. Int. J. Life Cycle Assess. 2015, 20, 1117–1126. [Google Scholar] [CrossRef]
- European Commission. Product Environmental Footprint Category Rules Guidance; European Commission: Brussels, Belgium, 2018. [Google Scholar]
- Nemecek, T.; Schnetzer, J. Methods of Assessment of Direct Field Emissions for LCIS of Agricultural Production Systems; Agroscope Reckenholz-Tänikon Research Station ART: Zurich, Switzerland, 2011. [Google Scholar]
- Nemecek, T.; Schnetzer, J.; Reinhard, J. Updated and harmonised greenhouse gas emissions for crop inventories. Int. J. Life Cycle Assess. 2016, 21, 1361–1378. [Google Scholar] [CrossRef]
- Myhre, G.; Shindell, D.; Bréon, F.-M.; Collins, W.; Fuglestvedt, J.; Huang, J.; Koch, D. Anthropogenic and natural radiative forcing. In Climate Change 2013 the Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: New York, NY, USA, 2013; pp. 659–740. ISBN 9781107415324. [Google Scholar]
- Bare, J. TRACI 2.0: The tool for the reduction and assessment of chemical and other environmental impacts 2.0. Clean Technol. Environ. Policy 2011, 13, 687–696. [Google Scholar] [CrossRef]
- Foster, G.R.; Toy, T.E.; Renard, K.G. Comparison of the USLE, RUSLE1.06c, and RUSLE2 for application to highly disturbed lands. First Interag. Conf. Res. Watersheds 2003, 27, 154–160. [Google Scholar]
- Foster, G.R.; Yoder, D.C.; Weesies, G.A.; McCool, D.K.; McGregor, K.C.; Bingner, R.L. RUSLE2 User’s Guide; USDA—Agricultural research service: Washington, DC, USA, 2003.
- United States Department of Agriculture (USDA). The Wind Erosion Prediction System. WEPS 1.5 User Manual; USDA-ARS Agricultural Systems Research Unit: Fort Collins, CO, USA, 2016.
- i Canals, L.M.; Romanyà, J.; Cowell, S.J. Method for assessing impacts on life support functions (LSF) related to the use of “fertile land” in Life Cycle Assessment (LCA). J. Clean. Prod. 2007, 15, 1426–1440. [Google Scholar] [CrossRef]
- Brandão, M.; Milà i Canals, L.; Clift, R. Soil organic carbon changes in the cultivation of energy crops: Implications for GHG balances and soil quality for use in LCA. Biomass Bioenergy 2011, 35, 2323–2336. [Google Scholar] [CrossRef]
- Goglio, P.; Smith, W.N.; Grant, B.B.; Desjardins, R.L.; McConkey, B.G.; Campbell, C.A.; Nemecek, T. Accounting for soil carbon changes in agricultural life cycle assessment (LCA): A review. J. Clean. Prod. 2015, 104, 23–39. [Google Scholar] [CrossRef]
- Johnson, J.M.F.; Allmaras, R.R.; Reicosky, D.C. Estimating source carbon from crop residues, roots and rhizodeposits using the national grain-yield database. Agron. J. 2006, 98, 622–636. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.M.F.; Papiernik, S.K.; Mikha, M.M.; Spokas, K.A.; Tomer, M.D.; Weyers, S.L. Soil processes and residue harvest management. In Soil Quality Biofuel Production; Taylor & Francis: New York, NY, USA, 2009; pp. 1–44. [Google Scholar]
- Johnson, J.M.F.; Acosta-Martinez, V.; Cambardella, C.A.; Barbour, N.W. Crop and soil responses to using corn stover as a bioenergy feedstock: Observations from the northern us corn belt. Agriculture 2013, 3, 72–89. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.M.F.; Novak, J.M.; Varvel, G.E.; Stott, D.E.; Osborne, S.L.; Karlen, D.L.; Lamb, J.A.; Baker, J.; Adler, P.R. Crop residue mass needed to maintain soil organic carbon levels: Can it be determined? (Special Issue: Crop residue considerations for sustainable bioenergy feedstock supplies.). Bioenergy Res. 2014, 7, 481–490. [Google Scholar] [CrossRef]
- Weidema, B.P.; Wesnæs, M.S. Data quality management for life cycle inventories-an example of using data quality indicators. J. Clean. Prod. 1996, 4, 167–174. [Google Scholar] [CrossRef]
- Goedkoop, M.; Oele, M.; Leijting, J.; Ponsioen, T.; Meijer, E. Introduction to LCA with SimaPro; SimaPro: Amersfoort, The Netherlands, 2016. [Google Scholar]
- United States Department of Agriculture (USDA). Revised Universal Soil Loss Equation version 2 (RUSLE2)—Handbook; United States Department of Agriculture (USDA): Washington, DC, USA, 2001; Volume 2.
- Smith, K.A. Changing views of nitrous oxide emissions from agricultural soil: Key controlling processes and assessment at different spatial scales. Eur. J. Soil Sci. 2017, 68, 137–155. [Google Scholar] [CrossRef]
- Hayatsu, M.; Tago, K.; Saito, M. Various players in the nitrogen cycle: Diversity and functions of the microorganisms involved in nitrification and denitrification. Soil Sci. Plant Nutr. 2008, 54, 33–45. [Google Scholar] [CrossRef]
- Venterea, R.T.; Halvorson, A.D.; Kitchen, N.; Liebig, M.A.; Cavigelli, M.A.; Del Grosso, S.J.; Motavalli, P.P.; Nelson, K.A.; Spokas, K.A.; Singh, B.P.; et al. Challenges and opportunities for mitigating nitrous oxide emissions from fertilized cropping systems. Front. Ecol. Environ. 2012, 10, 562–570. [Google Scholar] [CrossRef]
- Weyers, S.; Thom, M.; Forcella, F.; Eberle, C.; Matthees, H.; Gesch, R.; Ott, M.; Feyereisen, G.; Strock, J.; Wyse, D. Reduced potential for nitrogen loss in cover crop-soybean relay systems in a cold climate. J. Environ. Qual. 2019, 48, 660–669. [Google Scholar] [CrossRef]
- Grossel, A.; Nicoullaud, B.; Bourennane, H.; Lacoste, M.; Guimbaud, C.; Robert, C.; Hénault, C. The effect of tile-drainage on nitrous oxide emissions from soils and drainage streams in a cropped landscape in Central France. Agric. Ecosyst. Environ. 2016, 230, 251–260. [Google Scholar] [CrossRef]
- Gellings, C.; Parmenter, K. Energy efficiency in fertilizer production and use. In Encyclopedia of Life Support Systems (EOLSS); Eolss Publishers: Oxford, UK, 2004; pp. 123–136. [Google Scholar]
- Gaspar, A.P.; Laboski, C.A.M.; Naeve, S.L.; Conley, S.P. Dry matter and nitrogen uptake, partitioning, and removal across a wide range of soybean seed yield levels. Crop. Sci. 2017, 57, 2170–2182. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.A.; Eberle, C.A.; Thom, M.D.; Archer, D.W.; Forcella, F.; Gesch, R.W.; Wyse, D.L. Economics and agronomics of relay-cropping pennycress and Camelina with Soybean in Minnesota. Agron. J. 2019, 111, 1281–1292. [Google Scholar] [CrossRef]
- Berti, M.; Samarappuli, D.; Johnson, B.L.; Gesch, R.W. Integrating winter camelina into maize and soybean cropping systems. Ind. Crop. Prod. 2017, 107, 595–601. [Google Scholar] [CrossRef]
- Peterson, A.T.; Berti, M.T.; Samarappuli, D. Intersowing cover crops into standing soybean in the US upper midwest. Agronomy 2019, 9, 264. [Google Scholar] [CrossRef] [Green Version]
- Bennett, E.M.; Carpenter, S.R.; Caraco, N.F. Human impact on erodable phosphorus and eutrophication: A global perspective. Bioscience 2001, 51, 227–234. [Google Scholar] [CrossRef]
- Kaspar, T.C.; Singer, J.W. The use of cover crops to manage soil. In Soil Management: Building a Stable Base for Agriculture; Hatfield, J., Sauer, T., Eds.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 2011; pp. 321–337. [Google Scholar]
- Adviento-Borbe, M.A.A.; Haddix, M.L.; Binder, D.L.; Walters, D.T.; Dobermann, A. Soil greenhouse gas fluxes and global warming potential in four high-yielding maize systems. Glob. Chang. Biol. 2007, 13, 1972–1988. [Google Scholar] [CrossRef]
- Russell, A.E.; Cambardella, C.A.; Laird, D.A.; Jaynes, D.B.; Meek, D.W. Nitrogen fertilizer effects on soil carbon balances in Midwestern, U.S. agricultural systems. Ecol. Appl. 2009, 19, 1102–1113. [Google Scholar] [CrossRef] [Green Version]
- Trinsoutrot, I.; Recous, S.; Bentz, B.; Linères, M.; Chèneby, D.; Nicolardot, B. Biochemical quality of crop residues and carbon and nitrogen mineralization kinetics under nonlimiting nitrogen conditions. Soil Sci. Soc. Am. J. 2000, 64, 918–926. [Google Scholar] [CrossRef]
- Kaboneka, S.; Sabbe, W.E.; Mauromoustakos, A. Carbon decomposition kinetics and nitrogen mineralization from corn, soybean, and wheat residues. Commun. Soil Sci. Plant Anal. 1997, 28, 1359–1373. [Google Scholar] [CrossRef]
- Dabney, S.M.; Delgado, J.A.; Reeves, D.W. Using winter cover crops to improve soil and water quality. Commun. Soil Sci. Plant Anal. 2001, 32, 1221–1250. [Google Scholar] [CrossRef]
- Olson, K.; Ebelhar, S.A.; Lang, J.M. Long-term effects of cover crops on crop yields, soil organic carbon stocks and sequestration. Open J. Soil Sci. 2014, 4, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.A.; Wells, M.S.; Anderson, K.; Gesch, R.W.; Forcella, F.; Wyse, D.L. Yield tradeoffs and nitrogen between pennycress, camelina, and soybean in relay- and double-crop systems. Agron. J. 2017, 109, 2128–2135. [Google Scholar] [CrossRef]
- Johnson, G.A.; Kantar, M.B.; Betts, K.J.; Wyse, D.L. Field pennycress production and weed control in a double crop system with soybean in Minnesota. Agron. J. 2015, 107, 532–540. [Google Scholar] [CrossRef] [Green Version]
- Gentry, L.E.; Below, F.E.; David, M.B.; Bergerou, J.A. Role of N2 fixation in the soybean N credit in maize production. Plant Soil 2001, 263, 175–184. [Google Scholar] [CrossRef]
- Green, C.J.; Blackmer, A.M. Residue Decomposition Effects on Nitrogen Availability to Corn following Corn or Soybean. Soil Sci. Soc. Am. J. 1995, 59, 1065–1070. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. F. Crop. Res. 2008, 108, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aulakh, M.S.; Doran, J.W.; Walters, D.T.; Mosier, A.R.; Francis, D.D. Crop residue type and placement effects on denitrification and mineralization. Soil Sci. Soc. Am. J. 1991, 55, 1020–1025. [Google Scholar] [CrossRef] [Green Version]
- Carpenter-Boggs, L.; Pikul, J.L.; Vigil, M.F.; Riedell, W.E. Soil nitrogen mineralization influenced by crop rotation and nitrogen fertilization. Soil Sci. Soc. Am. J. 2000, 64, 2038–2045. [Google Scholar] [CrossRef] [Green Version]
- Hall, S.J.; Russell, A.E.; Moore, A.R. Do corn-soybean rotations enhance decomposition of soil organic matter? Plant Soil 2019, 444, 427–442. [Google Scholar] [CrossRef]
- Fernández, F.G.; Fabrizzi, K.P.; Naeve, S.L. Corn and soybean’s season-long in-situ nitrogen mineralization in drained and undrained soils. Nutr. Cycl. Agroecosyst. 2017, 107, 33–47. [Google Scholar] [CrossRef]
- Ciampitti, I.A.; Salvagiotti, F. New insights into soybean biological nitrogen fixation. Agron. J. 2018, 110, 1185–1196. [Google Scholar] [CrossRef] [Green Version]
- Vanotti, M.B.; Bundy, L.G. Soybean effects on soil nitrogen availability in crop rotations. Agron. J. 1995, 87, 676–680. [Google Scholar] [CrossRef]
- Deng, S.P.; Tabatabai, M.A. Effect of cropping systems on nitrogen mineralization in soils. Biol. Fertil. Soils 2000, 31, 211–218. [Google Scholar] [CrossRef]
- Rochette, P.; Angers, D.A.; Bélanger, G.; Chantigny, M.H.; Prévost, D.; Lévesque, G. Emissions of N2O from alfalfa and soybean crops in eastern Canada. Soil Sci. Soc. Am. J. 2004, 68, 493–506. [Google Scholar] [CrossRef]
- Martinez-Feria, R.A.; Castellano, M.J.; Dietzel, R.N.; Helmers, M.J.; Liebman, M.; Huber, I.; Archontoulis, S.V. Linking crop- and soil-based approaches to evaluate system nitrogen-use efficiency and tradeoffs. Agric. Ecosyst. Environ. 2018, 256, 131–143. [Google Scholar] [CrossRef]
- Wegner, B.R.; Chalise, K.S.; Singh, S.; Lai, L.; Abagandura, G.O.; Kumar, S.; Osborne, S.L.; Lehman, R.M.; Jagadamma, S. Response of soil surface greenhouse gas fluxes to crop residue removal and cover crops under a corn–soybean rotation. J. Environ. Qual. 2018, 47, 1146–1154. [Google Scholar] [CrossRef]
- Kessavalou, A.; Walters, D.T. Winter rye cover crop following soybean under conservation tillage: Residual soil nitrate. Agron. J. 1999, 91, 643–649. [Google Scholar] [CrossRef]
- Regehr, A.; Oelbermann, M.; Videla, C.; Echarte, L. Gross nitrogen mineralization and immobilization in temperate maize-soybean intercrops. Plant Soil 2015, 391, 353–365. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID Code | Sequence (Year 1 → Year 2) | Description |
|---|---|---|
| Continuous Soybean | ||
| S-S | Soybean → Soybean | Conventional sequence. Soybean was sown and harvested in Year 1 and in Year 2. No cover crop. |
| S/Cam-S | Soybean + Camelina → Soybean | Soybean was sown in Year 1 and camelina was interseeded into standing soybean in Year 1 (late summer). Soybean was harvested in fall of Year 1. (Soybean was relayed into standing camelina in the spring of Year 2. Camelina was harvested in early summer in Year 2, while soybean in the fall of the same year. |
| S/Pen-S | Soybean + Pennycress → Soybean | Soybean was sown in Year 1 and pennycress was interseeded into standing soybean in Year 1 (late summer). Soybean was harvested in fall of Year 1. (Soybean was relayed into standing pennycress in the spring of Year 2. Pennycress was harvested in early summer in Year 2, while soybean in the fall of the same year. |
| S/Rye-S | Soybean + Rye → Soybean | Soybean was sown in Year 1 and rye was interseeded into standing soybean in Year 1 (late summer). Soybean was harvested in fall of Year 1. (Rye was terminated before seeding soybean (spring of Year 2). Soybean was sown and harvested in Year 2. |
| Month | Ames | Morris | Prosper | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | LTA † | 2016 | 2017 | LTA | 2016 | 2017 | LTA | ||||||||||
| T | P | T | P | T | P | T | P | T | P | T | P | T | P | T | P | T | P | |
| Jan. | −6.6 | 15 | −4.1 | 47 | −6.3 | 17 | −9.7 | 7 | −9.1 | 13 | −12.0 | 19 | −11.1 | M ‡ | −11.1 | M | −13.5 | M |
| Feb. | −1.6 | 17 | 2.7 | 30 | −4.0 | 22 | −5.2 | 22 | −3.1 | 11 | −9.8 | 18 | −5.6 | M | −5.0 | M | −10.7 | M |
| Mar. | 7.3 | 38 | 3.8 | 79 | 3.3 | 50 | 3.1 | 21 | −0.9 | 12 | −2.6 | 35 | 2.8 | M | −2.2 | M | −3.1 | M |
| Apr. | 11.3 | 104 | 11.6 | 78 | 10.5 | 100 | 6.9 | 52 | 7.5 | 65 | 6.4 | 61 | 5.6 | 43 | 6.7 | 17 | 6.0 | 26 |
| May | 16.6 | 109 | 16.6 | 156 | 16.7 | 124 | 15.3 | 43 | 13.4 | 92 | 13.9 | 77 | 15.0 | 82 | 13.3 | 17 | 13.4 | 72 |
| June | 24.2 | 25 | 23.2 | 44 | 22.0 | 122 | 20.3 | 54 | 20.0 | 101 | 19.2 | 108 | 19.5 | 38 | 18.9 | 88 | 18.9 | 98 |
| July | 23.9 | 149 | 24.9 | 25 | 23.7 | 117 | 21.5 | 184 | 22.1 | 23 | 21.4 | 96 | 21.1 | 88 | 21.1 | 50 | 20.7 | 76 |
| Aug. | 23.1 | 209 | 21.2 | 85 | 22.6 | 122 | 20.7 | 94 | 18.4 | 175 | 20.0 | 83 | 20.6 | 26 | 18.3 | 53 | 19.7 | 54 |
| Sep. | 21.0 | 201 | 20.8 | 46 | 18.7 | 83 | 16.4 | 43 | 16.6 | 105 | 15.2 | 71 | 16.1 | 60 | 15.6 | 152 | 15.0 | 69 |
| Oct. | 14.9 | 15 | 13.0 | 154 | 11.9 | 61 | 9.5 | 87 | 8.5 | 69 | 7.6 | 60 | 8.3 | 49 | 7.8 | 7 | 7.3 | 50 |
| Nov. | 8.2 | 44 | 3.7 | 7 | 3.5 | 46 | 4.3 | 42 | −1.4 | 12 | −1.4 | 24 | 4.4 | M | −3.3 | M | −1.9 | M |
| Dec. | −4.2 | 30 | −3.8 | 4 | −4.1 | 29 | −8.9 | 33 | −9.0 | 7 | −9.1 | 18 | −10.0 | M | −11.1 | M | −9.9 | M |
| SITE CHARACTERISTICS | |||
|---|---|---|---|
| City, state | Ames, Iowa | Morris, Minnesota | Prosper, North Dakota |
| Geographical location | 42.00, −93.73 | 45.67, −95.80 | 46.97, −97.05 |
| Soil texture | Loam | Loam | Silt loam |
| Soil organic carbon (% SOC) | 1.8 | 3.4 | 2.4 |
| Drainage | Good | Good | Poor |
| FIELD MANAGEMENT | |||
| Management type | Conventional fertilization, no residue removal | Reduced fertilization (only cover crop), no residue removal | No fertilization, soybean residue removal in Year 1 |
| Tillage | Conventional tillage: disk plow and spring field cultivation in Year 1, no-till in Year 2 | Conventional tillage: disk and chisel plow and spring field cultivation in Year 1, no-till in Year 2 | Reduced tillage: chisel plow and spring field cultivation in Year 1, no-till in Year 2 |
| Residue management | No soybean and rye residue removal. 100% camelina and pennycress residue removal | No soybean and cover crop residue removal. | 100% soybean residue removal in Year 1, no soybean and cover crop residue removal in Year 2. |
| AGRICULTURAL INPUTS | |||
| Fertilization rate: N-P-K (kg ha−1) | Soybean: 48-123-112 (in Year 1 only) | Soybean: 0-0-0 | Soybean: 0-0-0 |
| Camelina and pennycress: 78-34-34 (in Year 2) | Camelina and pennycress: 78-34-34 (in Year 2) | Camelina and pennycress: 0-0-0 | |
| Winter rye: 0-0-0 | Winter rye: 0-0-0 | Winter rye: 0-0-0 | |
| Fertilizer’s type | Diammonium phosphate, sulfur-coated urea, muriate of potash | Diammonium phosphate, urea, muriate of potash | N/A |
| Herbicide/pesticide (kg a.i. ha−1) | Soybean (in Year 1): glyphosate (2.24), pendimenthalin (1.594) | Soybean (in Year 1): glyphosate (1.1) | Soybean: glyphosate (1.1) |
| Soybean (in Year 2): lactofen (0.56) | Soybean (in Year 2): glyphosate (1.1), lambdacyhalothrin (0.01) | Soybean: N/A | |
| Rye (to terminate): glyphosate (2.24) | Rye (to terminate): glyphosate (2.13) | Rye (to terminate): glyphosate (2.24) | |
| Seeding rate, pure live seed (kg ha−1) | Soybean: 77.0 | Soybean: 77.0 | Soybean: 79.6 |
| Camelina: 11.2 | Camelina: 11.2 | Camelina: 11.2 | |
| Pennycress: 16.8 | Pennycress: 16.8 | Pennycress: 16.8 | |
| Rye: 84.1 | Rye: 84.1 | Rye: 84.1 | |
| MACHINERY | |||
| Machinery use: (a) = fuel (kg ha−1); (b) = electricity (kWh ha−1) | |||
| S-S | (a) 68.8; (b) 0 | (a) 76.0; (b) 0 | (a) 68.0; (b) 0 |
| S/Cam-S | (a) 94.7; (b) 1.5 | (a) 99.7; (b) 2.7 | (a) 91.5; (b) 0.7 |
| S/Pen-S | (a) 95.0; (b) 4.7 | (a) 100.0; (b) 6.4 | (a) 91.7; (b) 3.2 |
| S/Rye-S | (a) 71.7; (b) 0 | (a) 78.0; (b) 0 | (a) 70.0; (b) 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cecchin, A.; Pourhashem, G.; Gesch, R.W.; Mohammed, Y.A.; Patel, S.; Lenssen, A.W.; Berti, M.T. The Environmental Impact of Ecological Intensification in Soybean Cropping Systems in the U.S. Upper Midwest. Sustainability 2021, 13, 1696. https://0-doi-org.brum.beds.ac.uk/10.3390/su13041696
Cecchin A, Pourhashem G, Gesch RW, Mohammed YA, Patel S, Lenssen AW, Berti MT. The Environmental Impact of Ecological Intensification in Soybean Cropping Systems in the U.S. Upper Midwest. Sustainability. 2021; 13(4):1696. https://0-doi-org.brum.beds.ac.uk/10.3390/su13041696
Chicago/Turabian StyleCecchin, Andrea, Ghasideh Pourhashem, Russ W. Gesch, Yesuf A. Mohammed, Swetabh Patel, Andrew W. Lenssen, and Marisol T. Berti. 2021. "The Environmental Impact of Ecological Intensification in Soybean Cropping Systems in the U.S. Upper Midwest" Sustainability 13, no. 4: 1696. https://0-doi-org.brum.beds.ac.uk/10.3390/su13041696