


The Activity and Gene Expression of Enzymes in Mycelia of Pleurotus Eryngii under Cadmium Stress
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Treatments
2.2. Liquid Fermentation Cultures of P. eryngii under Cadmium (Cd) Stress
2.3. Determination of MDA and H2O2 Contents
2.4. Determination of Antioxidant Enzyme Activity
2.5. Calculation of the Amounts of MDA, H2O2, and Enzymes Produced in Mycelia
2.6. Measurement of Free Sugar Concentrations
2.7. Standard Curve Plots
2.8. Detection of the Expression of Antioxidant Enzymes and Sugar Metabolism Enzymes by Fluorescence Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.8.1. RNA Extraction and cDNA Preparation
2.8.2. Primer Design and Fluorescence Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
3. Results
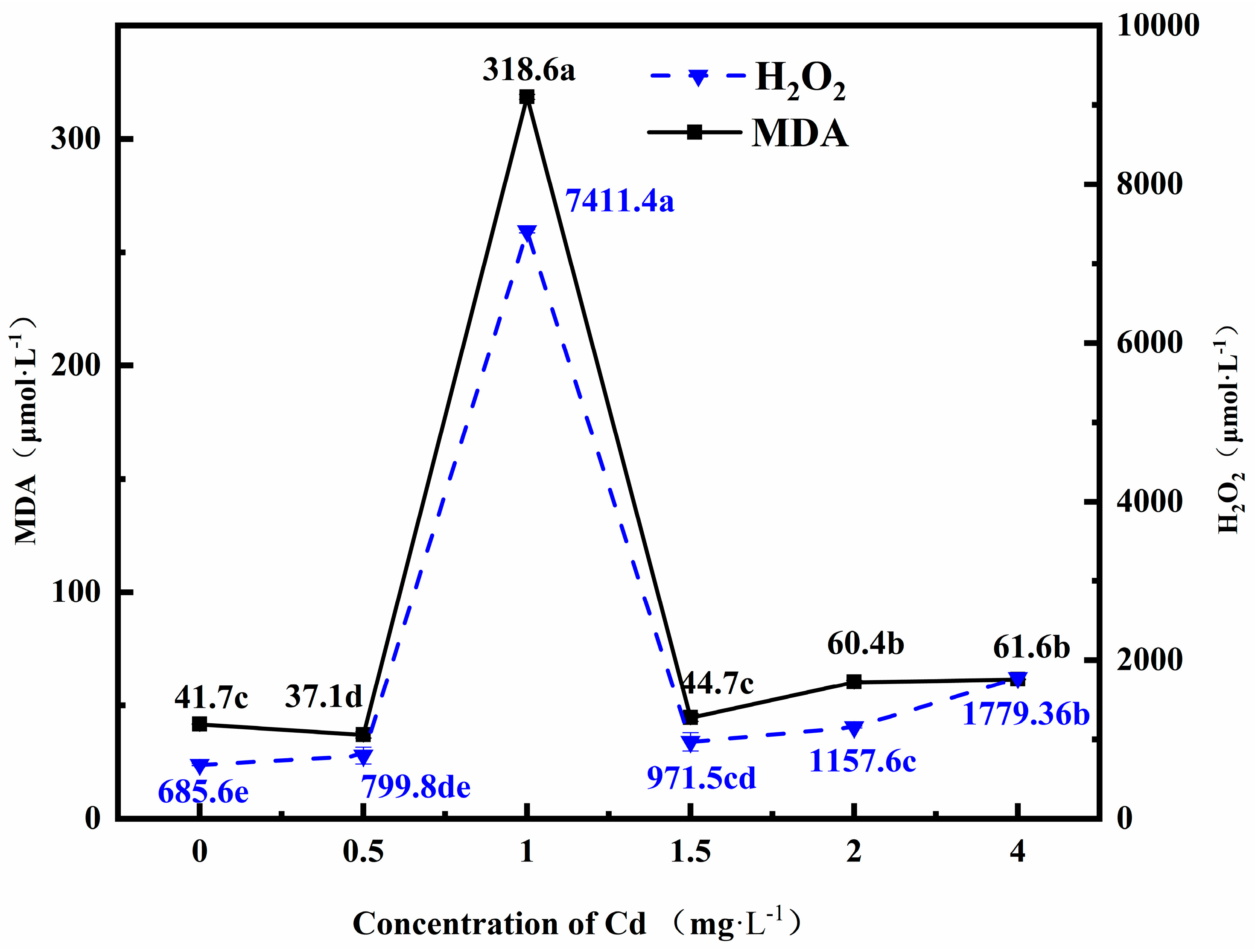
3.1. Changes in the Cell Membrane Damage Index in Fungi Grown under Cadmium (Cd) Stress
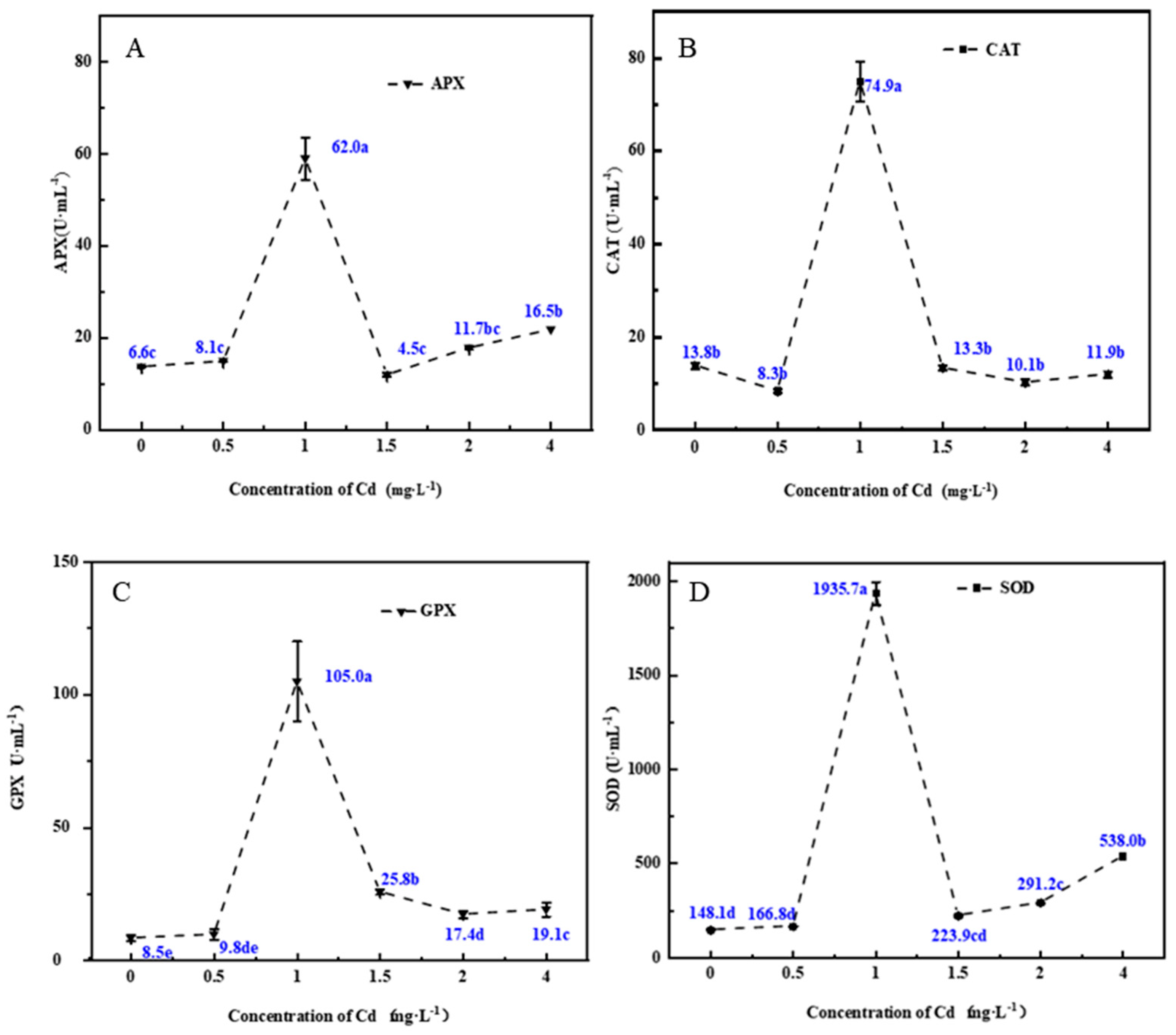
3.2. Changes in Antioxidant Enzyme Activities in the Fermentation Mycelia under Cd Stress
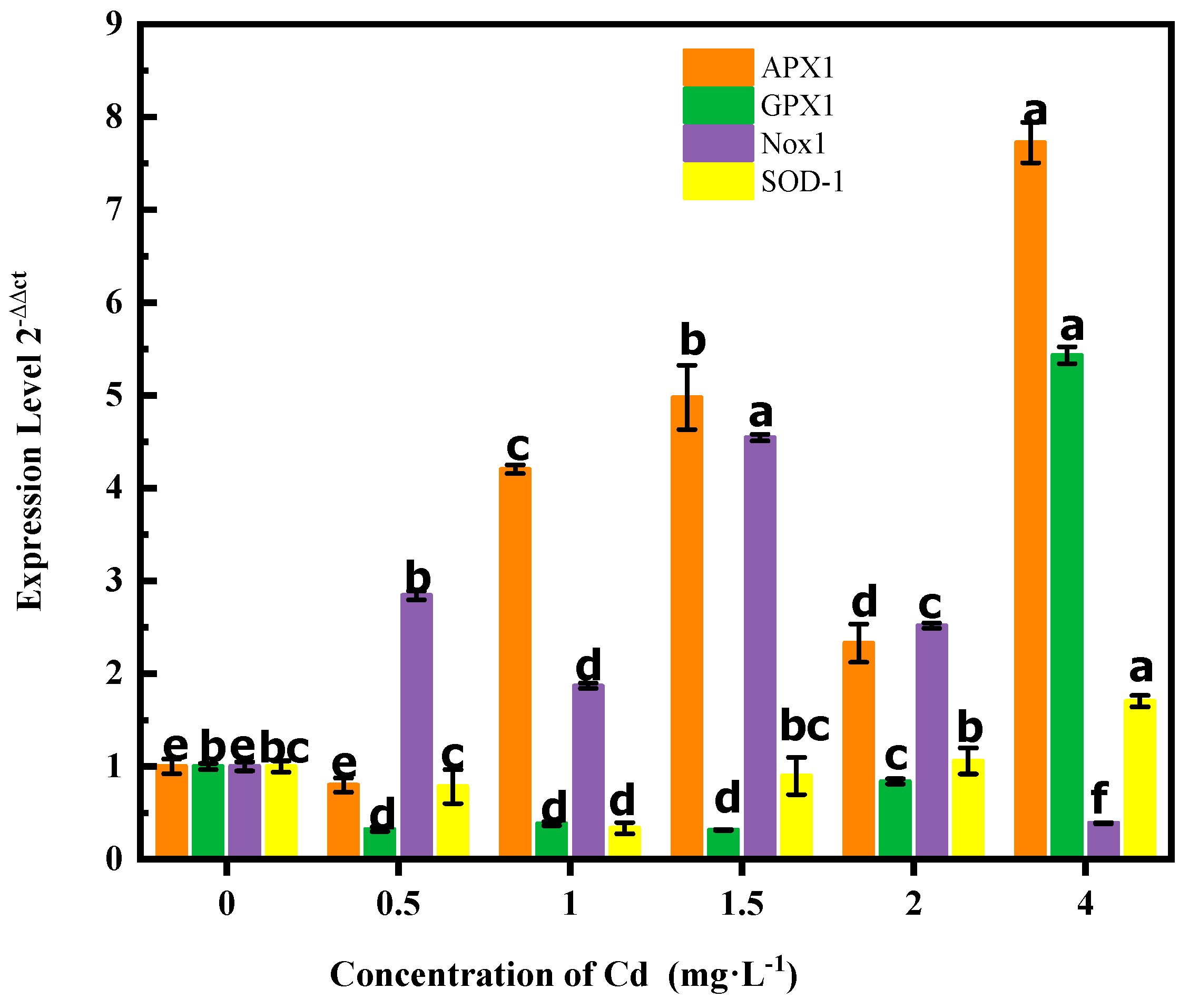
3.3. Changes in Antioxidant Enzyme Gene Expression during Cadmium (Cd) Stress
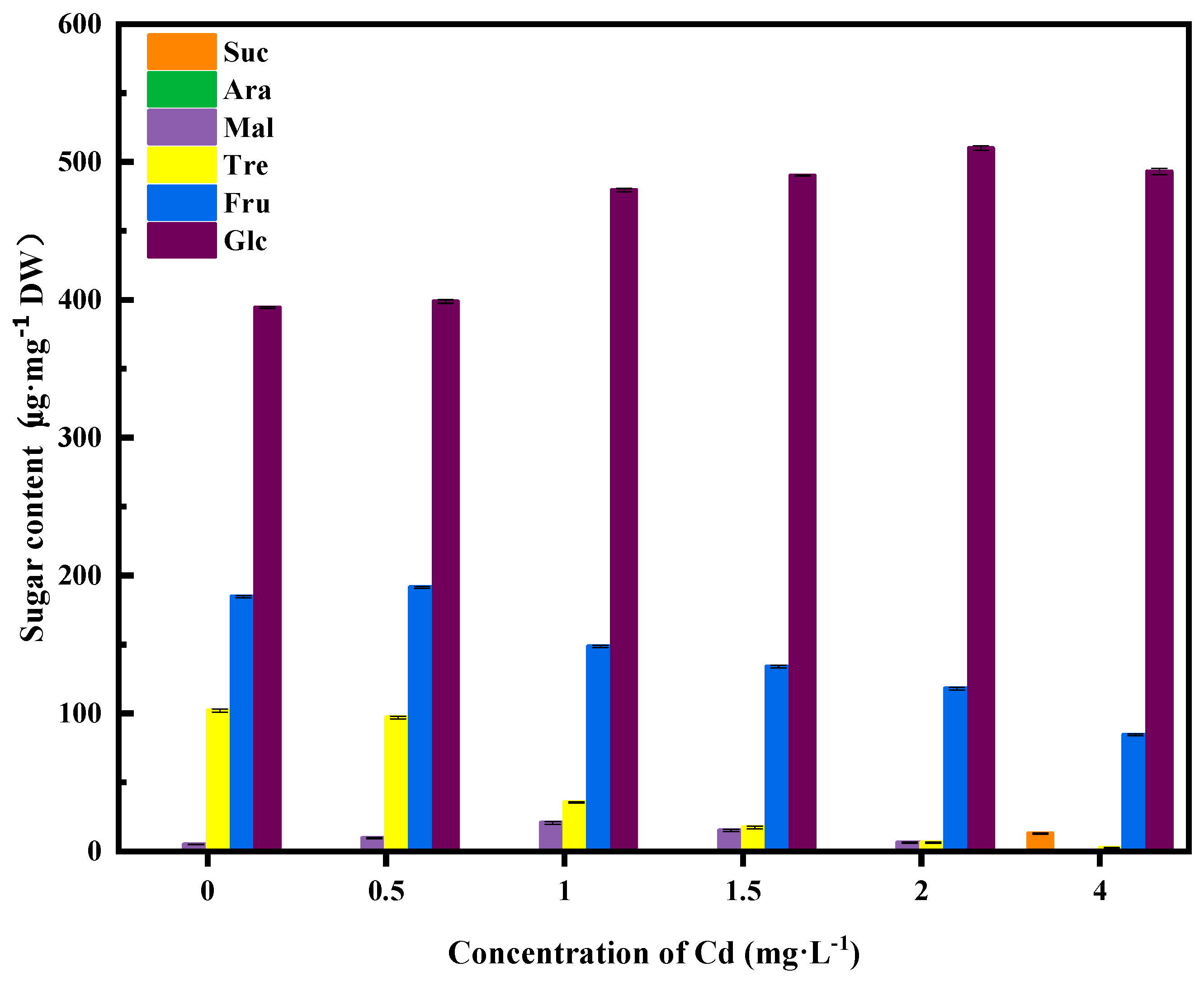
3.4. Variation in Levels of Soluble Sugars in the Fermentation Broth during Cd Stress
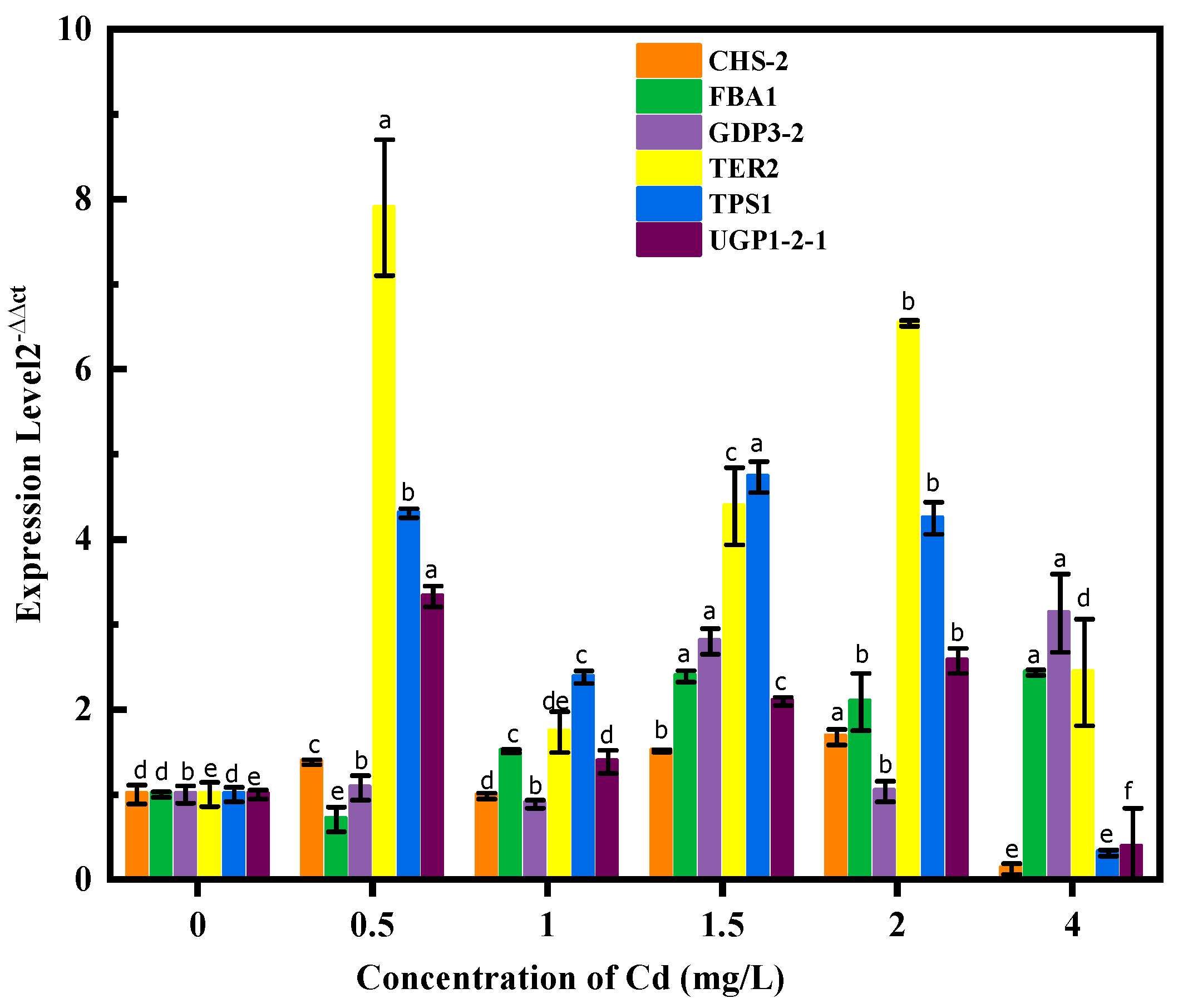
3.5. The Gene Expression of Soluble Sugar Metabolism–Related Enzymes during Cd Stress
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X.Y.; Wu, C.Y.; Zhang, G.C. Study on heavy metal pollution and control of vegetables. J. Anhui Agric. Sci. 2019, 47, 10–12. (In Chinese) [Google Scholar]
- Wang, M.; Zou, J.; Duan, X.; Jiang, W.; Liu, D. Cadmium accumulation and its effects on metal uptake in maize. Bioresour. Technol. 2007, 98, 82–88. (In Chinese) [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.W.; Zhang, J.; Du, J. Research progress of treatment of cadmium and chromium wastewater by microbiological technology. Biot. Resour. 2020, 42, 313–321. (In Chinese) [Google Scholar]
- Chatterjee, S.; Chatterjee, N.C.; Dutta, S.J. Bioreduction of chromium (Ⅵ) to chromium (Ⅲ) by a novel yeast strain Rhodotorula mucilaginosa (MTCC 9315). Afr. J. Biotechnol. 2012, 11, 14920–14929. [Google Scholar]
- Bahafid, W.; Joutey, N.T.; Sayel, H.; Mohamed, I.; Naïma, E. Chromium adsorption by three yeast strains isolated from sediments in Morocco. Geomicrobiol. J. 2013, 30, 422–429. [Google Scholar] [CrossRef]
- Fernández, P.M.; Cabral, M.E.; Delgado, O.D. Textile-dye polluted waters as a source for selecting chromate-reducing yeasts through Cr(Ⅵ)- enriched mi-crocosms. Int. Biodeterior. Biodegrad. 2013, 79, 28–35. [Google Scholar] [CrossRef]
- Huang, X.H.; Wang, C.H.; Xu, N. Tolerance and accumulation of four heavy metals in mycelium of Flammulina velutipes and Pleurotus eryngii. North. Hortic. 2019, 3, 143–150. (In Chinese) [Google Scholar]
- Zhang, Y.R.; Zhao, Y.; Song, P.P. Studies on growth and antioxidant system of Stropharia rugosoannulata mycelia by cadmium stress. Mol. Plant Breed. 2020, 3, 61–62. (In Chinese) [Google Scholar]
- Yu, H.; Zhang, J.Y.; Hu, Y.Q. Study on physiological response of Flammulina velutipes under stress of Mn2+ and Pb2+. North. Hortic. 2020, 18, 132–136. (In Chinese) [Google Scholar]
- Zhao, X.; Song, X.X.; Li, Y.P.; Li, Y.; Yu, C.; Zhao, Y.; Gong, M.; Shen, X.; Chen, M. Gene expression related to trehalose metabolism and its effect on Volvariella volvacea under low temperature stress. Sci. Rep. 2018, 8, 11011. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, X.Y.; Tang, C.H. Change of polyol and trehalose content and expression of their related metabolism enzymes in Ganoderma lingzhi SH during fruiting body growth. Mycosystema 2018, 37, 1090–1099. (In Chinese) [Google Scholar]
- Bai, Y.; Wang, J.; Han, J.; Han, J.; Xie, X.L.; Ji, C.G.; Lei, J.Y.; Yin, J.; Chen, L.; Wang, C.K.; et al. BCL2L10 inhibits growth and metastasis of hepatocellular carcinoma both in vitro and in vivo. Mol. Carcinogen. 2016, 56, 1137–1149. [Google Scholar] [CrossRef]
- Ding, A.Q.; Zheng, P.; Zhang, M.; Zhang, Q.Q. Impacts of electron donor and acceptor on the performance of electrotrophic denitrification. Environ. Sci. Pollut. Res. 2017, 24, 19693–19702. [Google Scholar] [CrossRef]
- Nie, J. Study on the Remediation Potential and Tolerance Mechanism of the Cadmium Contaminated Soil Treated by BH; Hunan University: Changsha, China, 2016. (In Chinese) [Google Scholar]
- Roxas, V.P.; Lodhi, S.A.; Garrett, D.K.; Allen, R.D. Stress tolerance in transgenic tobacco seedlings that overexpress glutathione S-transferase/glutathione peroxidase. Plant Cell Physiol. 2000, 41, 1229–1234. [Google Scholar] [CrossRef]
- Liang, C.; Wang, C.; Yang, X.F. Salt-tolerant Physiological Characters of Wheat Variety Dekang 961. Acta Bot. Boreali-Occident. Sin. 2019, 26, 2075–2082. (In Chinese) [Google Scholar]
- Li, M.; Zhao, Z.T.; Xu, J.R. Evolutionary genomic analysis of the chitin synthase gene family in fungi. In Proceedings of the 2014 Annual Conference of the Chinese Plant Pathology Society, Shenyang, China, 30 July 2014; p. 198. (In Chinese). [Google Scholar]
- Hou, Q.Q.; Liu, S.J.; Liu, C.B. Study on the catalytic mechanism of a novel bifunctional enzyme, fructose acetalase/phosphatase. In Proceedings of the 13th National Chemical Kinetics Conference of the Chinese Chemical Society, Wuhu, China, 23–28 August 2013; pp. 139–140. (In Chinese). [Google Scholar]
- Wu, Y.; Wu, M.; He, G.; Zhang, X.; Li, W.; Gao, Y.; Li, Z.; Wang, Z.; Zhang, C. Glyceraldehyde-3-phosphate dehydrogen-ase:a universal internal control for western blots in prokaryotic andeukaryotic cells. Anal. Biochem. 2012, 423, 15–22. [Google Scholar] [CrossRef]
- Lu, Q.; Mi, X.J.; Cui, J. Research Advances on the Mechanism of Glyceraldehydes-3-phosphate Dehydrogenase in Plant. Biotechnol. Bull. 2013, 8, 28. (In Chinese) [Google Scholar]
- Sandhoff, K.; Echten, G.V.; Schröder, M.; Schnabel, D.; Suzuki, K. Metabolism of glycolipids: The role of glycolipid-binding proteins in the function and pathobiochemistry of lysosomes. Biochem. Soc. Trans. 1992, 20, 695–699. [Google Scholar] [CrossRef]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotechnol. 2015, 16, 123–132. [Google Scholar] [CrossRef]
- Al-Bader, N.; Vanier, G.; Liu, H.; Gravelat, F.N.; Urb, M.; Hoareau, C. Role of trehalose biosynthesis in Aspergillus fumigatus development, stress response, and virulence. Infect. Immun. 2010, 78, 3007–3018. [Google Scholar] [CrossRef] [Green Version]
- Nazir, F.; Fariduddin, Q.Z.; Khan, T.A. Hydrogen peroxide as a signalling molecule in plants and its crosstalk with other plant growth regulators under heavy metal stress. Chemosphere 2020, 252, 126486–126505. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, T.; Li, Z.Q.; Jia, R.X.; Luo, D.J.; Tang, M.Q.; Huang, Z. Transcriptome analysis revealed key genes and pathways related to cadmium-stress tolerance in Kenaf (Hibiscus cannabinus L.). Ind. Crop. Prod. 2020, 158, 112970–112982. [Google Scholar] [CrossRef]
- Chen, Q.; Lu, X.; Guo, X.; Pan, Y.; Yu, B.; Tang, Z.; Guo, Q. Differential responses to Cd stress induced by exogenous application of Cu, Zn or Ca in the medicinal plant Catharanthus roseus. Ecotoxicol. Environ. Saf. 2018, 157, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, M.; Mansour, G.; Khalil, K. Physiological and antioxidative responses of medicinal plants exposed to heavy metals stress. Plant Gene 2017, 11, 247–254. [Google Scholar]
- Wang, M.Y.; Fu, Y.Q.; Yin, T. Study on growth and metabolites accumulation of Ganoderma lucidum mycelium by cadmium stress. Chin. Tradit. Herb. Drugs 2019, 50, 3444–3452. (In Chinese) [Google Scholar]
- Alaraidh, I.A.; Alsahli, A.A.; Abdel Razik, E.S. Alteration of antioxidant gene expression in response to heavy metal stress in Trigonella foenum-graecum L. S. Afr. J. Bot. 2018, 115, 90–93. [Google Scholar] [CrossRef]
- Gao, Y.Y.; Wang, Y.; Qian, J.; Si, W.S.; Tan, Q.; Xu, J.Y.; Zhao, Y.C. Melatonin enhances the cadmium tolerance of mushrooms through antioxidant-related metabolites and enzymes. Food Chem. 2020, 330, 127263–127293. [Google Scholar] [CrossRef]
- Du, Y.L.; Zhao, Q.; Chen, L.R.; Yao, X.D.; Zhang, W.; Zhang, B.; Xie, F.T. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 2020, 146, 1–12. [Google Scholar] [CrossRef]
- Liu, X.M.; Wu, X.L.; Gao, W.; Qu, J.B.; Chen, Q.; Huang, C.Y.; Zhang, J.X. Protective roles of trehalose in Pleurotus pulmonarius during heat stress response. J. Integr. Agric. 2019, 18, 428–437. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | ||||||
|---|---|---|---|---|---|---|
| Element | CK | 1 | 2 | 3 | 4 | 5 |
| Cd | 0 | 0.5 | 1 | 1.5 | 2 | 4 |
| Reagent | Volume (μL) | Final Concentration |
|---|---|---|
| 5 × PrimeScript RT Master Mix (Perfect Real Time) | 2 μL | 1× |
| Total RNA | - | 500 ng |
| RNase Free dH2O | up to 10 μL |
| Reagent | Volume (μL) |
|---|---|
| TB Green Premix Ex Taq II (Tli RNaseH Plus) (2×) | 5 |
| Primer F | 0.4 |
| Primer R | 0.4 |
| ROX Reference Dye (50×) | 0.2 |
| DNA Sample | 1 |
| ddH2O | 3 |
| Total Volume | 10 |
| Name | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Trehalase 2 (TRE2) | CTACGTACTACAGCGGCGAG | CCTTGGACGAAGATGCCAGT |
| Trehalose-6-phosphate synthase1 (TPS1) | TGATGACGTAGCAGATCGCC | AAGTTCATCTCGCCTGGGTG |
| Fructose-diphosphate aldolase (FBA1-1) | CACTCAGCCGGAGGACATTT | TTTGCTTGCTCGTGAACTGC |
| Chitin synthase (CHS-2) | GCAGCAATGAGAGCTCCAAC | GCCGTGTATCTCCTCGTCTG |
| UTP—glucose-1-phosphate uridyl transferase (UGP1-2-1) | TCAGTCGTCTTGTCAGCACC | CCAAGTAGCGGGTGAAGAGG |
| Glyceraldehyde-3-phosphate dehydrogenase (GDP3-2) | TCGCCGTTATCAAGGAAGCC | AAGTCGGTGGAGACTACGGA |
| Catalase1 (CAT1) | TTAATCACCATCCGCCAGCA | CCAACTCCTCGGGCCATATC |
| Ascorbate peroxidase1 (Apx1) | TGGTGTTTGCGCAATTCAGG | GTCAGCACCTTGGGTAGCAT |
| Glutathione peroxidase1 (GPX1) | CGAGAAGGGCCTCGAGATTC | GGCCATGAGTGGGAAAGTCA |
| NADPH oxidase isoform 1 (Nox1) | AGTCGCTGTCCTCATTGGTG | CTCCTGCAGCAGACTTTGGA |
| Superoxide dismutase (SOD-1) | ACGAATTCAGACTCATTCCCGT | AATATCGAGCAGTTGCGGGT |
| ACTIN1-1 | CCCCTGAGCGAAAGTACTCC | GCCGGACTCGTCGTATTCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.-H.; Xu, N.; Feng, L.-G.; Lai, D.-N.; Wu, F.; Xu, D.; Guo, X. The Activity and Gene Expression of Enzymes in Mycelia of Pleurotus Eryngii under Cadmium Stress. Sustainability 2022, 14, 4125. https://0-doi-org.brum.beds.ac.uk/10.3390/su14074125
Huang X-H, Xu N, Feng L-G, Lai D-N, Wu F, Xu D, Guo X. The Activity and Gene Expression of Enzymes in Mycelia of Pleurotus Eryngii under Cadmium Stress. Sustainability. 2022; 14(7):4125. https://0-doi-org.brum.beds.ac.uk/10.3390/su14074125
Chicago/Turabian StyleHuang, Xiao-Hui, Ning Xu, Li-Guo Feng, Deng-Ni Lai, Fang Wu, Dong Xu, and Xin Guo. 2022. "The Activity and Gene Expression of Enzymes in Mycelia of Pleurotus Eryngii under Cadmium Stress" Sustainability 14, no. 7: 4125. https://0-doi-org.brum.beds.ac.uk/10.3390/su14074125