3.1. Effect of CM on Enzyme Concentration
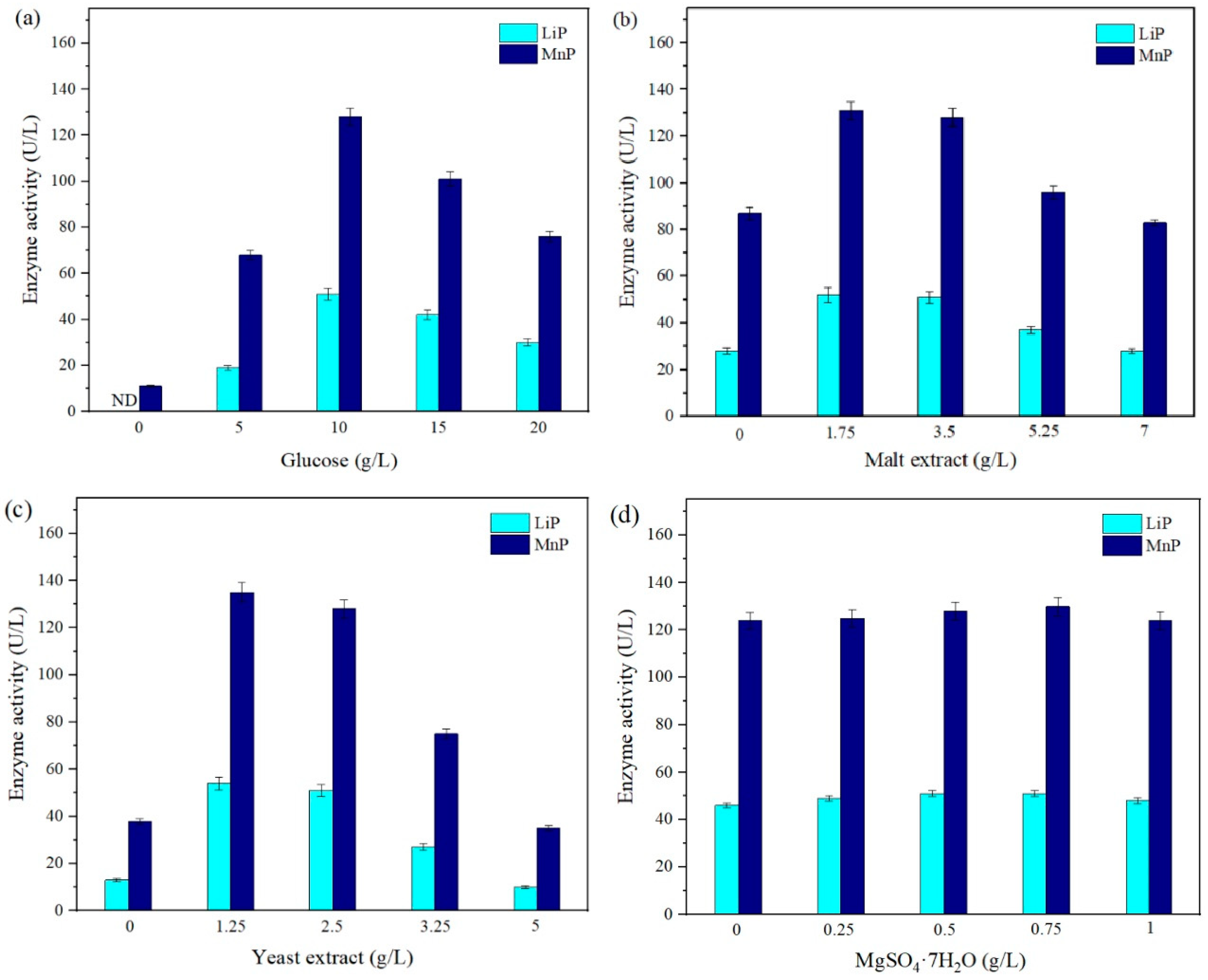
Enzyme activity in response to variations in the concentration of each ingredient in the standard CM is displayed in
Figure 2. According to the results, there was a negligible influence of the concentration of MgSO
4·7H
2O in the range of 0–1 g/L, whilst the variation in concentration of the rest of the medium components was more significant during the growth of
P. chrysosporium. From
Figure 2a, an increasing concentration of glucose up to 10 g/L boosted the secretion of extracellular enzymes, attaining 51 U/L LiP and 128 U/L MnP. However, the availability of greater concentrations apparently depressed microbial activity. The secretion of enzymes by
P. chrysosporium is denominated as secondary metabolism, depending on nutrient limitation conditions [
27]. Glucose, a conventional carbohydrate used in CM, is categorized as a simple sugar (monosaccharide), serving as a rich C source for microorganisms to use as a growth substrate [
21]. As the primary source of C in the selected CM, glucose was essential in the secretion of enzymes in the 0–10 g/L range, requiring at least 10 g/L to obtain a better performance.
Notably, glucose removal significantly affected the enzyme activity of
P. chrysosporium, resulting in 0 U/L LiP and 11 U/L MnP, which could be ascribed to the unavailability of simple C sources to support microbial activity [
27,
28]. From
Figure 2a,
P. chrysosporium could likely require a glucose concentration higher than 5 g/L to attain an optimal fungal metabolism. A supply of 5 g/L glucose in the CM could have also experienced an earlier development of enzymes due to the faster glucose consumption. Thus, the onset of enzymes could have occurred before 7 d, showing a lower concentration on day 7. In the same way, concentrations higher than 10 g/L glucose could have undergone a later development of enzyme activity having an onset after day 7 and, therefore leading to a lower concentration of enzymes at day 7 of incubation. Concentrations exceeding 15 g/L displayed detrimental influence as white-rot fungi such as
P. chrysosporium might require a C-limited environment to favor the secretion of extracellular enzymes, which turned out in 30 U/L LiP and 76 U/L MnP.
Based on
Figure 2b, the optimal concentration of malt extract was in the range of 1.75 to 3.5 g/L, reaching a maximum of 52 U/L LiP and 131 U/L MnP at a concentration of 1.75 g/L malt extract. Apparently, any increase or decrease outside this range of concentration had a negative impact on the secretion of LiP and MnP. Malt extract, a compound derived from malted barley, is mainly a source of carbohydrates encompassing reduced sugars (>90%) such as maltose, fructose, and sucrose and also has a smaller proportion of N constituents such as amino acids, peptides, and minerals [
29,
30]. A higher concentration of over 3.5 g/L of this component could have created nutrient-rich conditions, affecting the secondary metabolism of the microorganism. Further, the ideal behavior of the fungi was also affected by the removal of malt extract (0 g/L) but was probably supported by other nourishing medium components such as yeast extract and glucose, resulting in a concentration of 28 U/L LiP and 87 U/L MnP.
Comparably to malt extract, a concentration of yeast extract in the range of 1.25–2.5 g/L was more favorable for enzyme secretion, attaining a maximum of 54 U/L LiP and 135 U/L MnP, as shown in
Figure 2c. A concentration over this range was ineffective in improving the microbial activity of
P. chrysosporium, probably due to excess nutrients in the CM. In addition, the elimination of yeast extract (0 g/L) caused a sharp decrease in enzyme concentration (13 U/L LiP and 38 U/L MnP), highlighting the importance of this medium component in the CM. Yeast extract is a nourishing compound and is one of the primary sources of N, vitamins (biotin and vitamin B), and TE, e.g., manganese (Mn), magnesium (Mg), calcium (Ca), phosphorus (P), Zinc (Zn), iron (Fe), Cobalt (Co), as well as proteins (mainly nucleic acid and glutathione) and amino acids required for the metabolism, growth, and better function of this microorganism [
31]. Therefore, its composition makes yeast extract a highly indispensable ingredient in the CM. Contrary to the rest of the medium ingredients, there was an insignificant influence of the concentration of MgSO
4·7H
2O in the range of 0–1 g/L, resulting in similar concentrations of enzymes at different concentrations, which could be due to the supplementation of rich ingredients in the standard CM that have TE of a wide variety of minerals and nutrients, e.g., Mg, Mn.
3.2. Effect of CM on Sulfide Bio-Oxidation
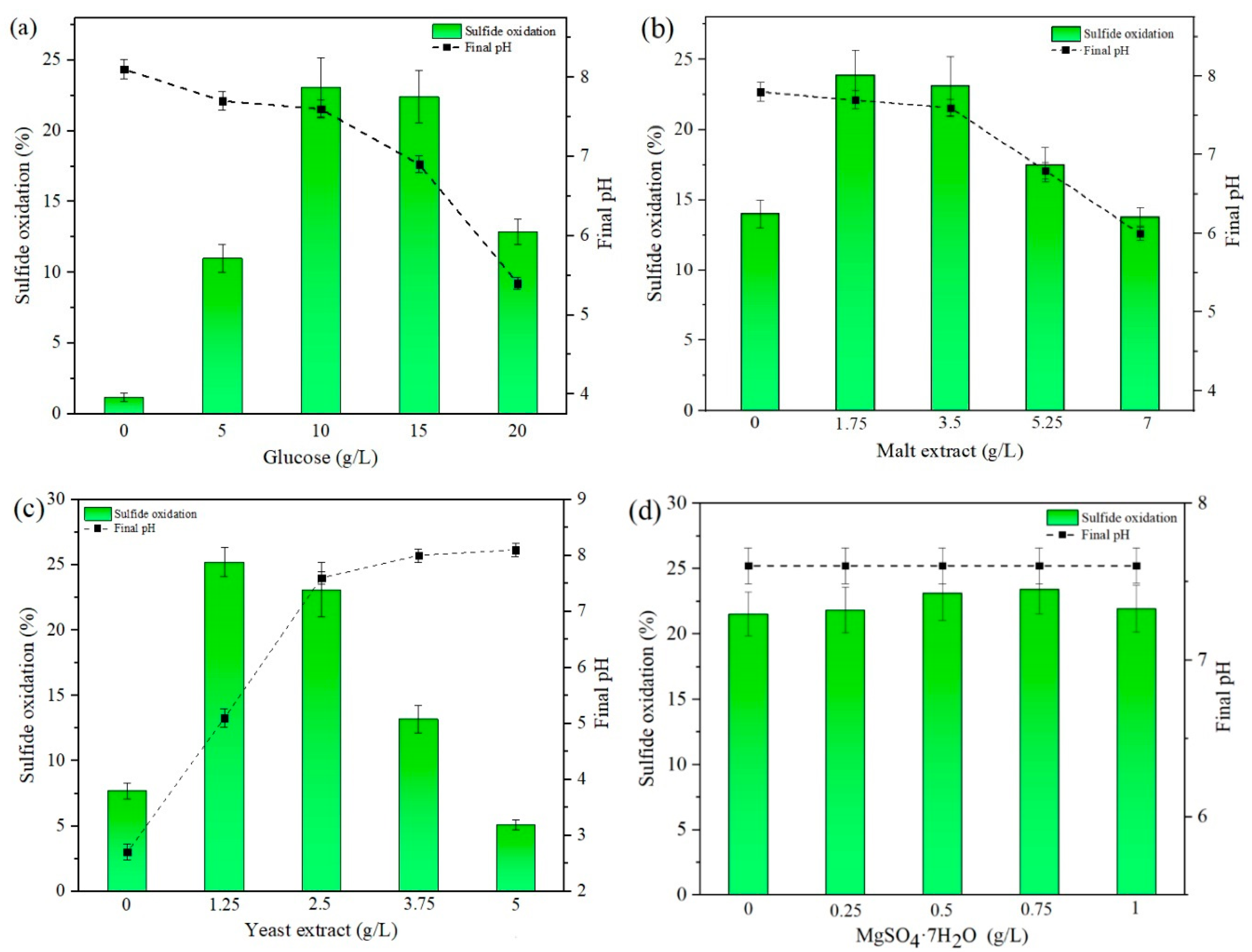
Figure 3 displays the changes observed in sulfide oxidation and pH solution after 14 d of contact time between
P. chrysosporium and the sulfidic ore. From
Figure 3, it was deduced that sulfide oxidation varied from nearly 1.2% to 25.2%, considering individual changes in the concentration of medium components, which generally coincided with the trends in enzyme activity. The mechanism of the white-rot fungi to alter reduced sulfur species depends on the secretion of extracellular enzymes, including LiP and MnP. The catalytic cycle of oxidative enzymes has been explained in previous publications [
4,
6] and essentially consists of a chain of oxidation-reduction reactions favoring the oxidation of reduced sulfur species and the reduction of enzymatic compounds secreted by
P. chrysosporium. However, the mechanism still needs further research to fully understand this type of system.
Concerning the experiments with variations in glucose concentration (
Figure 3a), a concentration between 10 to 15 g/L resulted in ca. 23% oxidation, whilst 0 and 20 g/L reduced the capacity to oxidize the sulfidic ore, generating 1.2% and 12.9%, correspondently.
Furthermore, 1.75–3.5 g/L malt extract and 1.25–2.5 g/L yeast extract led to better sulfide oxidation compared to other concentrations of these reagents, displaying a maximum of 23.9% and 25.2%, correspondingly (
Figure 3b,c). A concentration out of these two ranges of concentration led to a decrease in sulfide oxidation, reaching 13.8% and 5.1% at 20 g/L malt extract and yeast extract, respectively. The addition of different concentrations of MgSO
4·7H
2O in the interval of 0–1 g/L had an insignificant influence compared to other parameters, such as yeast extract and glucose, resulting in similar oxidation (ca. 23%) for all the evaluated conditions.
Sulfide oxidation increased proportionally to glucose concentration in the CM up to the range of 10 to 15 g/L, and subsequently, sulfide oxidation decreased at higher concentrations. This trend is an outcome of two coexisting factors: (1) glucose, a simple source of C, serves as the primary source of C supporting the microbial activity during the initial growth, and (2) the availability of higher concentration of glucose could prevent the secretion of ligninolytic enzymes under nutrient-rich cultures [
27,
28]. The higher sulfide oxidation observed in the concentration range of 1.75–3.5 g/L malt extract and 1.25–2.5 g/L yeast extract could be associated with the fact that a nutrient deficiency, but sufficiently enough nutrients to support the fungal activity benefits the microbial activity secreting a higher concentration of enzymes and therefore better sulfide oxidation [
20,
21]. The effect of MgSO
4·7H
2O on sulfide oxidation could be interpreted differently compared to other reagents in the CM because it had a negligible impact on ligninolytic enzyme secretion (
Figure 2d) and sulfide oxidation (
Figure 3d) in the evaluated interval. Even though the concentration of Mg has been reported to favor fungal growth [
32], it seems that the concentration of yeast extract and malt extract in the evaluated CM could have been sufficient to provide the Mg content and other TE required for fungal growth [
30,
31].
The initial pH for all the tested conditions was previously adjusted to pH 5.8 before starting the tests. However, major pH changes were measured after 14 d of bio-oxidation, as displayed in
Figure 3. Results showed that the final pH fluctuated from 2.7 to 8.1, depending on the concentrations of each reagent in the CM. A more notable decrease in pH was recorded when yeast extract was removed entirely from the CM (0 g/L), resulting in pH 2.7. By adding yeast extract to the CM, the pH changes could be attributed to the buffering properties of this compound and microbial activity in the CM [
33,
34]. The pH trends were similar for glucose and malt extract; a rise in their concentration produced a pH drop more remarkably. Glucose and malt extract are metabolized by white-rot fungi, releasing organic acids and protons into the solution and modifying pH [
34,
35]. Therefore, an increase in glucose and malt extract concentration seems to have caused a higher organic acid concentration and a more significant drop in the pH solution, recording 5.4 and 6.0 at 20 g/L and 7 g/L, respectively. Regarding the effect of different concentrations of MgSO
4·7H
2O on pH changes, the final pH remained at ca. 7.6 for all the evaluated concentrations, which seems to be reasonable based on the similarity in sulfide oxidation found in
Figure 3d.
3.3. Effect of Interaction of Medium Components on Sulfide Oxidation
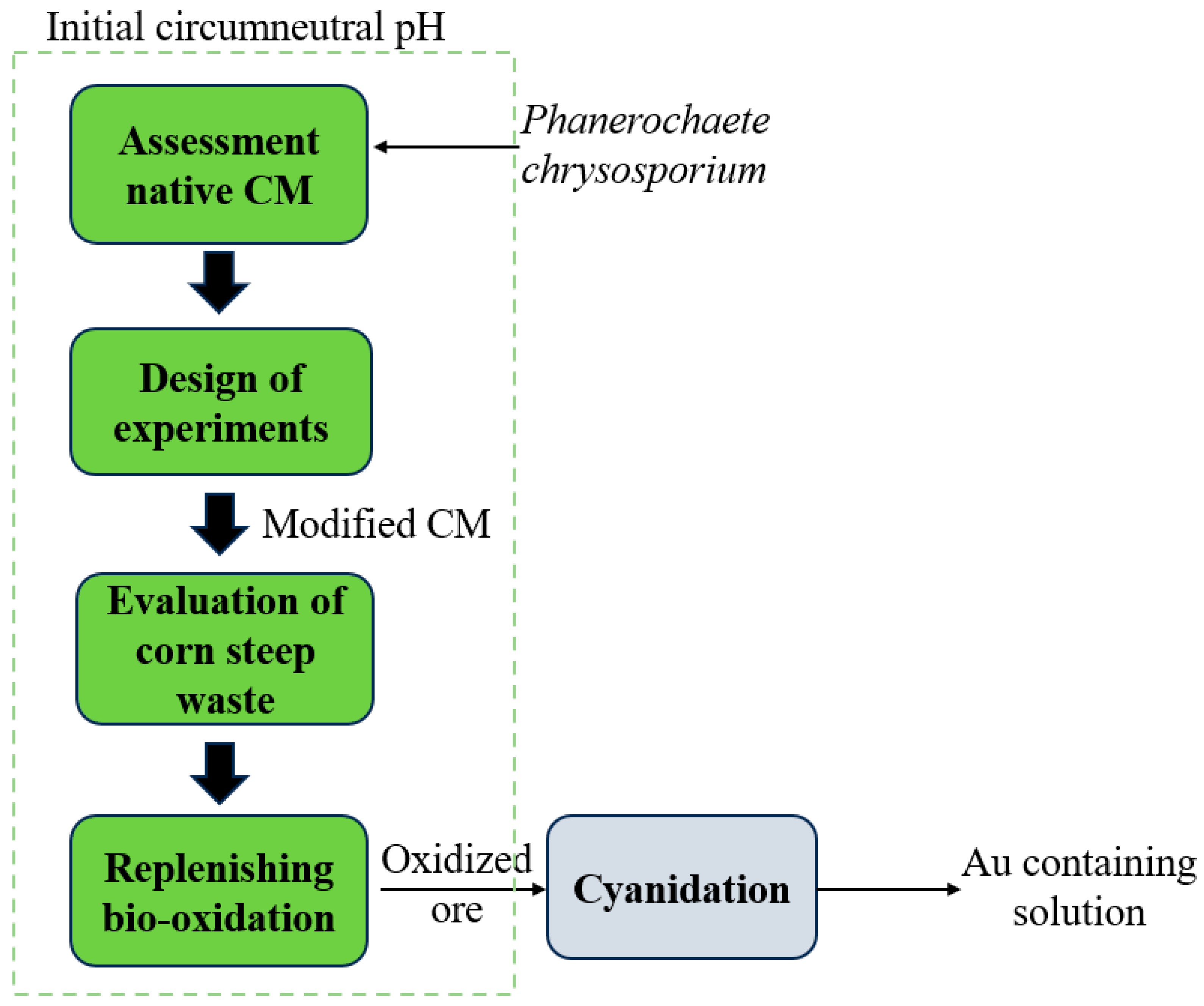
After analyzing the role of each component in the standard CM, a set of tests was suggested to optimize the bio-oxidation of the sulfidic ore. The experiments were proposed based on D-optimal RSM design, including 25 experiments displayed in
Table S1 (supplementary material) to obtain modified combinations of ingredients at different concentrations for more efficient bio-oxidation. The design contemplated the influential concentration of glucose (10–20 g/L), malt extract (0–3.5 g/L), yeast extract (0–2.5 g/L), and MgSO
4·7H
2O (0–0.5 g/L) found in the preliminary study. The design provides an overview of the relevance of individual parameters and their interactions, resulting in a model to predict bio-oxidation. The software outcomes are presented in diagrams of sulfide oxidation versus medium ingredients and ANOVA chart.
Table 3 shows a summary of the statistical data obtained from the multiple linear regression model. In order to guarantee a model reliability within 95% of the confidence, a
p-value (probability value) of the model is required to be less than 0.05 (F-value higher than 4) with a lack of fit
p-value greater than 0.05 [
36]. The model
p-value of <0.0001 indicates that it is significant enough and that there is a chance of only 0.01% that the model could take place because of noise. It is also desirable for a model to have a non-significant lack of fit. The lack of fit
p-value of the model resulted in 0.77, implying that it is not significant to pure error.
Table 3 also displays that most of the individual component concentrations were effective terms, as well as some interactions, such as glucose–malt extract. According to the simulated and experimental data analyzed using the software design expert, a quadratic model was proposed with an R
2 of 0.92 (
Table S2 in supplementary material). The modeled trends provided using the software concur with the experimental data and the prediction of the role of each evaluated ingredient concerning sulfide oxidation. The interactions of different parameters over bio-oxidation experiments are shown in
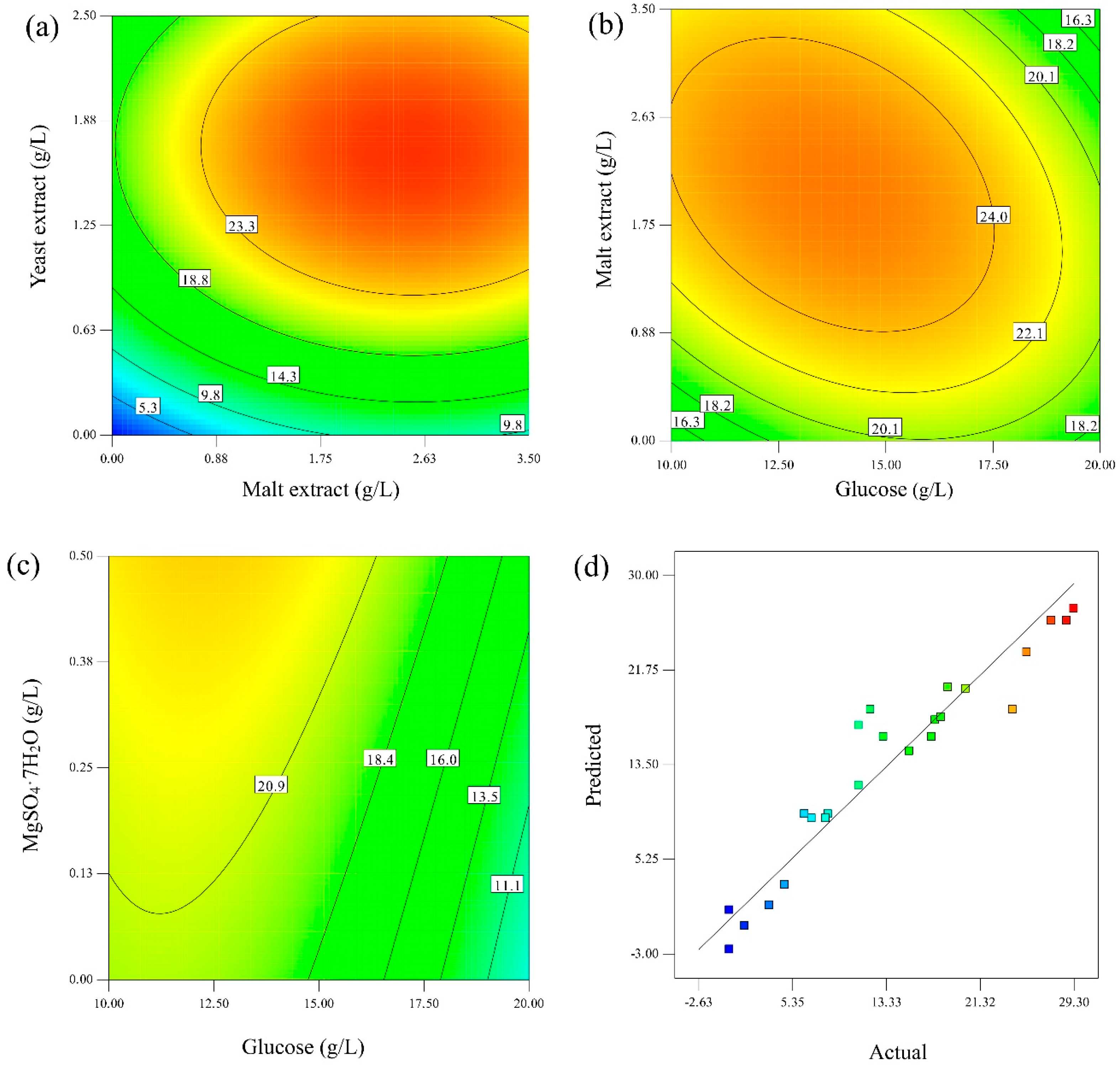
Figure 4.
Using a concentration of yeast extract <1 g/L, the concentration of malt extract had a negligible effect on enhancing the sulfide oxidation rate (
Figure 4a). Additionally, as both variables approached close to 0 g/L, a considerable reduction in sulfide oxidation was observed, demonstrating the importance of these components in the CM to sustain the microbial activity of the fungus. Variations in the concentration of malt extract became a more influential factor at a yeast extract concentration >1 g/L. If a concentration of 1.70 g/L yeast extract was provided, a change in malt extract concentration in the range of 0.80 to 3.5 was more significant in sulfide oxidation, reaching a maximum of 27.8% sulfide oxidation at 2.35 g/L of malt extract followed by a slight drop at a concentration higher than 3.5 g/L malt extract (25.6% sulfide oxidation). Yeast extract above >2.1 g/L produces a drop in sulfide oxidation, probably due to the nutrient-limited condition required by the fungus to produce more extracellular enzymes.
Regarding the interaction between glucose and malt extract shown in
Figure 4b, a concentration of malt extract in the range of 1.0–2.9 g/L resulted in >24% sulfide oxidation using a glucose concentration between 12 to 15 g/L. By keeping a concentration of malt extract at 3.0 g/L, a higher concentration of glucose (>15 g/L) seems detrimental, which could be explained by the excess of nutrients in the system. Malt extract is also an N source in a smaller proportion than yeast extract.
Figure 4b also highlights the importance of having a lower concentration of N sources to support the microbial activity of this microorganism via an N-deficient environment [
27].
Concerning glucose and MgSO
4·7H
2O interaction in
Figure 4c, MgSO
4·7H
2O in the range 0–0.5 g/L was determined to have a negligible effect using 10–15 g/L glucose, as mentioned in the previous section. From
Figure 4d, a good correlation was found between the predicted and actual data, which is indicated by the clustering of points around the diagonal, satisfactorily verifying the model’s robustness. Based on the analysis performed with D-optimal RSM, Equation (3) was suggested to best fit the sulfide oxidation prediction as a function of the medium ingredients with an R
2 of 0.92.
where C represents the concentration of each medium ingredient in g/L.
The value of R
2 = 0.92 suggests that the sulfide oxidation equation effectively characterizes the system using the tested experimental conditions. Results reveal that maximum sulfide oxidation could be attained using a CM composed of 12.86 g/L glucose, 2.20 g/L malt extract, 1.67 g/L yeast extract, and 0.49 g/L MgSO
4·7H
2O. The enzyme activity for LiP and MnP in the modified CM resulted in 58 ± 3 and 143 ± 5 U/L, respectively, after 7 d of incubation. Three confirmation experiments were conducted, and a sulfide oxidation of 28.7 ± 1.6% was reached after 14-d bio-oxidation with a final pH of 6.1 ± 0.2. In our previous research work employing the standard CM (10 g/L glucose, 3.5 g/L malt extract, 2.5 g/L, and 0.5 g/L MgSO
4·7H
2O) with the same ore [
6], sulfide bio-oxidation resulted in 23.1 ± 2.1% with final pH 7.6 ± 0.1 after 14 d. The maximum enzyme activity recorded using the standard CM was 51 ± 3 U/L LiP and 128 ± 4 U/L. Hence, the higher oxidation of the sulfidic ores could be explained by the increase in metabolites likely triggered by a nutrient-deficient CM.
A control test, utilizing the modified CM at identical experimental conditions except for the microbial inoculation, was also carried out to evaluate the impact of the CM on sulfide oxidation. The control tests using the modified CM resulted in 0.6 ± 0.2% sulfide oxidation, which could be associated with the chemical oxidation of sulfide, being negligible compared to the fungi-assisted experiments.
The same fungus has been used to oxidize sulfidic ores in an acidic pH environment. Ofori-Sarpong et al. [
7] tested
P. chrysosporium to oxidize 30%
w/
v sulfidic concentrate at an initial pH of 4.0, attaining 57% sulfide oxidation. In addition, Yang et al. [
8] also employed the microorganism to oxidize 5%
w/
v of almost pure pyrite at an initial pH of 5.0, reporting a maximum of 95 U/L LiP and 316 MnP with a 33% sulfide oxidation and a final pH of 2.1. Compared to previous studies, the performance of
P. chrysosporium appears to be depleted at an initial circumneutral pH using the standard CM and the proposed modified CM. Based on
Table 1, there was generally a decrease in microbial activity when comparing different CM at initial acidic and circumneutral pH. In our previous study [
6], a pH of 4.0 led to a higher concentration of metabolites than a pH of 7.0, which helped obtain a higher sulfide oxidation under similar conditions. An initial pH of 4.0 seems optimal to favor robust microbial growth; nonetheless, this experimental condition did not lead to a final neutral pH after bio-oxidation.
3.4. Effect of pH-Controlled Batch Cultures
After determining the modified CM to maximize sulfide oxidation, experiments were carried out using pH-controlled batch culture to maintain a circumneutral pH throughout bio-oxidation. In our previous study, the cultures with no pH control experienced a decrease in pH to near pH 4.0 once the fungus reached a robust growth after 7 d of incubation, which coincided with the onset of the maximal secretion of ligninolytic enzymes [
6]. Subsequently, pH showed an increasing trend as the bio-oxidation progressed over time, resulting in a final pH > 7.0, which could be mainly attributed to the consumption of organic acid buffer capacity of medium components as a result of fungal activity [
6,
35].
In this section,
P. chrysosporium was tested at pH 5.8, 6.0, 6.5, and 7.0, maintaining the pH solution with phosphate buffer to avoid a significant change in pH throughout the experiments.
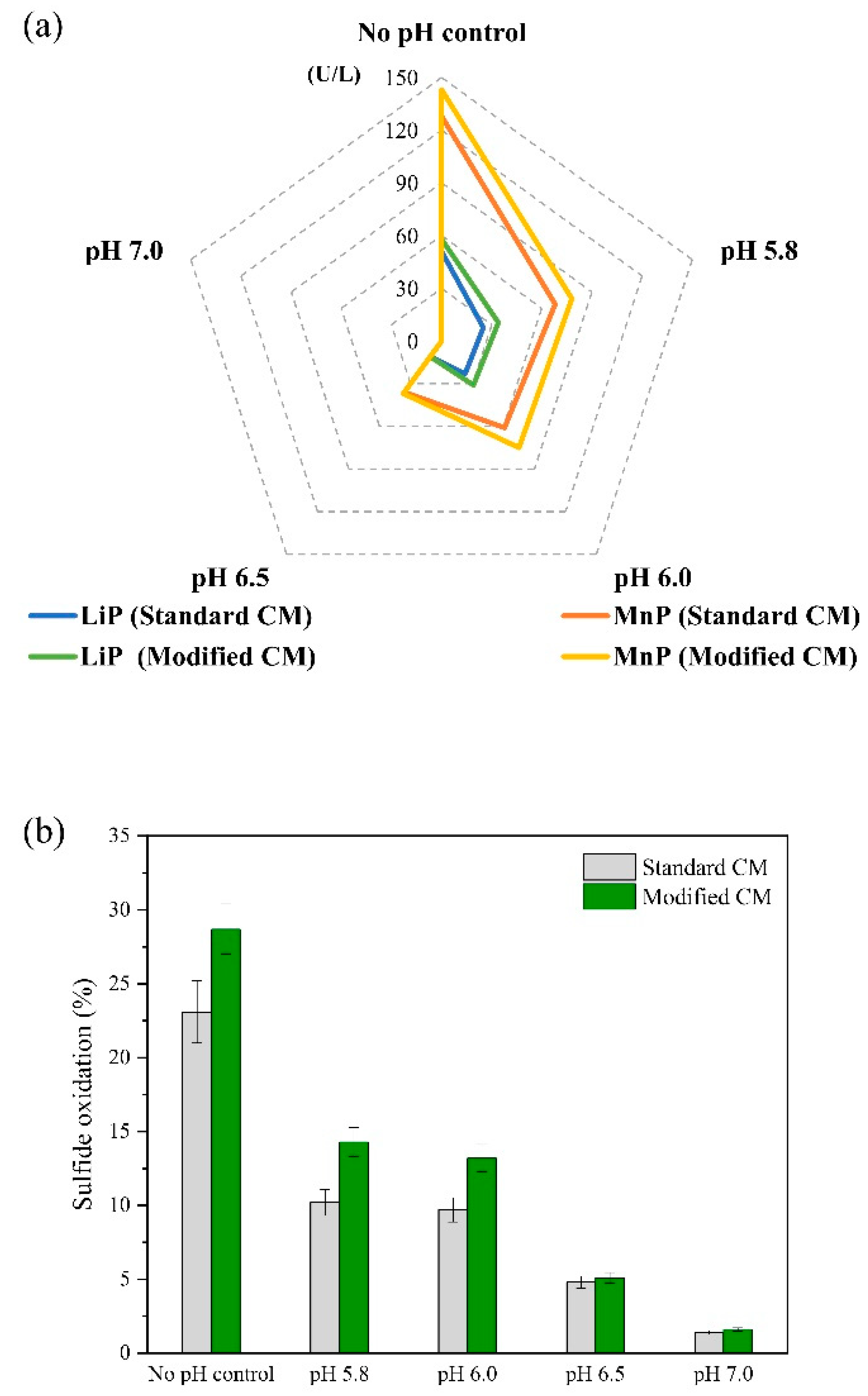
Figure 5 compares the data for enzyme activity and sulfide oxidation using the standard and modified CM under pH-controlled and no pH-control conditions. From
Figure 5a, trends observed in the modified and standard CM were mainly similar for microbial activity. However, as previously reported, the modified CM seems to favor a higher release of enzymes, likely due to a nutrient-deficient condition [
27]. The effect of pH-controlled batch cultures is also evidenced in
Figure 5a, showing that the higher the pH of the CM adjusted in the range of 5.8 to 7.0, the lower the microbial activity of
P. chrysosporium. A pH range of 4.0–4.5 has been reported to be more favorable for
P. chrysosporium to secrete ligninolytic enzymes [
13,
37]. Therefore, it is reasonable that higher pH values beyond pH 4.0 slowed down the enzyme activity, as indicated in
Figure 5a.
In the modified CM, a higher enzyme concentration was attained using a CM with no pH control, which led to 58 U/L LiP and 143 U/L MnP, followed by the controlled-pH culture adjusted to pH 5.8 with 34 U/L LiP and 78 U/L MnP. A pH-controlled culture conditioned at pH 7.0 likely resulted in the lowest enzyme activity, as enzymes were not detected on day 7 of incubation. In the standard CM, a similar trend was found with the highest enzyme activity of 51 U/L LiP and 128 U/L MnP using no pH control. The lower concentration of enzymes with increasing pH is presumably attributed to the fact that a pH environment outside of the range of 4.0–4.5 might have affected enzyme stability, remarkably leading to a decreased concentration of enzymes [
38]. pH is crucial for the stability and activity of heme-containing enzymes such as LiP and MnP, affecting their ionic form of the active sites and, consequently, the binding with the heme group [
39]. Another factor to consider is the higher concentration of phosphate salts over 4 g/L used to maintain a pH > 6.0, as an excessive concentration of phosphate salts could have limited the optimal growth and metabolism [
40].
Considering
Figure 5b, the comparison of sulfide oxidation at different pH-controlled environments reveals that pH-controlled cultures at pH > 5.8 induced a lower sulfide oxidation compared with no control experiments, which principally seems to be interconnected with the reduced activity of enzymes at elevated pHs. Using the modified CM conditions, 28.7% sulfide oxidation could be achieved by utilizing no pH control. The second-best sulfide oxidation was 14.3%, attained in a pH-controlled culture at pH 5.8, followed by 13.2% adjusted the culture to pH 6.0. Successively, the higher pH adjustment of the CM led to lower fungal activity and, consequently, lower sulfide oxidation, reaching only 5.1% and 1.6% while using the modified CM at pH 6.5 and pH 7.0, respectively. Sulfide oxidation in standard CM followed a comparable tendency at increasing pH values, as shown in
Figure 5b.
3.5. Evaluation of Corn Steep as a CM Component
Taking into consideration that the proposed investigated CM resulted in partial sulfide oxidation after 14 d, corn steep, a waste medium ingredient, was used as an alternative to increasing the viability of the process from an economic perspective. Thus, the addition of corn steep in the standard CM was investigated as a possible replacement for other nourishing components, including yeast extract and malt extract. Adding corn steep changed the initial pH solution of the CM to moderately acidic pH (pH 4). Therefore, the pH of all solutions was adjusted to pH 5.8 before inoculation for better comparison with the previous results, as
P. chrysosporium responds differently to different pH environments [
13,
28].
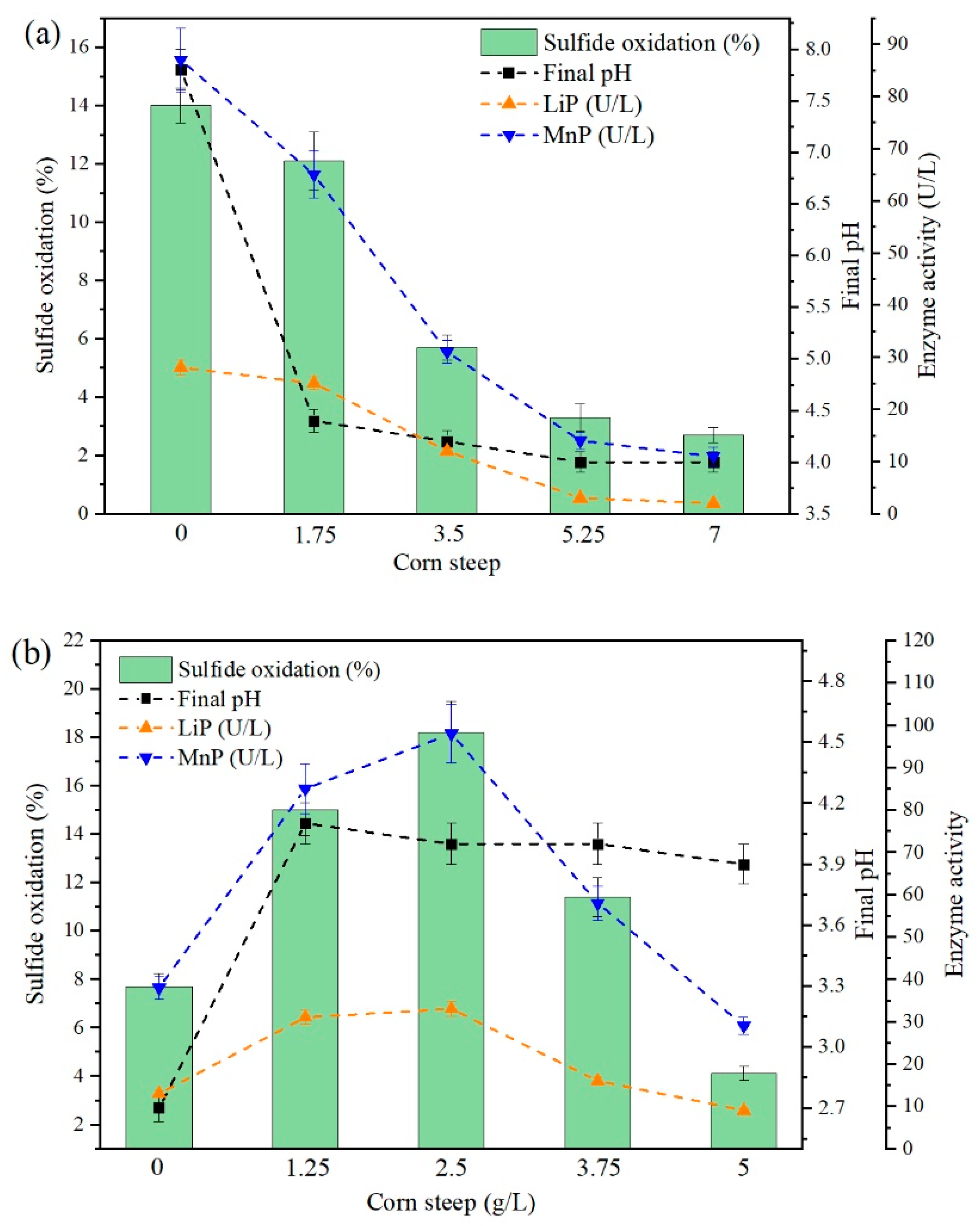
Figure 6 shows the effect of corn steep in the standard CM in terms of enzyme activity and sulfide oxidation by replacing the corresponding concentration of malt extract (0–7 g/L) and yeast extract (0–5 g/L). As shown in
Figure 6a, the substitution of malt extract for corn steep impacted microbial activity and sulfide oxidation, showing a decreasing trend for higher concentrations of corn steep in the CM in the range of 0–7 g /L. By replacing yeast extract for corn steep (
Figure 6b), the addition of corn steep displayed a good contribution to substituting yeast extract at increasing concentrations up to 2.5 g/L, attaining a maximum of 18.2% sulfide oxidation and 33 U/L LiP and 98 U/L MnP. Concentrations above 2.5 g/L seem to affect enzyme secretion and sulfide oxidation, leading to 9 U/L LiP and 29 U/L MnP with 4.1% sulfide oxidation with 5 g/L corn steep in the CM. The final pH in all culture media having corn steep was close to pH 4, which seems to be attributed to the composition of corn steep based on lactic acid [
41].
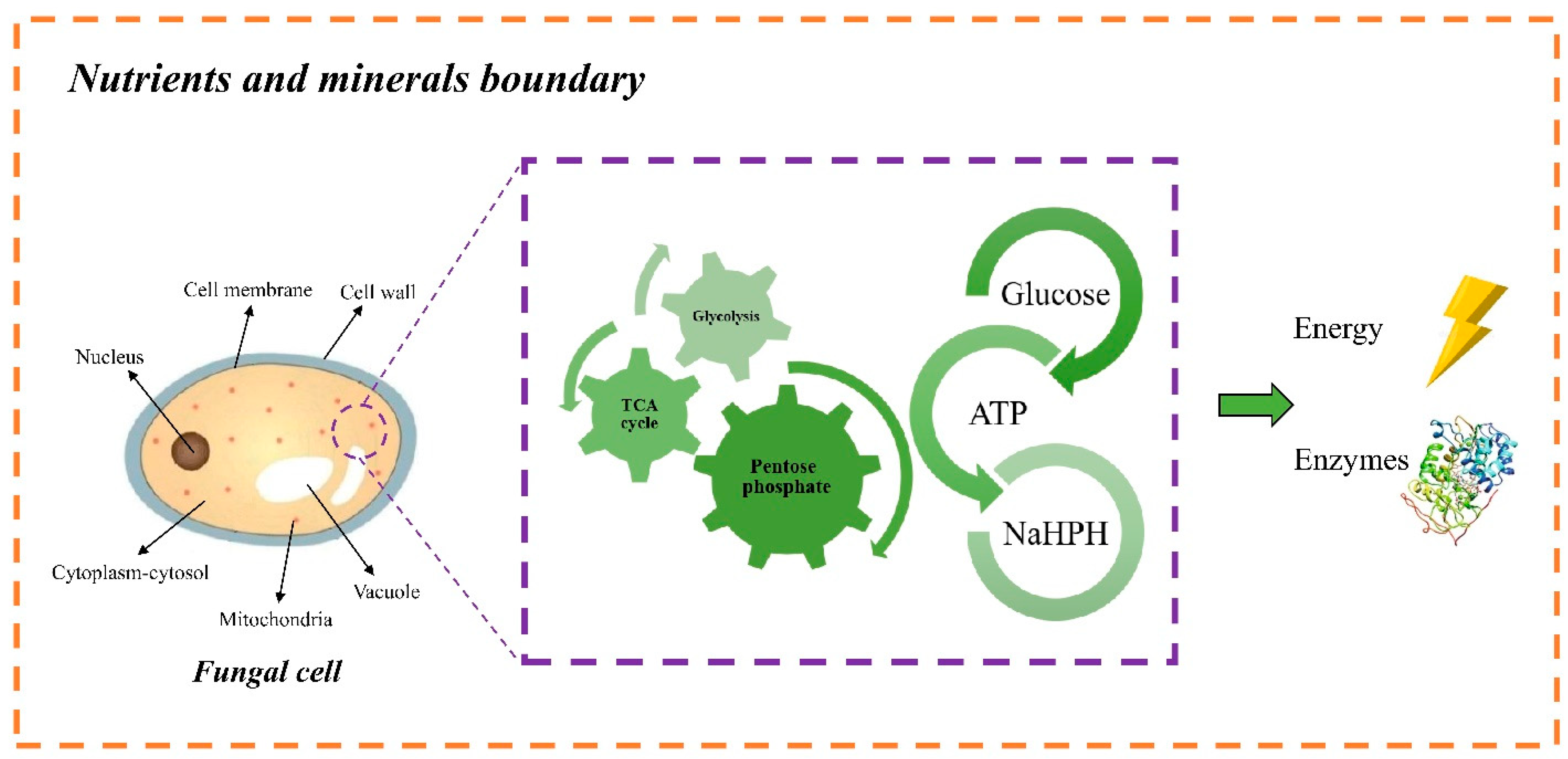
In the cells of
P. chrysosporium and other white-rot fungi, a wide variety of metabolic pathways take place simultaneously, e.g., glycolysis, pentose-phosphate pathway, tricarboxylic acid (TCA) cycle, which are all essential for the growth and functionality of fungal cells. Glycolytic pathways, together with the TCA cycle, produce the oxidation of carbohydrates, fatty acids, and amino acids, forming high-energy molecules, e.g., adenosine triphosphate (ATP) and nicotinamide adenine dinucleotide (NADPH), which support the formation of proteins and act as energy storage for the fungal cells [
42].
Figure 7 shows the role of the principal metabolic pathways in
P. chrysosporium cells. The gene expression for different types of enzymes, including LiP and MnP, as well as the intracellular and extracellular interactions, are highly influenced by the type of growth substrates and environmental conditions to which the microorganism is exposed in the CM [
43].
Enzyme activity for both LiP and MnP was detected in all the samples. Nonetheless, the enzyme concentration was lower when malt extract was replaced by corn steep at concentrations ≥1.75 g/L (2 U/L LiP and 11 U/L MnP), and yeast extract was substituted for corn steep at concentration ≥3.75 g/L (9 U/L LiP and 29 U/L). The maximum enzyme concentration (33 U/L LiP and 98 U/L MnP) was produced when yeast extract was substituted by corn steep in the CM in the range of 1.25 to 2.5 g/L, reaching a sulfide oxidation of 18.2% after 14 d bio-oxidation. The general trends can be interpreted by analyzing the general composition of each particular component in the CM. About 90% of malt extract is composed of reduced sugars, including but not limited to maltose, fructose, and sucrose, with a lower proportion of nitrogenous substrate, e.g., amino acids and peptides. On the other hand, yeast extract is a good source of N and other essential micronutrients for the better functioning of fungal cells comprising vitamins (biotin and vitamin B), TE, as well as proteins, free amino acids, and peptides. The nourishing composition of yeast extract makes it a valuable medium ingredient.
Based on the composition of corn steep, this substrate is also considered a source of N with nearly 7.7–8.2% of total N [
44,
45]. Similarly to yeast extract, corn steep is also a source of TE and vitamins. Additionally, it has a significant content of lactic acid, representing 10–30% of the dry mass and sugars, including glucose and fructose (below 5%) [
41]. Hence, corn steep is a substrate with a composition similar to yeast extract, providing N sources, vitamins, and TE required to sustain the fungal growth and metabolism, resulting in comparable outcomes when not supplying yeast extract in the CM. Consequently, corn steep could be considered to replace yeast extract; however, it resulted in an unsatisfactory response as a replacement of malt extract.
3.6. CM Comparison
The modified concentrations of reagents to attain maximal sulfide oxidation found in prior sections (12.86 g/L glucose, 2.20 g/L malt extract, 1.67 g/L yeast extract, and 0.49 g/L MgSO
4·7H
2O) were utilized as a reference for the substitution of yeast extract for corn steep in subsequent bio-oxidation experiments. In the modified CM containing yeast extract, enzyme activity was 58 U/L LiP and 143 U/L MnP after 7 d of growth, and sulfide oxidation after 14 d bio-oxidation was 28.7%. When replacing yeast extract for corn steep in the modified CM, enzyme activity resulted in 39 U/L LiP and 116 U/L MnP, and sulfide oxidation was 21.6% following 14 d of bio-oxidation, as shown in
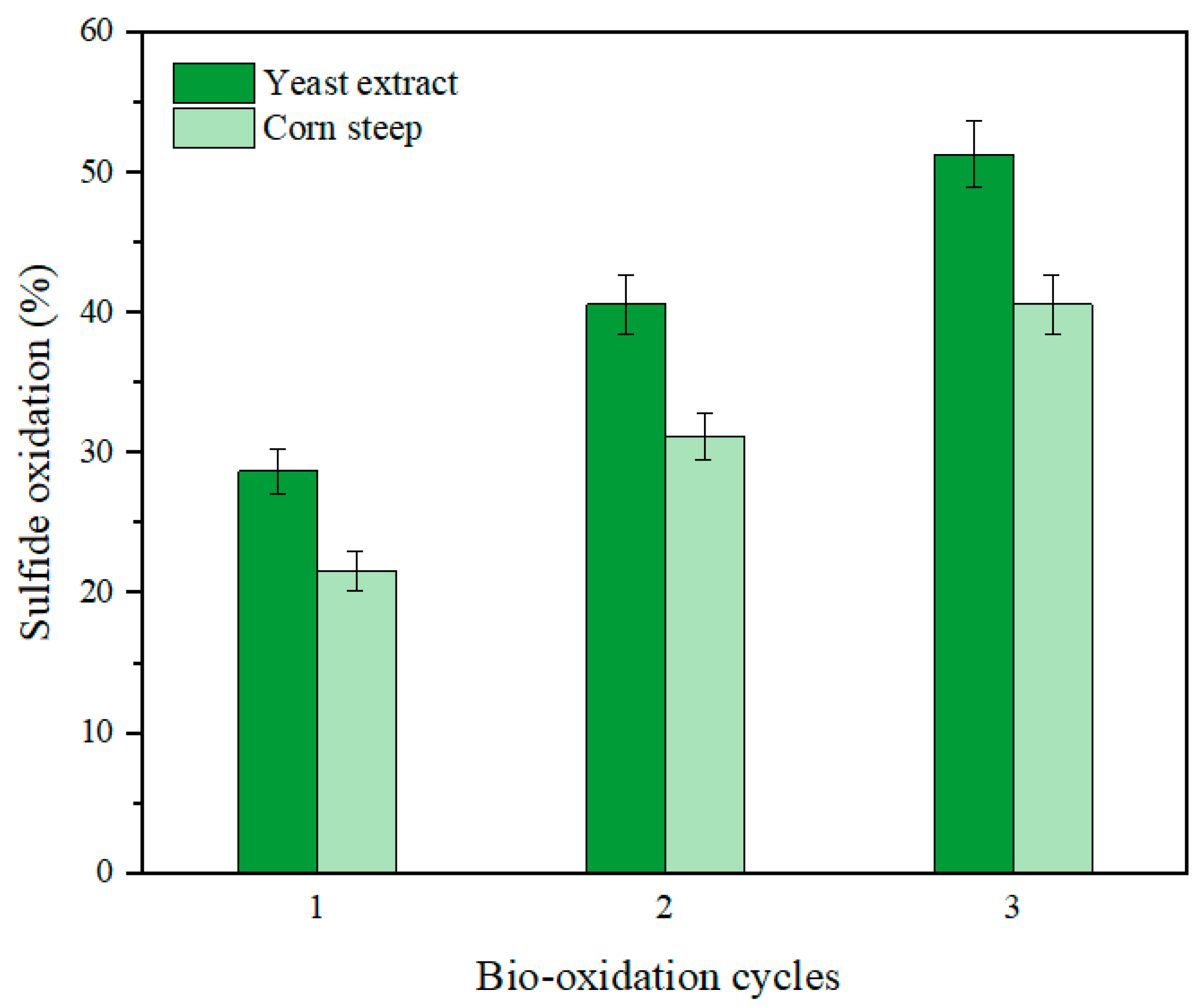
Figure 8.
Since sulfide oxidation stalled after 14 d of bio-oxidation with no considerable increase after extended reaction time, a replenishing strategy was used to further compare both CM in terms of sulfide oxidation as well as Au recovery [
6]. A comparison between the modified CM having equal concentration of yeast extract and corn steep is displayed in
Figure 8., showing sulfide oxidation after three cycles of 14-d bio-oxidation. Results revealed that sulfide oxidation gradually increased with the number of replenishing cycles up to 51.1% and 40.6% in the modified CM containing yeast extract and corn steep, respectively.
Table 4 shows the Au recovery obtained from the oxidized ore after three cycles of bio-oxidation for a total of 42 d, resulting in 52.1% and 43.8% using the modified CM having yeast extract and corn steep, correspondingly.
Comparing the results using yeast extract and corn steep in the modified CM, the former led to a slightly higher Au recovery of ca. 52.1%, whilst the latter could represent a more economical alternative. Corn steep is a waste product originating from corn industry plants and thus could be considered as a low-cost replacement ingredient for the CM of
P. chrysosporium. When comparing the cost of reagent in bulk, yeast extract turns out to be 16 times more expensive on average, considering the same mass of corn steep, as shown in
Table 4.
Due to the similar results in sulfide oxidation and Au recovery obtained in both CMs, it might be worth exchanging a slightly higher Au recovery for a more economical operational cost, considering low-budget reagents in replenishing bio-oxidation. Still, it is worth noting that sulfide oxidation <30% after one cycle of bio-oxidation is not significantly comparable to the conventional bio-oxidation process with acidophilic bacteria. Nevertheless, it should be noted that this research, for the first time, investigates the use of corn steep in bio-oxidation of low-grade refractory Au ores, and therefore, the process needs further research. There could still be other additional factors for further development to improve the process, such as replacing other components for waste products, e.g., glucose for molasses, and utilizing chemostat-like reactors with multiple sensors to keep an optimum level for nutrients, pH as well as metabolites, which could maximize sulfide oxidation and Au recovery in future research.
3.7. Implications and Limitations
This study shows a dependency of the fungus
P. chrysosporium on CM conditions, including mainly the nutrient availability and pH solution >5.8, leading to particular microbial activities and, subsequently, different trends in sulfide oxidation. Consequently, this research highlights that both a limitation and an excess of nutrients can restrict the performance of this microorganism and, therefore, sulfide oxidation. An adequate concentration of reagents is essential in bio-oxidation experiences, especially glucose, malt extract, and yeast extract, which were more relevant for the fungal activity. Another important implication of this investigation was the potential of corn steep waste to support microbial growth by serving as a source of micro- and macro-nutrients. This implication allows evaluating other waste or less refined products, e.g., molasses, to find a more cost-effective option to oxidize sulfidic ores. From an economic perspective, a full operation using waste products as a source of nutrients to grow the microorganism could considerably decrease the cost associated with the CM, increasing the viability of the process compared to the standard CM option. The use of waste containing CM would further strengthen the circularity of waste management, contributing to a decrease in the accumulation of municipal solid waste and global greenhouse emissions worldwide [
23].
The current investigation had certain limitations. Although corn steep waste supported the microbial activity of the fungus
P. chrysosporium to oxidize a sulfidic Au ore, the pH of the solution remained acidic throughout the tests. Likely, the composition of corn steep based on 10 to 30%
w/
w lactic acid could be the principal responsible for that pH trend compared to the yeast extract-bearing CM. In addition, the performance of the fungi still needs to be further strengthened to improve the bio-oxidation of sulfide ores compared with the conventional option assisted by acidophilic bacteria. The concentration of metabolites is a limitation that might also have restricted further oxidation. Therefore, more research should be done in that direction by considering bioengineered microorganisms that would improve the fungal activity under the studied experimental conditions [
4]. The increase in the concentration of metabolites at different pH ranges using current and new reagents, especially waste products, considering chemostat-like reactors, should also be evaluated to overcome these limitations.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}