
Pertinent Water-Saving Management Strategies for Sustainable Turfgrass in the Desert U.S. Southwest
 , ,
, ,
Abstract
:1. Introduction
2. Turfgrass Water Requirements
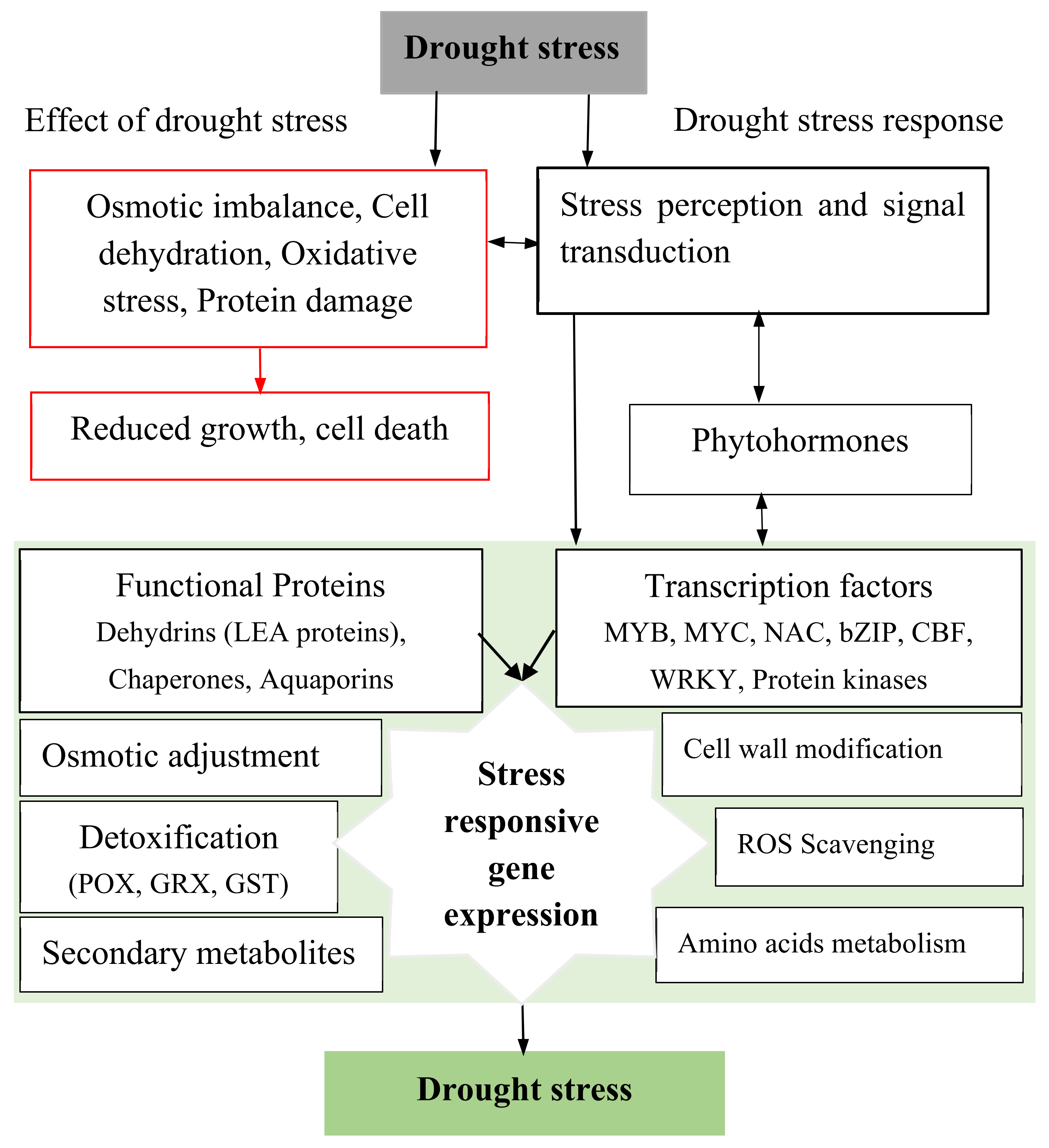
3. Drought Stress Effect on Turfgrass
4. Genetic Variation for Drought Tolerance in Turfgrass
5. Breeding Turfgrass for Drought Tolerance
6. Genetic Manipulation for Enhanced Drought Stress Tolerance
7. Cultural Practices Leveraging Turfgrass Drought Tolerance
7.1. Mowing Height and Frequency
7.2. Turfgrass Nutrition
7.3. Winter Overseeding
7.4. Irrigation Application Methods and Scheduling
7.5. Exogenous Application of Hormones and Osmoprotectants
7.6. Use of Native Grasses for Water Saving
7.7. Use of Recycled Water
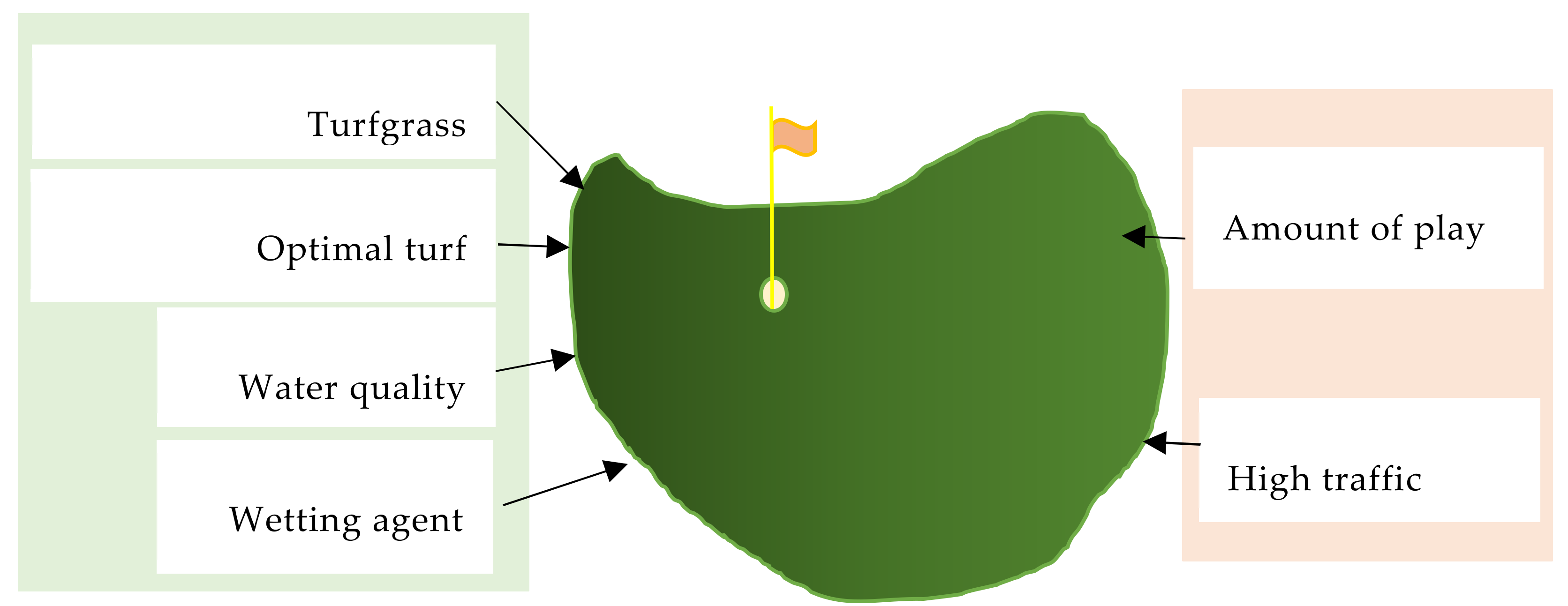
8. Turfgrass Aesthetic Quality and Sustainability
9. Summary and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Beard, J.B. Turfgrass: Science and Culture; Prentice-Hall Inc.: Englewood Cliffs, NJ, USA, 1973. [Google Scholar]
- Beard, J.B.; Green, R.L. The Role of turfgrasses in environmental protection and their benefits to humans. J. Environ. Qual. 1994, 23, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Gross, C.M.; Angle, J.S.; Welterlen, M.S. Nutrient and sediment losses from turfgrass. J. Environ. Qual. 1990, 19, 663–668. [Google Scholar] [CrossRef]
- Beard, J.B.; Johnson, E.C. The comparative heat dissipation from three typical urban surfaces: Asphalt, concrete, and bermudagrass turf. Tex. Turfgrass Res. 1985, PR-4329, 125–133. [Google Scholar]
- Bandaranayake, W.; Qian, Y.L.; Parton, W.J.; Ojima, D.S.; Follett, R.F. Estimation of soil organic carbon changes in turfgrass systems using the CENTURY model. Agron. J. 2003, 95, 558–563. [Google Scholar] [CrossRef]
- Braun, R.C.; Bremer, D.J. Carbon sequestration in zoysiagrass turf under different irrigation and fertilization management megimes. Agrosyst. Geosci. Environ. 2019, 2, 180060. [Google Scholar] [CrossRef]
- Stier, J.C.; Steinke, K.; Ervin, E.H.; Higginson, F.R.; McMaugh, P.E. Turfgrass benefits and Issues. In Turfgrass: Biology, Use, and Management; Stier, J.C., Horgan, B.P., Bonos, S.A., Eds.; American Society of Agronomy: Madison, WI, USA, 2013; Volume 56, pp. 105–145. [Google Scholar] [CrossRef] [Green Version]
- Duval, D.; Kerna, A.; Frisvold, G.; Umeda, K.; Li, R. Contribution of the Golf Industry to the Arizona Economy in 2014; The University of Arizona: Tucson, AZ, USA, 2016. [Google Scholar]
- Wheeler, K.; Nauright, J. A global perspective on the environmental impact of golf. Sport Soc. 2006, 9, 427–443. [Google Scholar] [CrossRef]
- Smolianov, P.; Morrissette, J.N.; Ridpath, B.D.; Boucher, C.; Dion, S.; Schoen, C.; Stone, N.; O’Connor, C.; Miles, K.B. Comparing the practices of US golf against a global model for integrated development of mass and high performance sport. Int. J. Golf Sci. 2020, 9, 1–24. [Google Scholar]
- Wang, H.; Marshall, C.W.; Cheng, M.; Xu, H.; Li, H.; Yang, X.; Zheng, T. Changes in land use driven by urbanization impact nitrogen cycling and the microbial community composition in soils. Sci. Rep. 2017, 7, 44049. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Karoly, D.; Vicarelli, M.; Neofotis, P.; Wu, Q.; Casassa, G.; Menzel, A.; Root, T.L.; Estrella, N.; Seguin, B.; et al. Attributing physical and biological impacts to anthropogenic climate change. Nature 2008, 453, 353–357. [Google Scholar] [CrossRef]
- Oliveira, A.; Lopes, A.; Correia, E.; Niza, S.; Soares, A. An urban climate-based empirical model to predict present and future patterns of the urban thermal signal. Sci. Total Environ. 2021, 790, 147710. [Google Scholar] [CrossRef]
- Suckling, E. Seasonal-to-decadal climate forecasting. In Weather & Climate Services for the Energy Industry; Troccoli, A., Ed.; Palgrave Macmillan: Cham, Switzerland, 2018; pp. 123–137. [Google Scholar] [CrossRef]
- Glennon, R.; The Conversation. As Colorado River Basin States Confront Water Shortages, It’s Time to Focus on Reducing Demand. Available online: https://theconversation.com/as-colorado-river-basin-states-confront-water-shortages-its-time-to-focus-on-reducing-demand-165646 (accessed on 28 June 2022).
- Schiavon, M.; Leinauer, B.; Serena, M.; Maier, B.; Sallenave, R. Plant growth regulator and soil surfactants’ effects on saline and deficit irrigated warm-season grasses: I. Turf quality and soil moisture. Crop Sci. 2014, 54, 2815–2826. [Google Scholar] [CrossRef] [Green Version]
- Colmer, T.; Barton, L. A review of warm-season turfgrass evapotranspiration, responses to deficit irrigation, and drought resistance. Crop Sci. 2017, 57, S-98–S-110. [Google Scholar] [CrossRef]
- Huang, B. Turfgrass water requirements and factors affecting water usage. In Water Quality Quantity Issues Turfgrasses Urban Landscapes; Council for Agricultural Science and Technology: Ames, IA, USA, 2008; Volume 27, pp. 193–203. [Google Scholar]
- Haghverdi, A.; Reiter, M.; Sapkota, A.; Singh, A. Hybrid bermudagrass and tall fescue turfgrass irrigation in central California: I. Assessment of visual quality, soil moisture and performance of an ET-Based smart controller. Agronomy 2021, 11, 1666. [Google Scholar] [CrossRef]
- Yue, C.; Cui, M.; Kong, X.; Watkins, E.; Barnes, M. Landscape irrigation and water conservation in urban areas: An analysis of information-based strategies. Horttechnology 2022, 32, 213–225. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat Stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.-B.; Chu, L.-Y.; Jaleel, C.A.; Zhao, C.-X. Water-deficit stress-induced anatomical changes in higher plants. Comptes Rendus Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Fan, J.B.; Zhang, W.H.; Amombo, E.; Hu, L.X.; Kjorven, J.O.; Chen, L. Mechanisms of Environmental Stress Tolerance in Turfgrass. Agronomy 2020, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.S. Breeding for abiotic stress resistance in sorghum. In Breeding Sorghum for Diverse End Uses; Aruna, C., Visarada, K.B.R.S., Bhat, B.V., Tonapi, V.A., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 325–340. [Google Scholar] [CrossRef]
- Senthilkumar, M.; Amaresan, N.; Sankaranarayanan, A. Determination of physiological traits telated to abiotic stresses. In Plant-Microbe Interactions; Springer: New York, NY, USA, 2021; pp. 157–163. [Google Scholar] [CrossRef]
- Vesala, T.; Sevanto, S.; Grönholm, T.; Salmon, Y.; Nikinmaa, E.; Hari, P.; Hölttä, T. Effect of leaf water potential on internal humidity and CO2 dissolution: Reverse transpiration and improved water use efficiency under negative pressure. Front. Plant Sci. 2017, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, F.; Kuromori, T.; Urano, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Drought stress responses and resistance in plants: From cellular responses to long-distance intercellular communication. Front. Plant Sci. 2020, 11, 556972. [Google Scholar] [CrossRef] [PubMed]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Bales, R.C.; Liverman, D.M. Climate patterns and trends in the Southwest. In Climate Variability and Change in the Southwest: Impacts, Information Needs, and Issues for Policymaking; Merideth, R., Liverman, D., Bales, R., Patterson, M., Eds.; The University of Arizona: Tucson, AZ, USA, 1997; p. 87. [Google Scholar]
- Pendergrass, A.G.; Knutti, R.; Lehner, F.; Deser, C.; Sanderson, B.M. Precipitation variability increases in a warmer climate. Sci. Rep. 2017, 7, 17966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Lambrides, C.J.; Fukai, S. Drought resistance of bermudagrass (Cynodon spp.) ecotypes collected from different climatic zones. Environ. Exp. Bot. 2013, 85, 22–29. [Google Scholar] [CrossRef]
- Aydinsakir, K.; Buyuktas, D.; Bastug, R.; Yilmaz, S. Evapotranspiration and quality characteristics of some bermudagrass turf cultivars under deficit irrigation. Grassl. Sci. 2016, 62, 224–232. [Google Scholar] [CrossRef]
- Wang, J.; Burgess, P.; Bonos, S.A.; Meyer, W.A.; Huang, B. Differential physiological responses and genetic variations in fine fescue species for heat and drought stress. J. Am. Soc. Hortic. Sci. 2017, 142, 367–375. [Google Scholar] [CrossRef]
- Cross, J.W.; Bonos, S.A.; Huang, B.; Meyer, W.A. Evaluation of heat and drought as components of summer stress on tall fescue genotypes. HortScience 2013, 48, 1562–1567. [Google Scholar] [CrossRef]
- Huang, B.; Fry, J.; Wang, B. Water relations and canopy characteristics of tall fescue cultivars during and after drought stress. HortScience 1998, 33, 837–840. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef]
- He, C.; Du, Y.; Fu, J.; Zeng, E.; Park, S.; White, F.; Zheng, J.; Liu, S. Early drought-Responsive genes are variable and relevant to drought tolerance. G3 (Bethesda) 2020, 10, 1657–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janiak, A.; Kwasniewski, M.; Sowa, M.; Gajek, K.; Żmuda, K.; Kościelniak, J.; Szarejko, I. No time to waste: Transcriptome study reveals that drought tolerance in barley may me attributed to stressed-like expression patterns that exist before the occurrence of stress. Front. Plant Sci. 2018, 8, 2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B. Mechanisms and strategies for improving drought resistance in turfgrass. Acta Hortic. 2008, 783, 221–227. [Google Scholar] [CrossRef]
- Fischer, K.S.; Johnson, E.C.; Edmeades, G.O. Breeding and selection for drought resistance in tropical maize. In Drought Resistance in Crops with Emphasis on Rice; IRRI: Laguna, Philippines, 1982; pp. 377–399. [Google Scholar]
- Gulman, S.L.; Turner, N.C. Differences in root and shoot development of tomato (Lycopersicon esculentum) varieties across contrasting soil environments. Plant Soil 1978, 49, 127–136. [Google Scholar] [CrossRef]
- Bawazir, A.A.; Idle, D.B. Drought resistance and root morphology in sorghum. Plant Soil 1989, 119, 217–221. [Google Scholar] [CrossRef]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M.G.; Kagale, S. Drought response in wheat: Key genes and regulatory mechanisms controlling root system architecture and transpiration efficiency. Front. Chem. 2017, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, B.M.; Hanna, W.W.; Baxter, L.L.; Raymer, P.L.; Waltz, F.C.; Kowalewski, A.R.; Chandra, A.; Genovesi, A.D.; Wherley, B.G.; Miller, G.L.; et al. ‘DT-1′, a drought-tolerant triploid turf bermudagrass. HortScience 2018, 53, 1711–1714. [Google Scholar] [CrossRef]
- Burton, G.W. Tifway (Tifton 419) bermudagrass. Crop Sci. 1966, 6, 93–94. [Google Scholar] [CrossRef]
- Riley, R.J. Cynodon dactylon Plant Named ‘Riley’s Super Sport’. U.S. Patent PP11181, 18 January 2002. [Google Scholar]
- Youngner, V.B. Santa Ana—A new vegetatively propagated turf bermudagrass developed for California. Weeds Trees Turf. 1967, 6, 34–36. [Google Scholar]
- Gibeault, V.A.; Cockerham, S.T. “El Toro” Zoysiagrass. 1988; 38. [Google Scholar]
- Shearman, R.C.; Riordan, T.P.; Abeyo, B.G.; Heng-Moss, T.M.; Lee, D.J.; Gaussoin, R.E.; Gulsen, O.; Budak, H.; Serba, D.D. Buffalograss: Though Native Turfgrass, USGA Turfgrass and Environmental Research Online. 2006; 5, 1–13.
- Goel, S.; Madan, B. Genetic engineering of crop plants for abiotic stress tolerance. In Emerging Technologies and Management of Crop Stress Tolerance; Ahmad, P., Rasool, S.B.T., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 99–123. [Google Scholar] [CrossRef]
- Bushman, B.S.; Robbins, M.D.; Warnke, S.E.; Martin, R.; Harris-Shultz, K.; Amundsen, K.E. Gene expression differences for drought stress response in three cool-season turfgrasses. Int. Turfgrass Soc. Res. J. 2020, 14, 694–700. [Google Scholar] [CrossRef]
- Amundsen, K.; Warnke, S.E.; Bushman, B.S.; Robbins, M.D.; Martin, R.; Harris-Shultz, K. Colonial bentgrass transcript-expression differences compared with creeping bentgrass in response to water-deficit stress. Crop Sci. 2021, 61, 2135–2147. [Google Scholar] [CrossRef]
- Zhao, J.; Yuan, S.; Zhou, M.; Yuan, N.; Li, Z.; Hu, Q.; Bethea, F.G., Jr.; Liu, H.; Li, S.; Luo, H. Transgenic creeping bentgrass overexpressing Osa-miR393a exhibits altered plant development and improved multiple stress tolerance. Plant Biotechnol. J. 2019, 17, 233–251. [Google Scholar] [CrossRef] [Green Version]
- Sarmast, M.K. Ameliorating drought-induced stress in turfgrass through genetic manipulation. In Water Stress Plants; Rahman, I.M.M., Rahman, M., Begum, Z.A., Hasegawa, H., Eds.; IntechOpen: Rijeka, Croatia, 2016. [Google Scholar] [CrossRef] [Green Version]
- Asada, K. Ascorbate peroxidase—A hydrogen peroxide-scavenging enzyme in plants. Physiol. Plant 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Jennifer, A.D.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Feng, Z.; Zhang, B.; Ding, W.; Liu, X.; Yang, D.-L.; Wei, P.; Cao, F.; Zhu, S.; Zhang, F.; Mao, Y.; et al. Efficient genome editing in plants using a CRISPR/Cas system. Cell Res. 2013, 23, 1229–1232. [Google Scholar] [CrossRef] [Green Version]
- Barnes, B.D.; Kopecký, D.; Lukaszewski, A.J.; Baird, J.H. Evaluation of turf-type interspecific hybrids of meadow fescue with perennial ryegrass for improved stress tolerance. Crop Sci. 2014, 54, 355–365. [Google Scholar] [CrossRef]
- Pepin, G.W.; Funk, C.R. Intraspecific hybridization as a method of breeding kentucky bluegrass (Poa pratemis L.) for turf. Crop Sci. 1971, 11, 445–448. [Google Scholar] [CrossRef]
- Warnke, S.E. Detection of interspecific hybrids between creeping bentgrass and colonial bentgrass using DNA marker analysis. Agrosystems Geosci. Environ. 2020, 3, e20048. [Google Scholar] [CrossRef]
- Reasor, E.H.; Brosnan, J.T.; Trigiano, R.N.; Elsner, J.E.; Henry, G.M.; Schwartz, B.M. The genetic and phenotypic variability of interspecific hybrid bermudagrasses (Cynodon dactylon (L.) Pers. × C. transvaalensis Burtt-Davy) used on golf course putting greens. Planta 2016, 244, 761–773. [Google Scholar] [CrossRef] [Green Version]
- Bushman, B.S.; Warnke, S.E. Genetic and genomic approaches for improving turfgrass. Turfgrass Biol. Use Manag. 2013, 56, 683–711. [Google Scholar] [CrossRef]
- Phillips, A.; Seetharam, A.; AuBuchon-Elder, T.; Buckler, E.; Gillespie, L.; Hufford, M.; Llaca, V.; Romay, M.C.; Soreng, R.J.; Kellogg, E.A.; et al. A happy accident: A novel turfgrass reference genome. bioRxiv 2022. [Google Scholar] [CrossRef]
- Huang, B.; Fry, J.D. Turfgrass evapotranspiration. J. Crop Prod. 2000, 2, 317–333. [Google Scholar] [CrossRef]
- Hayden, L.; Cadenasso, M.L.; Haver, D.; Oki, L.R. Residential landscape aesthetics and water conservation best management practices: Homeowner perceptions and preferences. Landsc. Urban Plan. 2015, 144, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Huang, B. Mowing height effects on summer turf growth and physiological activities for two creeping bentgrass cultivars. HortScience 2003, 38, 444–448. [Google Scholar] [CrossRef]
- Salaiz, T.A.; Horst, G.L.; Shearman, R.C. Mowing height and vertical mowing frequency effects on putting green quality. Crop Sci. 1995, 35, 1422–1425. [Google Scholar] [CrossRef]
- Gelernter, W.D.; Stowell, L.J.; Johnson, M.E.; Brown, C.D. Documenting trends in energy use and environmental practices on US golf courses. Crop Forage Turfgrass Manag. 2017, 3, 1–7. [Google Scholar] [CrossRef]
- Cathey, S.E.; Kruse, J.K.; Sinclair, T.R.; Dukes, M.D. Tolerance of three warm-season turfgrasses to increasing and prolonged soil water deficit. HortScience 2011, 46, 1550–1555. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.; Fry, J.D. Water relations and drought tolerance of four turfgrasses. J. Am. Soc. Hortic. Sci. 1997, 122, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Hejl, R.W.; Wherley, B.G.; Fontanier, C.H. Long-term Performance of Warm-season Turfgrass Species under Municipal Irrigation Frequency Restrictions. HortScience 2021, 56, 1221–1225. [Google Scholar] [CrossRef]
- Kneebone, W.R.; Pepper, I.L. Consumptive water use by sub-irrigated turfgrasses under desert conditions. Agron. J. 1982, 74, 419–423. [Google Scholar] [CrossRef]
- Barton, L.; Wan, G.G.Y.; Buck, R.P.; Colmer, T.D. Nitrogen increases evapotranspiration and growth of a warm-season turfgrass. Agron. J. 2009, 101, 17–24. [Google Scholar] [CrossRef]
- Chhetri, M.; Fontanier, C.; Koh, K.; Wu, Y.; Moss, J.Q. Turf performance of seeded and clonal bermudagrasses under varying light environments. Urban For. Urban Green 2019, 43, 126355. [Google Scholar] [CrossRef]
- Chang, Z.; Liu, Y.; Dong, H.; Teng, K.; Han, L.; Zhang, X. Effects of cytokinin and nitrogen on drought tolerance of creeping bentgrass. PLoS ONE 2016, 11, e0154005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saud, S.; Fahad, S.; Cui, G.; Yajun, C.; Anwar, S. Determining nitrogen isotopes discrimination under drought stress on enzymatic activities, nitrogen isotope abundance and water contents of Kentucky bluegrass. Sci. Rep. 2020, 10, 6415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saud, S.; Fahad, S.; Yajun, C.; Ihsan, M.Z.; Hammad, H.M.; Nasim, W.; Amanullah, Jr.; Arif, M.; Alharby, H. Effects of nitrogen supply on water stress and recovery mechanisms in kentucky bluegrass plants. Front. Plant Sci. 2017, 8, 983. [Google Scholar] [CrossRef] [Green Version]
- Henry, J.M.; Gibeault, V.A. Paspalum vaginatum winter color management study. Calif. Turfgrass Cult. 1985, 35, 4–7. [Google Scholar]
- Van Dam, J. The economics of overseeding and colorizing dormant bermudagrass. Calif. Turfgrass Cult. 1972, 22, 26–28. [Google Scholar]
- Van Dam, J.; Kurtz, K. A turfgrass colorant study. Calif. Turfgrass Cult. 1971, 21, 17–20. [Google Scholar]
- Youngner, V.B.; Fuchigami, T. Colorants for dormant bermuda and other subtropical grasses. South Calif. Turfgrass Cult. 1958, 8, 7–9. [Google Scholar]
- Shearman, R.C.; Wit, L.A.; Severmutlu, S.; Budak, H.; Gaussoin, R.E. Colorant effects on dormant buffalograss turf performance. HortTechnology 2005, 15, 244–246. [Google Scholar] [CrossRef]
- Liu, H.; McCarty, B.; Baldwin, C.M.; Sarvis, W.G.; Long, S.H. Painting dormant bermudagrass putting greens. Golf Course Mgt 2007, 75, 86–91. [Google Scholar]
- Hargey, D.; Wherley, B.; Reynolds, C.; White, R.; Parker, G. Performance of winter overseeded and colorant-treated ‘Tifway’ bermudagrass receiving weekly irrigation and simulated traffic. HortScience 2016, 51, 580–587. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.C. Principles of Turfgrass Irrigation. PennState Ext. 2016. Available online: https://extension.psu.edu (accessed on 28 June 2022).
- Ledieu, J.; De Ridder, P.; De Clerck, P.; Dautrebande, S. A method of measuring soil moisture by time-domain reflectometry. J. Hydrol. 1986, 88, 319–328. [Google Scholar] [CrossRef]
- Rouse, J.W. Monitoring the Vernal Advancement and Retrogradation of Natural Vegetation; No. RSC-1978-2; Texas A&M University: College Station, TX, USA, 1973. [Google Scholar]
- Kim, D.M.; Zhang, H.; Zhou, H.; Du, T.; Wu, Q.; Mockler, T.C.; Berezin, M.Y. Highly sensitive image-derived indices of water-stressed plants using hyperspectral imaging in SWIR and histogram analysis. Sci. Rep. 2015, 5, 15919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamsen, F.J.; Pinter, P.J., Jr.; Barnes, E.M.; LaMorte, R.L.; Wall, G.W.; Leavitt, S.W.; Kimball, B.A. Measuring wheat senescence with a digital camera. Crop Sci. 1999, 39, 719–724. [Google Scholar] [CrossRef]
- Huete, A.R. Remote sensing for environmental monitoring. In Environmental Monitoring and Characterization; Artiola, J.F., Pepper, I.L., Brusseau, M.L., Eds.; Academic Press: Burlington, VT, USA, 2004; pp. 183–206. [Google Scholar] [CrossRef]
- An, N.; Goldsby, A.L.; Price, K.P.; Bremer, D.J. Using hyperspectral radiometry to predict the green leaf area index of turfgrass. Int. J. Remote Sens. 2015, 36, 1470–1483. [Google Scholar] [CrossRef]
- Grabow, G.L.; Ghali, I.E.; Huffman, R.L.; Miller, G.L.; Bowman, D.; Vasanth, A. Water application efficiency and adequacy of ET-based and soil moisture—based irrigation controllers for turfgrass irrigation. J. Irrig. Drain Eng. 2013, 139, 113–123. [Google Scholar] [CrossRef]
- Moeller, A.D. Identify soil moisture status more accurately than ever before. Michigan State Univ. Greeen Sect. Rec. 2012, 50, 1–5. [Google Scholar]
- Carrow, R.N.; Krum, J.M.; Flitcroft, I.; Cline, V. Precision turfgrass management: Challenges and field applications for mapping turfgrass soil and stress. Precis. Agric. 2010, 11, 115–134. [Google Scholar] [CrossRef]
- Serena, M.; Velasco-Cruz, C.; Friell, J.; Schiavon, M.; Sevostianova, E.; Beck, L.; Sallenave, R.; Leinauer, B. Irrigation scheduling technologies reduce water use and maintain turfgrass quality. Agron. J. 2020, 112, 3456–3469. [Google Scholar] [CrossRef]
- Hejl, R.W.; Wherley, B.G.; McInnes, K.; Straw, C.M.; Fontanier, C. Evaluation of irrigation scheduling approaches within sand-capped turfgrass systems. Agron. J. 2022, 114, 1694–1704. [Google Scholar] [CrossRef]
- Krum, J.M.; Carrow, R.N.; Karnok, K. Spatial mapping of complex turfgrass sites: Site-specific management units and protocols. Crop Sci. 2010, 50, 301–315. [Google Scholar] [CrossRef]
- Straw, C.M.; Wardrop, W.S.; Horgan, B.P. Golf course superintendents’ knowledge of variability within fairways: A tool for precision turfgrass management. Precis. Agric. 2020, 21, 637–654. [Google Scholar] [CrossRef]
- Hejl, R.; Straw, C.; Wherley, B.; Bowling, R.; McInnes, K. Factors leading to spatiotemporal variability of soil moisture and turfgrass quality within sand-capped golf course fairways. Precis. Agric. 2022, 23, 1908–1917. [Google Scholar] [CrossRef]
- Barnes, M.R.; Yue, C.; Watkins, E. Homeowner perceptions of watering restriction scenarios in the Minneapolis–St. Paul metropolitan area. Crop Forage Turfgrass Manag. 2021, 7, e20131. [Google Scholar] [CrossRef]
- Wheeler, M.M.; Larson, K.L.; Andrade, R. Attitudinal and structural drivers of preferred versus actual residential landscapes in a desert city. Urban Ecosyst. 2020, 23, 659–673. [Google Scholar] [CrossRef]
- Ignatieva, M.; Haase, D.; Dushkova, D.; Haase, A. Lawns in cities: From a globalised urban green SPACE phenomenon to sustainable nature-based solutions. Land 2020, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Chan, Z.; Shi, H. Improved abiotic stress tolerance of bermudagrass by exogenous small molecules. Plant Signal. Behav. 2015, 10, e991577. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Yin, S.; Gan, L.; Zhang, X. Mitigating effect of glycinebetaine pretreatment on drought stress responses of creeping bentgrass. HortScience 2018, 53, 1842–1848. [Google Scholar] [CrossRef] [Green Version]
- Oddo, E.; Russo, G.; Grisafi, F. Effects of foliar application of glycine betaine and chitosan on Puccinellia distans (Jacq.) Pari, subjected to salt stress. Biol. Futur. 2019, 70, 47–55. [Google Scholar] [CrossRef]
- Rademacher, W. Plant growth regulators: Backgrounds and uses in plant production. J. Plant Growth Regul. 2015, 34, 845–872. [Google Scholar] [CrossRef]
- Fidanza, M.A.; Wetzel Iii, H.C.; Agnew, M.L.; Kaminski, J.E. Evaluation of fungicide and plant growth regulator tank-mix programmes on dollar spot severity of creeping bentgrass. Crop Prot. 2006, 25, 1032–1038. [Google Scholar] [CrossRef]
- March, S.R.; Martins, D.; McElroy, J.S. Growth inhibitors in turfgrass. Plant. Daninha 2013, 31, 733–747. [Google Scholar] [CrossRef] [Green Version]
- Glab, T.; Szewczyk, W.; Gondek, K.; Knaga, J.; Tomasik, M.; Kowalik, K. Effect of plant growth regulators on visual quality of turfgrass. Sci. Hortic. 2020, 267, 109314. [Google Scholar] [CrossRef]
- Cohen, I.; Netzer, Y.; Sthein, I.; Gilichinsky, M.; Tel-Or, E. Plant growth regulators improve drought tolerance, reduce growth and evapotranspiration in deficit irrigated Zoysia japonica under field conditions. Plant Growth Regul. 2019, 88, 9–17. [Google Scholar] [CrossRef]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Mark Cigan, A. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dlamini, P.J. Drought stress tolerance mechanisms and breeding effort in sugarcane: A review of progress and constraints in South Africa. Plant Stress 2021, 2, 100027. [Google Scholar] [CrossRef]
- Dunning, S. Evaluation of Ornamental Grasses for Use as Golf Course Plantings; UF/IFAS Extension; University of Florida: Gainesville, FL, USA, 2014; 23p. [Google Scholar]
- Burayu, W.; Umeda, K. Versatile native grasses and a turf-alternative groundcover for the arid southwest United States. J. Environ. Hortic. 2021, 39, 160–167. [Google Scholar] [CrossRef]
- Watkins, E.; Brilman, L.; Kopec, D. Development of native grasses for turf. Turfgrass Biol. Use Manag. 2013, 56, 661–681. [Google Scholar] [CrossRef]
- Chadde, S.W. Grasses of the Southwest: A Key to Common Species Using Vegetative Features; Orchard Innovations: Sunnyvale, CA, USA, 2020. [Google Scholar]
- Evanylo, G.; Ervin, E.; Zhang, X. Reclaimed water for turfgrass irrigation. Water 2010, 2, 685–701. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.; Semiat, R.; Duke, M.; Rahardianto, A.; Cohen, Y. Seawater use and desalination technology. In Treatise Water Science; Wilderer, P.B.T., Ed.; Elsevier: Oxford, UK, 2011; pp. 73–109. [Google Scholar] [CrossRef]
- Sevostianova, E.; Leinauer, B.; Sallenave, R.; Karcher, D.; Maier, B. Soil salinity and quality of sprinkler and drip irrigated warm-season turfgrasses. Agron. J. 2011, 103, 1773–1784. [Google Scholar] [CrossRef]
- Schaan, C.M.; Devitt, D.A.; Morris, R.L.; Clark, L. Cyclic irrigation of turfgrass using a shallow saline aquifer. Agron. J. 2003, 95, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Dean, D.E.; Devitt, D.A.; Verchick, L.S.; Morris, R.L. Turfgrass quality, growth, and water use influenced by salinity and water stress. Agron. J. 1996, 88, 844–849. [Google Scholar] [CrossRef]
- Zalacáin, D.; Martínez-Pérez, S.; Bienes, R.; García-Díaz, A.; Sastre-Merlín, A. Turfgrass biomass production and nutrient balance of an urban park irrigated with reclaimed water. Chemosphere 2019, 237, 124481. [Google Scholar] [CrossRef] [PubMed]
- Hull, R.J. Sodium plays role in certain turfgrass processes. Turfgrass Trends 2004, 64–67. [Google Scholar]
- Ashraf, M.; Athar, H.R.; Harris, P.J.C.; Kwon, T.R. Some Prospective Strategies for Improving Crop Salt Tolerance; Academic Press: San Diego, CA, USA, 2008; Volume 97, pp. 45–110. [Google Scholar] [CrossRef]
- Song, J.; Wang, B. Using euhalophytes to understand salt tolerance and to develop saline agriculture: Suaeda salsa as a promising model. Ann. Bot. 2015, 115, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Chang, B.; Wherley, B.; Aitkenhead-Peterson, J.; Ojeda, N.; Fontanier, C.; Dwyer, P. Effect of wetting agent on nutrient and water retention and runoff from simulated urban lawns. HortScience 2020, 55, 1005–1013. [Google Scholar] [CrossRef]
- Xiang, M.; Schiavon, M.; Orlinski, P.; Forconi, A.; Baird, J.H. Identification of wetting agents for water conservation on deficit-irrigated hybrid bermudagrass fairways. Agron. J. 2021, 113, 3846–3856. [Google Scholar] [CrossRef]
- Kostka, S.J.; Cisar, J.L.; Mitra, S.; Park, D.M.; Ritsema, C.J.; Dekker, L.W.; Franklin, M.A. Irrigation efficiency: Surfactants can save water and help maintain turfgrass quality. In Soil Surfactants Can Save Water and Help Maintain Turfgrass Quality; Golf Course Industry: Cuyahoga County, OH, USA, 2007; pp. 91–95. [Google Scholar]
- Kostka, S.J.; Bially, P.T. Synergistic surfactant interactions for enhancement of hydrophilicity in water repellent soils. Int. Turfgrass Soc. Res. J. 2005, 10, 108–114. [Google Scholar]
- Braun, R.C.; Bremer, D.J.; Hoyle, J.A. Simulated traffic on turfgrasses during drought stress: I. Performance and recovery of turf canopies. Crop Sci. 2020, 61, 2926–2938. [Google Scholar] [CrossRef]
- Hejl, R.W.; Wherley, B.G.; White, R.H.; Thomas, J.C.; Fontanier, C.H. Deficit Irrigation and Simulated Traffic on ‘Tifway’ Bermudagrass Summer Performance and Autumn Recovery. Crop Sci. 2016, 56, 809–817. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Cultivar | Release Year | Reference or Source |
|---|---|---|---|
| Bermudagrass (C. dactylon × C. transvaalensis) (C. dactylon) | TifTuf Tifway | 2014 1960 | [48] [49] |
| Celebration * Santa Ana | 2002 1966 | [50] [51] | |
| Zoysiagrass (Zoysia japonica × Z. matrella) | El Toro Empire Zeon | 1986 - 1996 | [52] Developed in Brazil Proprietary |
| Buffalograss (Buchloe dactyloides) | Prestige Legacy | 2003 2000 | [53] [53] |
| St. Augustine (Stenotaphrum secundatum) | Palmetto Citrablue | 1990 2018 | Proprietary University of Florida |
| Seashore paspalum (Paspalum vaginatum) | SeaDwarf Salam Sea Star+ | - 1990s 2014 | Proprietary Proprietary University of Georgia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serba, D.D.; Hejl, R.W.; Burayu, W.; Umeda, K.; Bushman, B.S.; Williams, C.F. Pertinent Water-Saving Management Strategies for Sustainable Turfgrass in the Desert U.S. Southwest. Sustainability 2022, 14, 12722. https://0-doi-org.brum.beds.ac.uk/10.3390/su141912722
Serba DD, Hejl RW, Burayu W, Umeda K, Bushman BS, Williams CF. Pertinent Water-Saving Management Strategies for Sustainable Turfgrass in the Desert U.S. Southwest. Sustainability. 2022; 14(19):12722. https://0-doi-org.brum.beds.ac.uk/10.3390/su141912722
Chicago/Turabian StyleSerba, Desalegn D., Reagan W. Hejl, Worku Burayu, Kai Umeda, Bradley Shaun Bushman, and Clinton F. Williams. 2022. "Pertinent Water-Saving Management Strategies for Sustainable Turfgrass in the Desert U.S. Southwest" Sustainability 14, no. 19: 12722. https://0-doi-org.brum.beds.ac.uk/10.3390/su141912722