
Biological Indicators of Soil Quality under Different Tillage Systems in Retisol

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Location
2.2. Meteorological Conditions
2.3. Experimental Design and Treatments
2.4. The Evaluation of the Intensity of the Soil Organic Matter Mineralization
2.5. Investigation of Soil Chemical and Physical Properties
2.6. Evaluation of Nutrients Transformation Processes
2.7. The Investigation of Soil Microorganisms
2.8. The Metagenomic Analysis of Soil Microorganisms
2.9. Statistical Analysis
3. Results
3.1. The Alternation of Soil Chemical and Physical Properties in Different Agroecosystems
3.2. The Dynamic of Soil Microorganism Physiological Groups
3.3. Mineralization Intensity of Soil Organic Matter in Different Agroecosystems
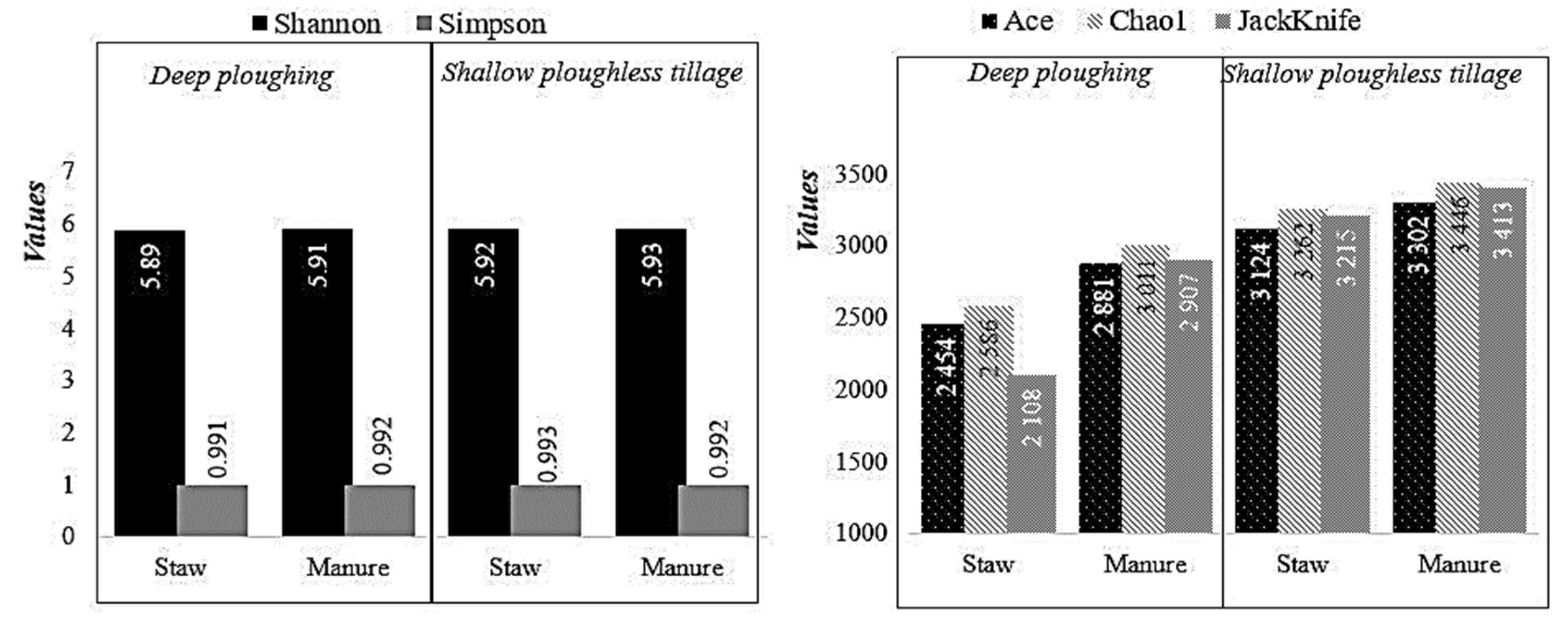
3.4. The Alternation of Soil Bacterial Community in Different Agroecosystems
3.5. Carbon Stock in the Soil under Different Combination of Tillage and Organic Fertilization
4. Discussion
4.1. Soil Properties in Differently Formed Agroecosystems
4.2. Soil Microbiota Activity in Differently Formed Agroecosystems
4.3. Carbon Stock in the Soil under Different Combination of Tillage and Organic Fertilization
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Merrill, S.D.; Liebig, M.A.; Tanaka, D.L.; Krupinsky, J.M.; Hanson, J.D. Comparison of soil quality and productivity at two sites differing in profile structure and topsoil properties. Agric. Ecosyst. Environ. 2013, 179, 53–61. [Google Scholar] [CrossRef]
- de Moraes Sá, J.C.; Lal, R.; Cerri, C.C.; Lorenz, K.; Hungria, M.; de Faccio Carvalho, P.C. Low-carbon agriculture in South America to mitigate global climate change and advance food security. Environ. Int. 2017, 98, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Rahimzadeh, A. Political ecology of climate change: Shifting orchards and a temporary landscape of opportunity. World Dev. Perspect. 2017, 6, 25–31. [Google Scholar] [CrossRef]
- Cassman, K.G. Ecological intensification of maize-based cropping systems. Better Crop. 2017, 101, 4–6. [Google Scholar]
- Juhola, S.; Klein, N.; Käyhkö, J.; Neset, T.-S.S. Climate change transformations in Nordic agriculture? J. Rural. Stud. 2017, 51, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Kadžienė, G. Dirvožemių Savybės Kaitos Integruotas Vertinimas Skirtingose Žemės Dirbimo-Tręšimo Sistemose; Akademija: Kaunas, Lithuania, 2009; p. 119. [Google Scholar]
- Kochiieru, M.; Feiziene, D.; Feiza, V.; Volungevičius, J.; Velykis, A.; Slepetiene, A.; Deveikyte, I.; Seibutis, V. Freez-ing-thawing impact on aggregate stability as affected by land management, soil genesis and soil chemical and physical quality. Soil Tillage Res. 2020, 203, 1047–1055. [Google Scholar] [CrossRef]
- Sollins, P.; Gregg, J.W. Soil organic matter accumulation in relation to changing soil volume, mass, and structure: Concepts and calculations. Geoderma 2017, 301, 60–71. [Google Scholar] [CrossRef]
- Janušauskaitė, D.; Velykis, A.; Satkus, A. Sunkaus priemolio dirvožemio armens ir poarmenio gerinimo priemonių povei-kis mikrobiologiniams procesams. ZEMDIRBYSTE 2008, 2, 45–60. [Google Scholar]
- Xue, P.; Minasny, B.; McBratney, A.B. Land-use affects soil microbial co-occurrence networks and their putative functions. Appl. Soil Ecol. 2022, 169, 104–118. [Google Scholar] [CrossRef]
- Hemkemeyer, M.; Schwalb, S.A.; Heinze, S.; Joergensen, R.G.; Wichern, F. Functions of elements in soil microorganisms. Microbiol. Res. 2021, 126832. [Google Scholar] [CrossRef]
- Adetutu, E.M.; Ball, A.S.; Weber, J.; Aleer, S.; Dandie, C.E.; Juhasz, A.L. Impact of bacterial and fungal processes on 14C-hexadecane mineralization in weathered hydrocarbon contaminated soil. Sci. Total. Environ. 2012, 414, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Stark, C.; Condron, L.; Stewart, A.; Di, H.; O’Callaghan, M. Influence of organic and mineral amendments on microbial soil properties and processes. Appl. Soil Ecol. 2007, 35, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lee, C.G.; Watanabe, T.; Murase, J.; Asakawa, S.; Kimura, M. Identification of microbial communities that assimilate substrate from root cap cells in an aerobic soil using a DNA-SIP approach. Soil Biol. Biochem. 2011, 43, 1928–1935. [Google Scholar] [CrossRef]
- Orgiazzi, A.; Dunbar, M.B.; Panagos, P.; de Groot, G.A.; Lemanceau, P. Soil biodiversity and DNA barcodes: Opportunities and challenges. Soil Biol. Biochem. 2015, 80, 244–250. [Google Scholar] [CrossRef]
- Vieira, A.; Moura, M.; Silva, L. Soil metagenomics in grasslands and forests—A review and bibliometric analysis. Appl. Soil Ecol. 2021, 167, 104047. [Google Scholar] [CrossRef]
- Santos, S.; Nielsen, T.K.; Hansen, L.H.; Winding, A. Comparison of three DNA extraction methods for recovery of soil pro-tist DNA. J. Microbiol. Methods 2015, 115, 13–19. [Google Scholar] [CrossRef]
- Mocali, S.; Benedetti, A. Exploring research frontiers in microbiology: The challenge of metagenomics in soil microbiology. Res. Microbiol. 2010, 161, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Gilbert, J.; Meyer, F. Metagenomics—A guide from sampling to data analysis. Microb. Inform. Exp. 2012, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, S.; Yang, C.-C.; Iwasaki, W. Metagenomics and Bioinformatics in Microbial Ecology: Current Status and Beyond. Microbes Environ. 2016, 31, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Repsiene, R.; Karcauskiene, D. Changes in the chemical properties of acid soil and aggregate stability in the whole profile under long-term management history. Acta Agric. Scand. Sect. B-Plant Soil Sci. 2016, 66, 671–676. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan. Community Ecology Package; 2015. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 20 July 2021).
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9. 2013. Available online: http://purl.oclc.org/estimates (accessed on 20 July 2021).
- Draycott, A.P. Sugar Beet; Blackwell Publishing Ltd.: Oxford, UK, 2006; pp. 114–129. [Google Scholar]
- Volungevicius, J.; Feiza, V.; Amaleviciute-Volunge, K.; Liaudanskiene, I.; Slepetiene, A.; Kuncevicius, A.; Vengalis, R.; Vėlius, G.; Prapiestiene, R.; Poskiene, J. Transformations of different soils under natural and anthropogenized land management. ZEMDIRBYSTE 2019, 106, 3–14. [Google Scholar] [CrossRef]
- Volungevicius, J.; Amalevičiūtė-Volungė, K.; Feiziene, D.; Feiza, V.; Slepetiene, A.; Liaudanskienė, I.; Versuliene, A.; Vaisvalavicius, R. The effects of agrogenic transformation on soil profile morphology, organic carbon and physico-chemical properties in Retisols of Western Lithuania. Arch. Agron. Soil Sci. 2018, 64, 1910–1923. [Google Scholar] [CrossRef]
- Acir, N.; Günal, H.; Çelik, I.; Barut, Z.B.; Budak, M.; Kılıç, S. Effects of long-term conventional and conservational tillage systems on biochemical soil health indicators in the Mediterranean region. Arch. Agron. Soil Sci. 2020, 1–14. [Google Scholar] [CrossRef]
- Salem, H.M.; Meselhy, A.; Elhagarey, M.; Ali, A.M.; Wu, W. Soil erosion control and wheat productivity are improved by a developed ridge-furrow and reservoir tillage systems. Arch. Agron. Soil Sci. 2020, 1–10. [Google Scholar] [CrossRef]
- Samuel, A.D.; Bungau, S.; Tit, D.M.; Melinte Frunzulica, C.E.; Purza, L.; Badea, G.E. Effects of long-term application of organic and mineral fertilizers on soil enzymes. Rev. Chim. 2018, 69, 2608–2612. [Google Scholar] [CrossRef]
- Samuel, A.D.; Bungau, S.; Fodorm, I.K.; Tit, D.M.; Blidar, C.F.; David, A.T. Effects of liming and fertilization on the dehydro-genase and catalase activities. Rev. Chim. 2019, 70, 3464–3468. [Google Scholar] [CrossRef]
- Holub, P.; Klem, K.; Tůma, I.; Vavříková, J.; Surá, K.; Veselá, B.; Urban, O.; Záhora, J. Application of organic carbon affects mineral nitrogen uptake by winter wheat and leaching in subsoil: Proximal sensing as a tool for agronomic practice. Sci. Total Environ. 2020, 717, 137058. [Google Scholar] [CrossRef] [PubMed]
- Kögel-Knabner, I.; Amelung, W. Soil organic matter in major pedogenic soil groups. Geoderma 2021, 384, 114785. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. Organic Amendments: Microbial Community Structure, Activity and Abundance of Agriculturally Relevant Microbes Are Driven by Long-Term Fertilization Strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Xiao, X.; Li, C.; Cheng, K.; Shi, L.; Pan, X.; Li, W.; Wen, L.; Wang, K. Tillage and crop residue incorporation effects on soil bacterial diversity in the double-cropping paddy field of southern China. Arch. Agron. Soil Sci. 2021, 67, 435–446. [Google Scholar] [CrossRef]
- Tripolskaja, L.; Bagdanavičienė, Z.; Romanovskaja, D. Mineralinio azoto ir dirvožemio mikrobinis aktyvumas irstant or-ganinėms trąšoms rudens–žiemos laikotarpiu. Žem. Ūkio Moksl. 2002, 2, 2–12. [Google Scholar]
- Zhao, X.; Zhao, Y.; Xi, B.; Wei, Z.; Wu, J.; Zhao, T. Seasonal population changes in the ammonia-oxidizing bacteria commu-nity structure of Songhua Lake, China. Process Saf. Environ. Prot. 2016, 104, 523–530. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven bio geoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Jirout, J. Nitrous oxide productivity of soil fungi along a gradient of cattle impact. Fungal Ecol. 2015, 17, 155–163. [Google Scholar] [CrossRef]
- Kutateladze, L.; Zakariashvili, N.; Jobava, M.; Burduli, T.; Sadunishvili, T. Microscopic fungi spread in different types of soils in Western Georgia. Ann. Agrar. Sci. 2016, 14, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Bohacz, J. Lignocellulose-degrading enzymes, free-radical transformations during composting of lignocellulosic waste and biothermal phases in small-scale reactors. Sci. Total. Environ. 2017, 580, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Antongiovanni, M.; Sargentini, C. Variability in chemical composition of straws. CIHEAM 1991, 16, 49–53. [Google Scholar]
- Tuomela, M.; Oivanen, P.; Hatakka, A. Degradation of synthetic 14C-lignin by various white-rot fungi in soil. Soil Biol. Biochem. 2002, 34, 1613–1620. [Google Scholar] [CrossRef]
- Tan, B.; Yin, R.; Yang, W.; Zhang, J.; Xu, Z.; Liu, Y.; He, S.; Zhou, W.; Zhang, L.; Li, H.; et al. Soil fauna show different degradation patterns of lignin and cellulose along an elevational gradient. Appl. Soil Ecol. 2020, 155, 103673. [Google Scholar] [CrossRef]
- Torres, I.F.; Bastida, F.; Hernandez, T.; Bombach, P.; Richnow, H.; García, C. The role of lignin and cellulose in the carbon-cycling of degraded soils under semiarid climate and their relation to microbial biomass. Soil Biol. Biochem. 2014, 75, 152–160. [Google Scholar] [CrossRef]
- Mirzavand, J.; Asadi-Rahmani, H.; Moradi-Talebbeigi, R. Biological indicators of soil quality under conventional, reduced, and no-tillage systems. Arch. Agron. Soil Sci. 2020, 1–14. [Google Scholar] [CrossRef]
- Šimon, T.; Javurek, M.; Mikanova, O.; Vach, M. The influence of tillage systems on soil organic matter and soil hydrophobi-city. Soil Tillage Res. 2009, 105, 44–48. [Google Scholar] [CrossRef]
- Bungau, S.; Behl, T.; Aleya, L.; Bourgeade, P.; Aloui-Sossé, B.; Purza, A.L.; Abid, A.; Samuel, A.D. Expatiating the impact of anthropogenic aspects and climatic factors on long-term soil monitoring and management. Environ. Sci. Pollut. Res. 2021, 28, 1–23. [Google Scholar] [CrossRef]
- Kakabouki, I.; Folina, A.; Efthimiadou, A.; Karydogianni, S.; Zisi, C.; Kouneli, V.; Kapsalis, N.C.; Katsenios, N.; Travlos, I. Evaluation of Processing Tomato Pomace after Composting on Soil Properties, Yield, and Quality of Processing Tomato in Greece. Agron. 2021, 11, 88. [Google Scholar] [CrossRef]
- Paustian, K.; Lehmann, J.; Ogle, S.; Reay, D.; Robertson, G.; Smith, P. Climate-smart soils. Nat. Cell Biol. 2016, 532, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhillon, M.R.; Del Corso, B.; Figueiredo, E.; Nambi, W.R. Soil organic carbon, total nitrogen, and soil pH, in a long-term continuous winter wheat (Triticum aestivum L.) experiment. Commun. Soil Sci. Plant Anal. 2018, 49, 803–813. [Google Scholar] [CrossRef]
- Chen, J.; Wei, X. Controlled-Release Fertilizers As a Means to Reduce Nitrogen Leaching and Runoff in Container-Grown Plant Production Nitrogen in Agriculture—Updates; IntechOpen: London, UK, 2018; pp. 33–52. [Google Scholar] [CrossRef] [Green Version]
- Pospisilova, L.; Fasurová, N.; Petrášová, V. Humus content and quality under different soil tillage systems. Soil Water Res. 2010, 5, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Villamil, M.B.; Little, J.; Nafziger, E.D. Corn residue, tillage, and nitrogen rate effects on soil properties. Soil Tillage Res. 2015, 151, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Hábová, M.; Pospíšilová, L.; Hlavinka, P.; Trnka, M.; Barančíková, G.; Tarasovičová, Z.; Takáč, J.; Koco, Š.; Menšík, L.; Nerušil, P. Carbon pool in soil under organic and conventional farming systems. Soil Water Res. 2019, 14, 145–152. [Google Scholar] [CrossRef]
- Segnini, A.; Carvalho, J.L.N.; Bolonhezi, D.; Pereira Milori, D.M.B.; da Silva, W.T.L.; Simões, M.L.; Cantarella, H.; de Maria, I.C.; Neto, L.M. Carbon stock and humification index of organic matter affected by sugarcane straw and soil man-agement. Sci. Agric. 2013, 70, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Litvinovich, A.; Pavlova, O.; Lavrishchev, A.; Bure, V.; Saljnikov, E. Empirical models of transformations of humic acids and humin in Umbric Albeluvisol Abruptic as influenced by liming. Zemdirb.-Agric. 2017, 104, 115–122. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Soil Tillage | Organic Fertilizers | |||||
|---|---|---|---|---|---|---|
| Deep Ploughing | Shallow Ploughless Tillage | Plant Residue | Straw | Green Manure 1st Cut | Manure 40 t ha−1 | |
| pHKCl | 5.6 | 5.2 * | 5.5 | 5.4 | 5.6 | 5.6 |
| K2O, mg kg−1 | 289 | 228 * | 260 | 271 | 265 | 283 |
| P2O5, mg kg−1 | 204 | 140 ** | 176 | 188 | 181 | 202 ** |
| Ntotal, % | 0.17 | 0.16 ** | 0.17 | 0.17 | 0.16 * | 0.16 * |
| Corg. % | 1.59 | 1.46 ** | 1.56 | 1.58 | 1.54 | 1.53 |
| Treatments | Primary Soil Tillage | Organic Fertilizers | |||||
|---|---|---|---|---|---|---|---|
| Deep Ploughing | Shallow Ploughless Tillage | Shallow Ploughless Tillage + Deep Loosening | Plant Residue | Straw | Green Manure 1st Cut | Manure 40 t ha−1 | |
| pH | 5.51 ± 0.2 | 5.25 ± 0.3 | 5.37 ± 0.2 | 5.34 ± 0.3 | 5.43 ± 0.2 | 5.34 ± 0.2 | 5.39 ± 0.2 |
| K2O, mg kg−1 | 284.25 ± 82 | 327.88 ± 116 | 273.58 ± 59 | 284.17 ± 110 | 294.78 ± 59 | 304.78 ± 103 | 300.22 ± 90 |
| P2O5, mg kg−1 | 189.96 ± 36 | 174.17 ± 55 | 136.92 ± 36 | 171.50 ± 44 | 169.50 ± 51 | 153.17 ± 43 | 175.22 ± 56 |
| Ntotal, % | 0.13 ± 0.01 | 0.14 ± 0.02 | 0.14 ± 0.02 | 0.13 ± 0.02 | 0.13 ± 0.02 | 0.14 ± 0.02 | 0.14 ± 0.01 |
| Corg., % | 1.59 ± 0.2 | 1.74 ± 0.2 | 1.47 ± 0.2 | 1.66 ± 0.2 | 1.53 ± 0.3 | 1.64 ± 0.2 | 1.59 ± 0.3 |
| Moisture, % | 18.85 ± 0.9 | 20.49 ± 0.9 | 19.11 ± 1.3 | 18.83 ± 0.7 | 20.06 ± 1.3 | 19.79 ± 1.3 | 19.12 ± 1.1 |
| Porosity, % | 47.88 ± 6.6 | 49.01 ± 3.1 | 49.57 ± 3.5 | 47.51 ± 7.7 | 47.97 ± 1.9 | 51.19 ± 3.0 | 48.79 ± 3.3 |
| Bulk density, Mg m−3 | 1.37 ± 0.07 | 1.35 ± 0.09 | 1.34 ± 0.09 | 1.36 ± 0.09 | 1.39 ± 0.06 | 1.30 ± 0.08 | 1.37 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilkiene, M.; Mockeviciene, I.; Karcauskiene, D.; Suproniene, S.; Doyeni, M.O.; Ambrazaitiene, D. Biological Indicators of Soil Quality under Different Tillage Systems in Retisol. Sustainability 2021, 13, 9624. https://0-doi-org.brum.beds.ac.uk/10.3390/su13179624
Vilkiene M, Mockeviciene I, Karcauskiene D, Suproniene S, Doyeni MO, Ambrazaitiene D. Biological Indicators of Soil Quality under Different Tillage Systems in Retisol. Sustainability. 2021; 13(17):9624. https://0-doi-org.brum.beds.ac.uk/10.3390/su13179624
Chicago/Turabian StyleVilkiene, Monika, Ieva Mockeviciene, Danute Karcauskiene, Skaidre Suproniene, Modupe Olufemi Doyeni, and Dalia Ambrazaitiene. 2021. "Biological Indicators of Soil Quality under Different Tillage Systems in Retisol" Sustainability 13, no. 17: 9624. https://0-doi-org.brum.beds.ac.uk/10.3390/su13179624