
The Eco-Evolutionary Imperative: Revisiting Weed Management in the Midst of an Herbicide Resistance Crisis

 ,
,
Abstract
:1. Eco-Evolutionary Thinking and Weed Management
2. Weed Science and Management: What Went Wrong?
3. How Did We Get Here? A Brief History of Pest Management
4. The Doomed Marriage between Integrated Weed Management and Herbicide Resistance
5. Eco-Evolutionary Thinking and Weed Management
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dobzhansky, T. Nothing in biology makes sense except in the light of evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species by Natural Selection; Murray: London, UK, 1859. [Google Scholar]
- Antolin, M.F.; Herbers, J.M. Perspective: Evolution’s struggle for existence in America’s public schools. Evolution 2001, 55, 2379–2388. [Google Scholar] [CrossRef] [PubMed]
- Thrall, P.H.; Oakeshott, J.G.; Fitt, G.; Southerton, S.; Burdon, J.J.; Sheppard, A.; Russell, R.J.; Zalucki, M.; Heino, M.; Denison, R.F. Evolution in agriculture: The application of evolutionary approaches to the management of biotic interactions in agro-ecosystems. Evol. Appl. 2011, 4, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Neve, P.; Vila-Aiub, M.; Roux, F. Evolutionary-thinking in agricultural weed management. New Phytol. 2009, 184, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Neve, P.; Busi, R.; Renton, M.; Vila-Aiub, M.M. Expanding the eco-evolutionary context of herbicide resistance research. Pest Manag. Sci. 2014, 70, 1385–1393. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.; Kinnison, M.T.; Bernatchez, L. In light of evolution: Interdisciplinary challenges in food, health, and the environment. Evol. Appl. 2011, 4, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.P.; Jorgensen, P.S.; Kinnison, M.T.; Bergstrom, C.T.; Denison, R.F.; Gluckman, P.; Smith, T.B.; Strauss, S.Y.; Tabashnik, B.E. Applying evolutionary biology to address global challenges. Science 2014, 346, 1245993. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, G.E. The Ecological Theater and the Evolutionary Play; Yale University Press: New Haven, CT, USA, 1965. [Google Scholar]
- Ellner, S.P. Rapid evolution: From genes to communities, and back again? Funct. Ecol. 2013, 27, 1087–1099. [Google Scholar] [CrossRef]
- Hairston, N.G.; Ellner, S.P.; Geber, M.A.; Yoshida, T.; Fox, J.A. Rapid evolution and the convergence of ecological and evolutionary time. Ecol. Lett. 2005, 8, 1114–1127. [Google Scholar] [CrossRef]
- Hendry, A.P.; Kinnison, M.T. Perspective: The pace of modern life: Measuring rates of contemporary microevolution. Evolution 1999, 53, 1637–1653. [Google Scholar] [CrossRef]
- Reznick, D.A.; Bryga, H.; Endler, J.A. Experimentally induced life-history evolution in a natural population. Nature 1990, 346, 357–359. [Google Scholar] [CrossRef]
- Grant, P.R.; Grant, B.R. Unpredictable evolution in a 30-year study of Darwin’s finches. Science 2002, 296, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Antonovics, J. The input from population genetics: “The new ecological genetics”. Syst. Bot. 1976, 1, 233–245. [Google Scholar] [CrossRef]
- Pimentel, D. Animal population regulation by genetic feedback mechanism. Am. Nat. 1961, 95, 65–79. [Google Scholar] [CrossRef]
- Elderd, B.D.; Dushoff, J.; Dwyer, G. Host-Pathogen Interactions, Insect outbreaks, and natural selection for disease resistance. Am. Nat. 2008, 172, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.A.; Sivars-Becker, L. Rapid evolution and ecological host-parasite dynamics. Ecol. Lett. 2007, 10, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Ellner, S.P.; Jones, L.E.; Bohannan, B.J.M.; Lenski, R.E.; Hairston, N.G., Jr. Cryptic population dynamics: Rapid evolution masks trophic interactions. PLoS Biol. 2007, 5, 1868–1879. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Jones, L.E.; Ellner, S.P.; Fussmann, G.F.; Hairston, N.G. Rapid evolution drives ecological dynamics in a predator-prey system. Nature 2003, 424, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Abrams, P.A.; Matsuda, H. Prey adaptation as a cause of predator-prey cycles. Evolution 1997, 51, 1742–1750. [Google Scholar] [CrossRef]
- Schreiber, S.J.; Buerger, R.; Bolnick, D.I. The community effects of phenotypic and genetic variation within a predator population. Ecology 2011, 92, 1582–1593. [Google Scholar] [CrossRef] [PubMed]
- Fussmann, G.F.; Loreau, M.; Abrams, P.A. Eco-evolutionary dynamics of communities and ecosystems. Funct. Ecol. 2007, 21, 465–477. [Google Scholar] [CrossRef]
- Coulson, T.; Benton, T.G.; Lundberg, P.; Dall, S.R.X.; Kendall, B.E. Putting evolutionary biology back in the ecological theatre: A demographic framework mapping genes to communities. Evol. Ecol. Res. 2006, 8, 1155–1171. [Google Scholar]
- Nesse, R.M.; Bergstrom, C.T.; Ellison, P.T.; Flier, J.S.; Gluckman, P.; Govindaraju, D.R.; Niethammer, D.; Omenn, G.S.; Perlman, R.L.; Schwartz, M.D.; et al. Making evolutionary biology a basic science for medicine. Proc. Nat. Acad. Sci. USA 2010, 107, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Hendry, A.P.; Kinnison, M.T.; Heino, M.; Day, T.; Smith, T.B.; Fitt, G.; Bergstrom, C.T.; Oakeshott, J.; Jorgensen, P.S.; Zalucki, M.P.; et al. Evolutionary principles and their practical application. Evol. Appl. 2011, 4, 159–183. [Google Scholar] [CrossRef] [PubMed]
- Schoener, T.W. The newest synthesis: Understanding the interplay of evolutionary and ecological dynamics. Science 2011, 331, 426. [Google Scholar] [CrossRef] [PubMed]
- Loeuille, N.; Barot, S.; Georgelin, E.; Kylafis, G.; Lavigne, C. Eco-evolutionary dynamics of agricultural networks: Implications for sustainable management. Adv. Ecol. Res. 2013, 49, 339–435. [Google Scholar]
- Keren, I.N.; Menalled, F.D.; Weaver, D.K.; Robison-Cox, J.F. Interacting agricultural pests and their effect on crop yield: Application of a Bayesian decision theory approach to the joint management of Bromus tectorum and Cephus cinctus. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Zimdahl, R.L. Weed Science—A Plea for Thought-Revisited; Springer Brief in Agriculture: Dordrecht, The Netherlands, 2012; p. 73. [Google Scholar]
- Grube, A.; Donaldson, D.; Kiely, T.; Wu, L. Pesticides Industry Sales and Usage–2006 and 2007 Market Estimates; U.S. Environmental Protection Agency: Washington, DC, USA, 2011. Available online: https://nepis.epa.gov/Exe/ZyPDF.cgi/P100A8DN.PDF?Dockey=P100A8DN.PDF#_ga=1.47126055.1632740713.1445374245 (accessed on 13 October 2016).
- North American Spine Society (NASS). Agricultural Chemical Use Program. United States Department of Agriculture. Available online: http://www.nass.usda.gov/Surveys/Guide_to_NASS_Surveys/Chemical_Use/index.php (accessed on 11 October 2016).
- Shaner, D.L.; Beckie, H.J. The future for weed control and technology. Pest Manag. Sci. 2014, 70, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Zimdahl, R.L. Who are you and where are you going. Weed Tech. 1994, 8, 388–391. [Google Scholar]
- Lewis, W.J.; vanLenteren, J.C.; Phatak, S.C.; Tumlinson, J.H. A total system approach to sustainable pest management. Proc. Nat. Acad. Sci. USA 1997, 94, 12243–12248. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, D.A.; Bastiaans, L.; Sattin, M. The role of ecology in the development of weed management systems: An outlook. Weed Res. 2000, 40, 49–62. [Google Scholar] [CrossRef]
- Jasieniuk, M.; Brûlé-Babel, A.L.; Morrison, I.N. The evolution and genetics of herbicide resistance in weeds. Weed Sci. 1996, 44, 176–193. [Google Scholar] [CrossRef]
- Heap, I. International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.com/summary/home.aspx (accessed on 13 October 2016).
- Hall, L.M.; Holtum, J.A.M.; Powles, S.B. Mechanisms responsible for cross resistance and multiple resistance. In Herbicide Resistance in Plants: Biology and Biochemistry; Powles, S.B., Holtum, J.A.M., Eds.; Lewis Publishers: Boca Raton, FL, USA, 1994; pp. 243–261. [Google Scholar]
- Lehnhoff, E.A.; Keith, B.K.; Dyer, W.E.; Peterson, R.K.; Menalled, F. Multiple herbicide resistance in wild oat and impacts on physiology, germinability, and seed production. Agron. J. 2013, 105, 854–862. [Google Scholar] [CrossRef]
- Stoate, C.; Boatman, N.D.; Borralho, R.J.; Carvalho, C.R.; de Snoo, G.R.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef]
- Duke, S.O. Why have no new herbicide modes of action appeared in recent years? Pest Manag. Sci. 2012, 68, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Stern, V.M.; Smith, R.; van Bosch, R.; Hagen, K.S. The integration of chemical and biological control of the spotted alfalfa aphid. The integrated control concept. Field experiments on the effects of insecticides. Impact of commercial insecticide treatments. Hilgardia 1959, 29, 81–101. [Google Scholar] [CrossRef]
- Geier, P.W. Management of insect pests. Ann. Rev. Entomol. 1966, 11, 471–490. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.F. Integration of biological and chemical control. Introduction and principles. Bull. Entomol. Soc. Am. 1962, 8, 188–189. [Google Scholar] [CrossRef]
- Pedigo, L.P. Entomology and Pest Management; Macmillan Publishing Company: New York, NY, USA, 1989. [Google Scholar]
- Higley, L.G.; Wintersteen, W.K. Thresholds and environmental quality. In Economic Thresholds for Integrated Pest Management; Higley, L., Pedigo, L., Eds.; University of Nebraska Press: Lincoln, Nebraska, 1996; pp. 249–274. [Google Scholar]
- Kogan, M. Integrated pest management: Historical perspectives and contemporary developments. Ann. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.R. Symposium on integrated weed management systems. Technology for crop production and protection held at the Annual Meeting of Weed Science Society of America. Las Vegas, Nevada, USA FEB. 19 1981. Weed Sci. 1982, 30 (Suppl. S1), 2. [Google Scholar]
- Swanton, C.J.; Weise, S.F. Integrated weed management. The rationale and approach. Weed Tech. 1991, 5, 657–663. [Google Scholar]
- Owen, M.D. K.; Beckie, H.J.; Leeson, J.Y.; Norsworthy, J.K.; Steckel, L.E. Integrated pest management and weed management in the United States and Canada. Pest Manag. Sci. 2015, 71, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Liebman, M.; Gallandt, E.R. Many little hammers: Ecological management of crop-weed interactions. In Ecology in Agriculture; Jackson, L.E., Ed.; Academic: San Diego, CA, USA, 1997; pp. 291–343. [Google Scholar]
- Norsworthy, J.K.; Ward, S.M.; Shaw, D.R.; Llewellyn, R.S.; Nichols, R.L.; Webster, T.M.; Bradley, K.W.; Frisvold, G.; Powles, S.B.; Burgos, N.R.; et al. Reducing the Risks of Herbicide Resistance: Best Management Practices and Recommendations. Weed Sci. 2012, 60, 31–62. [Google Scholar] [CrossRef]
- Weed Science Society of America (WSSA). Available online: http://wssa.net/about-us/ (accessed on 8 December 2016).
- Weed Science Society of America (WSSA). WSSA Infographics on Herbicide Resistance Management. Available online: http://wssa.net/herbicide-resistance/wssa-infographics-on-herbicide-resistance-management/ (accessed on 8 December 2016).
- Owen, M.D.K. Diverse approaches to herbicide-resistant weed management. Weed Sci. 2016, 64, 570–584. [Google Scholar] [CrossRef]
- Liebman, M.; Mohler, C.L.; Staver, C.P. Ecological Management of Agricultural Weeds; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Delye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the evolution of herbicide resistance in weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.S.; Dixon, P.M.; Liebman, M. Using matrix models to determine cropping system effects on annual weed demography. Ecol. Appl. 2004, 14, 655–668. [Google Scholar] [CrossRef]
- Westerman, P.; Liebman, M.; Menalled, F.D.; Heggenstaller, A.H.; Hartzler, R.G.; Dixon, P.M. Are many little hammers effective?—Velvetleaf (Abutilon theophrasti) population dynamics in two- and four-year crop rotation systems. Weed Sci. 2005, 53, 382–392. [Google Scholar] [CrossRef]
- Davis, P.B.; Menalled, F.D.; Peterson, R.K.D.; Maxwell, B.D. Refinement of weed risk assessments for biofuels using Camelina sativa as a model species. J. Appl. Ecol. 2011, 48, 989–997. [Google Scholar] [CrossRef]
- VanGessel, M.J. Glyphosate-resistant horseweed from Delaware. Weed Sci. 2001, 49, 703–705. [Google Scholar] [CrossRef]
- Dauer, J.T.; Mortensen, D.A.; Vangessel, M.J. Temporal and spatial dynamics of long-distance Conyza canadensis seed dispersal. J. Appl. Ecol. 2007, 44, 105–114. [Google Scholar] [CrossRef]
- Soteres, J.K.; Peterson, M.A. Industry views of monitoring and mitigation of herbicide resistance. Weed Sci. 2015, 63, 972–975. [Google Scholar] [CrossRef]
- Beckie, H.J.; Reboud, X. Selecting for weed resistance: Herbicide rotation and mixture. Weed Tech. 2009, 23, 363–370. [Google Scholar] [CrossRef]
- Evans, J.A.; Tranel, P.J.; Hager, A.G.; Schutte, B.; Wu, C.; Chatham, L.A.; Davis, A.S. Managing the evolution of herbicide resistance. Pest Manag. Sci. 2016, 72, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Neve, P.; Sadler, J.; Powles, S.B. Multiple herbicide resistance in a glyphosate-resistant rigid ryegrass (Lolium rigidum) population. Weed Sci. 2004, 52, 920–928. [Google Scholar] [CrossRef]
- Duke, S.O. Perspectives on transgenic, herbicide-resistant crops in the United States almost 20 years after introduction. Pest Manag. Sci. 2015, 71, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, L.; Coppola, G.; Zelasco, S. New technologies for insect-resistant and herbicide-tolerant plants. Trends Biotech. 2016, 34, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, D.A.; Egan, J.F.; Maxwell, B.D.; Ryan, M.R.; Smith, R.G. Navigating a Critical Juncture for Sustainable Weed Management. Bioscience 2012, 62, 75–84. [Google Scholar] [CrossRef]
- Egan, J.F.; Maxwell, B.D.; Mortensen, D.A.; Ryan, M.R.; Smith, R.G. 2,4-Dichlorophenoxyacetic acid (2,4-D)-resistant crops and the potential for evolution of 2,4-D-resistant weeds. Proc. Natl. Acad. Sci. USA 2011, 108, E37. [Google Scholar] [CrossRef] [PubMed]
- Landis, D.A.; Menalled, F.D.; Costamagna, A.C.; Wilkinson, T.K. Manipulating plant resources to enhance beneficial arthropods in agricultural landscapes. Weed Sci. 2005, 53, 902–908. [Google Scholar] [CrossRef]
- Smith, R.G.; Menalled, F.D.; Robertson, G.P. Temporal yield variability under conventional and alternative management systems. Agron. J. 2007, 99, 1629–1634. [Google Scholar] [CrossRef]
- Smith, R.G. A succession-energy framework for reducing non-target impacts of annual crop production. Agric. Syst. 2015, 133, 14–21. [Google Scholar] [CrossRef]
- Baucom, R.S. The remarkable repeated evolution of herbicide resistance. Am. J. Bot. 2016, 103, 181–183. [Google Scholar] [CrossRef] [PubMed]
- Booth, B.D.; Swanton, C.J. Assembly theory applied to weed communities. Weed Sci. 2002, 50, 2–13. [Google Scholar] [CrossRef]
- Petit, S.; Alignier, A.; Colbach, N.; Joannon, A.; Le Coeur, D.; Thenail, C. Weed dispersal by farming at various spatial scales. A review. Agric. Sustain. Dev. 2013, 33, 205–217. [Google Scholar] [CrossRef]
- Maxwell, B.D.; Mortimer, A.M. Selection for herbicide resistance. Herbic. Resist. Plants Biol. Biochem. 1994, 1, 205–212. [Google Scholar]
- Hoffmann, A.A.; Merila, J. Heritable variation and evolution under favourable and unfavourable conditions. Trends Ecol. Evol. 1999, 14, 96–101. [Google Scholar] [CrossRef]
- McGuigan, K.; Sgro, C.M. Evolutionary consequences of cryptic genetic variation. Trends Ecol. Evol. 2009, 24, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, B.D.; O’Donovan, J.T. Understanding weed-crop interactions to manage weed problems. In Non Chemical Weed Management: Principles, Concepts and Technology; CAB International/North America Office: Cambridge, MA, USA, 2007; pp. 17–33. [Google Scholar]
- Weiner, J.; Andersen, S.B.; Wille, W.K.M.; Griepentrog, H.W.; Olsen, J.M. Evolutionary Agroecology: The potential for cooperative, high density, weed-suppressing cereals. Evol. Appl. 2010, 3, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, B.D. Weed thresholds—The space component and considerations for herbicide resistance. Weed Tech. 1992, 6, 205–212. [Google Scholar]
- Warner, K.D. Extending agroecology: Grower participation in partnerships is key to social learning. Renew. Agric. Food Sys. 2006, 21, 84–94. [Google Scholar] [CrossRef]
- Ervin, D.; Jussaume, R. Integrating social science into managing herbicide-resistant weeds and associated environmental impacts. Weed Sci. 2014, 62, 403–414. [Google Scholar] [CrossRef]
- Dauer, J.T.; Luschei, E.C.; Mortensen, D.A. Effects of landscape composition on spread of an herbicide-resistant weed. Landsc. Ecol. 2009, 24, 735–747. [Google Scholar] [CrossRef]
- Dauer, J.T.; Mortensen, D.A.; Luschei, E.C.; Isard, S.A.; Shields, E.; Van-Gessel, M.J. Conyza canadensis seed ascent in the lower atmosphere. Agric. For. Meteorol. 2009, 149, 526–534. [Google Scholar] [CrossRef]
- Okada, M.; Hanson, B.D.; Hembree, K.J.; Peng, Y.; Shrestha, A.; Stewart, C.N., Jr.; Wright, S.D.; Jasieniuk, M. Evolution and spread of glyphosate resistance in Conyza canadensis in California. Evol. Appl. 2013, 6, 761–777. [Google Scholar] [CrossRef]
- National Research Council. Risk Assessment in the Federal Government: Managing the Process; National Academy Press: Washington, DC, USA, 1983. [Google Scholar]
- National Research Council. Understanding Risk: Informing Decisions in a Democratic Society; National Academy Press: Washington, DC, USA, 1996. [Google Scholar]
- Intergovernmental Panel on Climate Change. International Standards for Phytosanitary Management (ISPM) 2: Framework for Pest Risk Analysis; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; p. 16. [Google Scholar]
- Pheloung, P.C.; Williams, P.A.; Halloy, S.R. A weed risk assessment model for use as a biosecurity tool evaluating plant introductions. J. Environ. Manag. 1999, 57, 239–251. [Google Scholar] [CrossRef]
- Schleier, J.J., III; Sing, S.E.; Peterson, R.K.D. Regional ecological risk assessment for the introduction of Gambusia affinis (western mosquitofish) into Montana watersheds. Biol. Invasions 2008, 10, 1277–1287. [Google Scholar] [CrossRef]
- Sing, S.E.; Peterson, R.K.D. Assessing environmental risks for established invasive weeds: Dalmatian (Linaria dalmatica) and yellow (L. vulgaris) toadflax in North America. Int. J. Environ. Res. Public Health 2011, 8, 2828–2853. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.S.; Peterson, R.K.D.; Macedo, P.A. An ecological risk assessment for insecticides used in adult mosquito management. Int. Environ. Assess. Manag. 2007, 3, 373–382. [Google Scholar] [CrossRef]
- Peterson, R.K.D. Comparing ecological risks of pesticides: The utility of a Risk Quotient ranking approach across refinements of exposure. Pest Manag. Sci. 2006, 62, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.K.D.; Hulting, A.N.G. A comparative ecological risk assessment for herbicides used on spring wheat: The effect of glyphosate when used within a glyphosate-tolerant wheat system. Weed Sci. 2004, 52, 834–844. [Google Scholar] [CrossRef]
- Stanton, R.A.; Pratley, J.E.; Hudson, D.; Dill, G.M. A risk calculator for glyphosate resistant in Lolium rigidum (Gaud.). Pest Manag. Sci. 2008, 64, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liebman, M.; Baraibar, B.; Buckley, Y.; Childs, D.; Christensen, S.; Cousens, R.; Eizenberg, H.; Heijting, S.; Loddo, D.; Merotto, A.; et al. Ecologically sustainable weed management: How do we get from proof-of-concept to adoption? Ecol. Appl. 2016, 26, 1352–1369. [Google Scholar] [CrossRef] [PubMed]


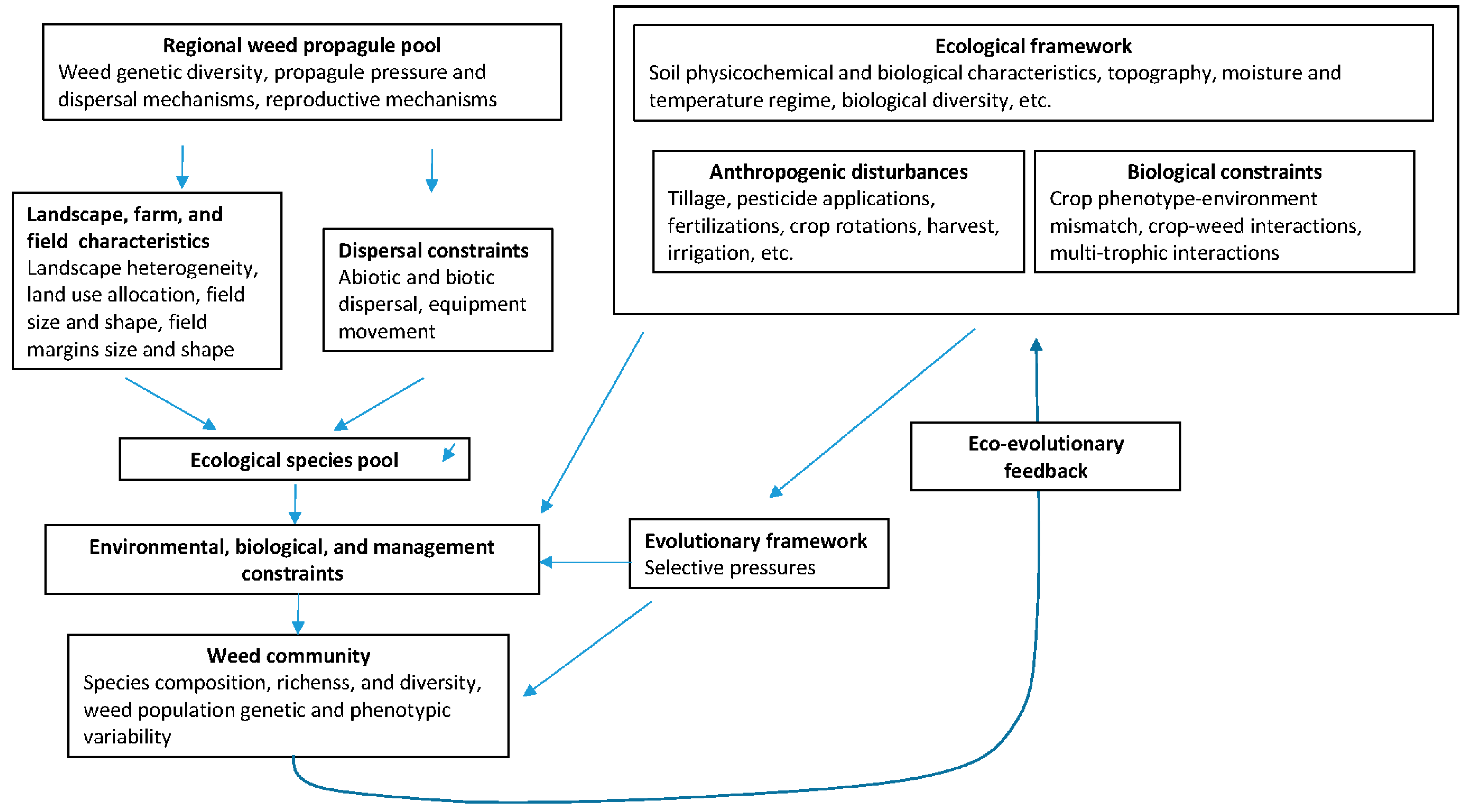{kind=link}
| Concepts | Actions |
|---|---|
| Spatiotemporal framework: evolution occurs at small spatial (within crop fields) and time scales (within season) | Maintain susceptible weed genotypes across the landscape. Augment the temporal and spatial variation of selection pressures by diversifying crop rotation and management tactics. Minimize the phenotype-environment mismatch of the cropping systems by using locally adapted and competitive crop varieties. |
| Genetic variability: the probability of a resistant mutant occurring in a population is directly related to its size | Complement reactive weed management practices with predictive tools. Incorporate intra- and inter-population genetic variability into demographic models. |
| Landscape and regional implications: the selection of herbicide resistant biotypes transcends any one farm | Since weed propagules can travel large distances either via nature (i.e., wind) or due to anthropogenic forces (contaminated grain and equipment), local networks of famers should coordinate actions to prevent the spread of herbicide resistant biotypes. USDA-APHIS and State Department of Agriculture must collaborate to develop state and federal weed laws and procedures that better monitor interstate movement of noxious and obnoxious weed seeds and vegetative propagules. Societal pressures together with local, state, or federal regulations should mandate the maximum use of specific herbicide active ingredients or a resistant crop trait within a region. Take advantage of genetic tool and dispersal models to design agricultural landscapes that minimize the spread of undesired genotypes. |
| Societal pressures, incentives, and regulations: incentives in the form of positive or negative reinforcement can help the adoption of eco-evolutionary practices | Registration of new crop biotechnologies that involve herbicides should explicitly address the management of herbicide resistance. Design programs to disincentive the overreliance on one specific technology package and encourage the adoption of ecologically based practices. Utilize fees collected from these programs to develop and deliver an ecological-based weed management program aimed at reducing herbicide selective pressures. Develop environmental payment structure/stewardship programs that connects IWM to broader environmental goals such as reducing the selective pressured towards undesired genotypes. Consider the suite of ecosystem functions provided by cropping systems when developing incentive policies, educational activities, and in developing research funding priorities for federal and state competitive research programs. |
| Risk assessment: utilize the paradigm of risk analysis to formally incorporate an eco-evolutionary perspective into IWM strategies | Jointly consider the inherent ability of the herbicide to select for resistance and the amount of contact between the herbicide and the weed population to assess the risk of resistance. Develop risk models that include weed phenotypic variation, density of weeds, total population of weeds, seedbank characteristics, treated area, frequency of application, landscape heterogeneity, and agronomic factors. |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menalled, F.D.; Peterson, R.K.D.; Smith, R.G.; Curran, W.S.; Páez, D.J.; Maxwell, B.D. The Eco-Evolutionary Imperative: Revisiting Weed Management in the Midst of an Herbicide Resistance Crisis. Sustainability 2016, 8, 1297. https://0-doi-org.brum.beds.ac.uk/10.3390/su8121297
Menalled FD, Peterson RKD, Smith RG, Curran WS, Páez DJ, Maxwell BD. The Eco-Evolutionary Imperative: Revisiting Weed Management in the Midst of an Herbicide Resistance Crisis. Sustainability. 2016; 8(12):1297. https://0-doi-org.brum.beds.ac.uk/10.3390/su8121297
Chicago/Turabian StyleMenalled, Fabian D., Robert K. D. Peterson, Richard G. Smith, William S. Curran, David J. Páez, and Bruce D. Maxwell. 2016. "The Eco-Evolutionary Imperative: Revisiting Weed Management in the Midst of an Herbicide Resistance Crisis" Sustainability 8, no. 12: 1297. https://0-doi-org.brum.beds.ac.uk/10.3390/su8121297