Nutrient Concentrations of Bush Bean (Phaseolus vulgaris L.) and Potato (Solanum tuberosum L.) Cultivated in Subarctic Soils Managed with Intercropping and Willow (Salix spp.) Agroforestry

Abstract
:1. Introduction
2. Materials and Methods
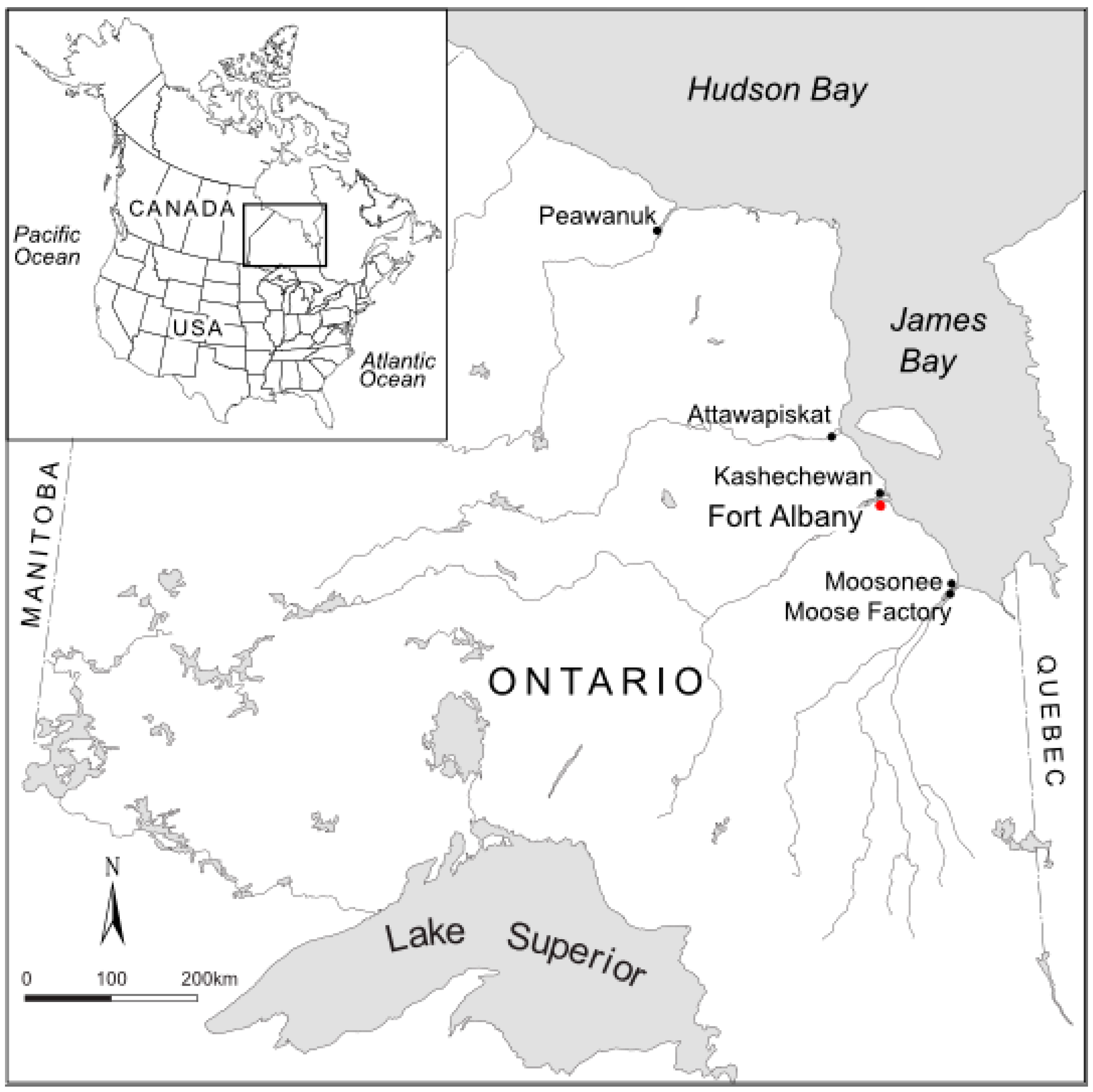
2.1. Location
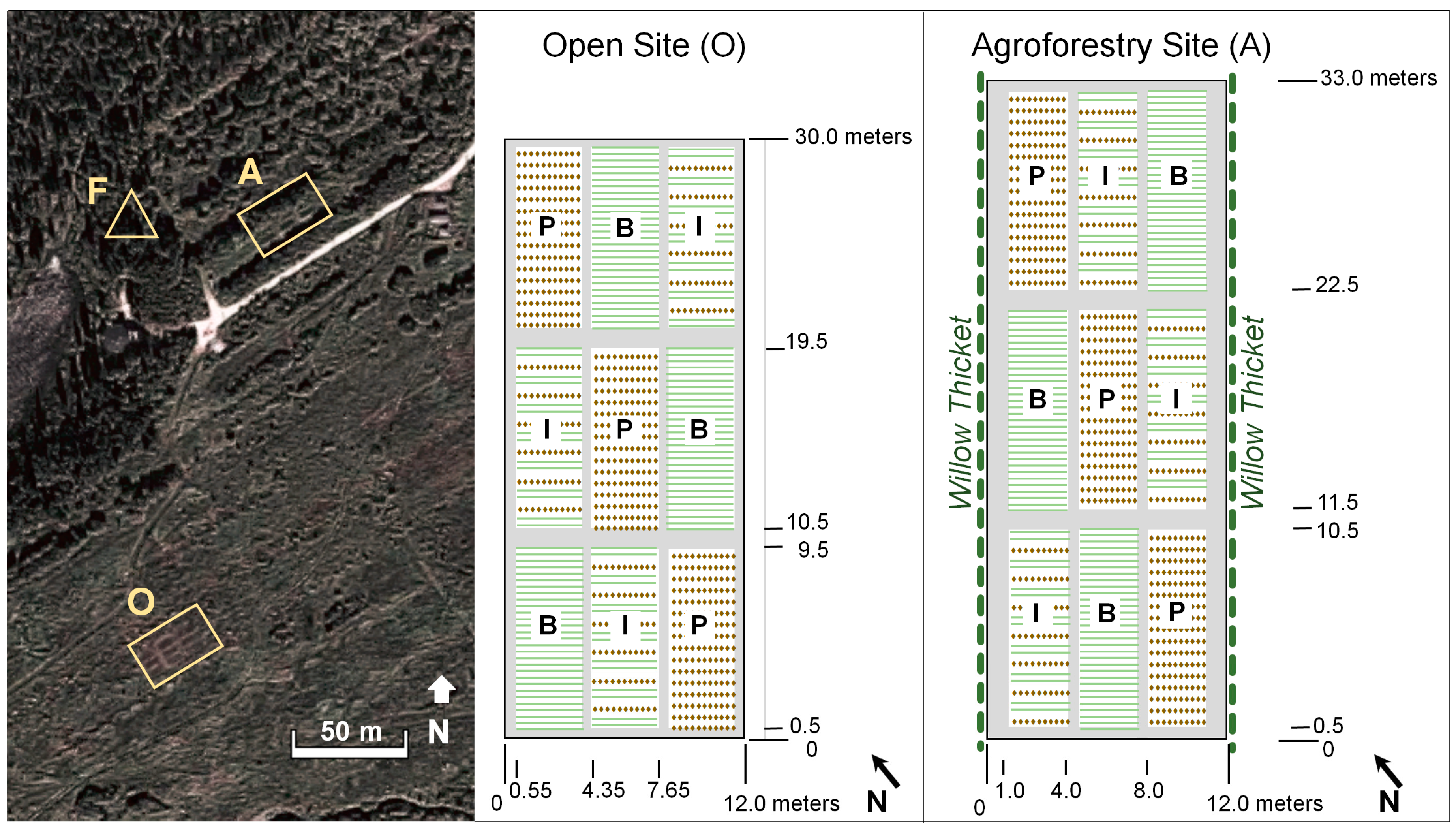
2.2. Site Preparations
2.3. Crop Tissue Analysis
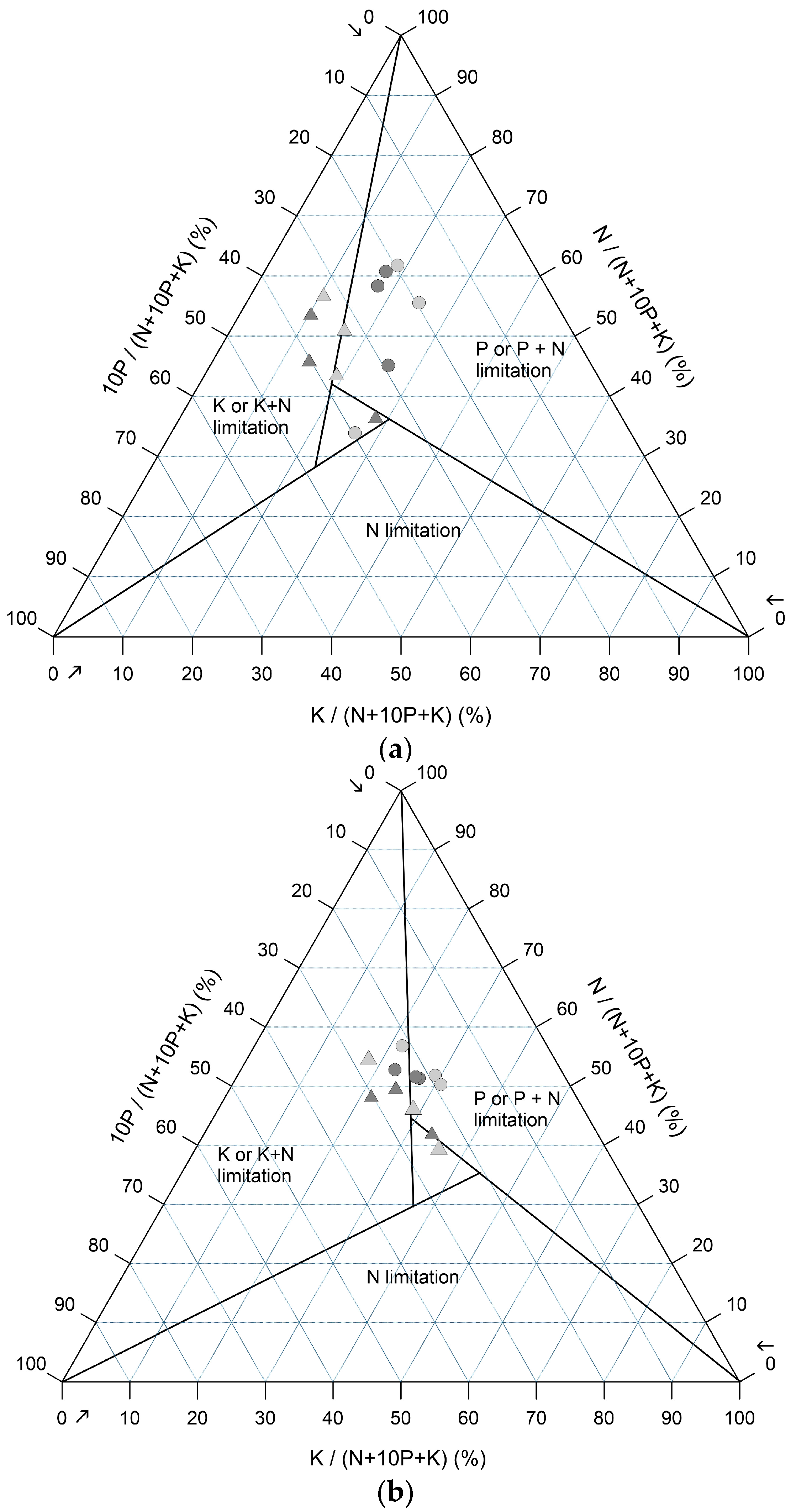
2.4. NPK Ternary Diagrams
2.5. Statistical Analysis
3. Results
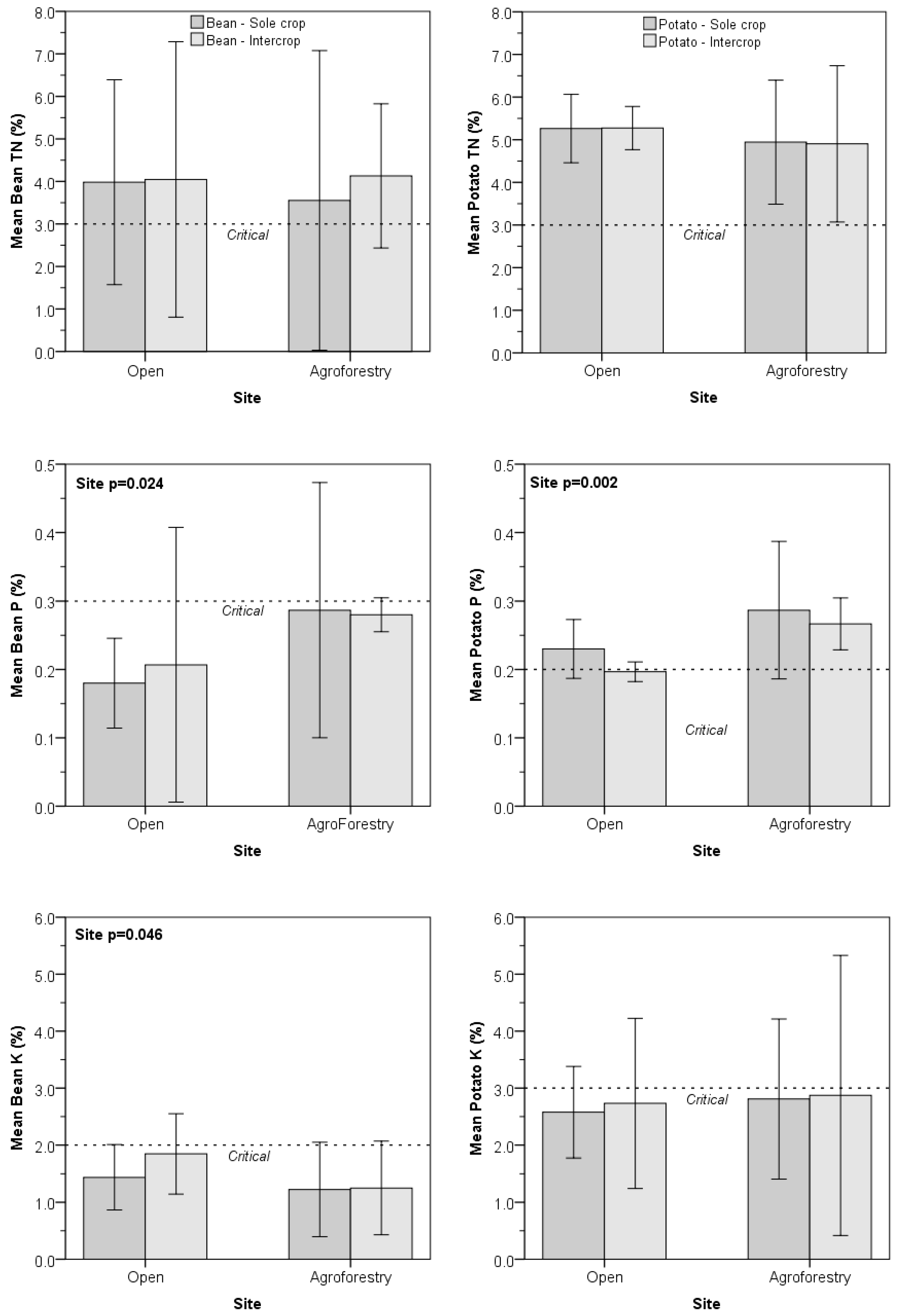
3.1. Comparison for Site and Management
3.2. Comparision to Critical Levels
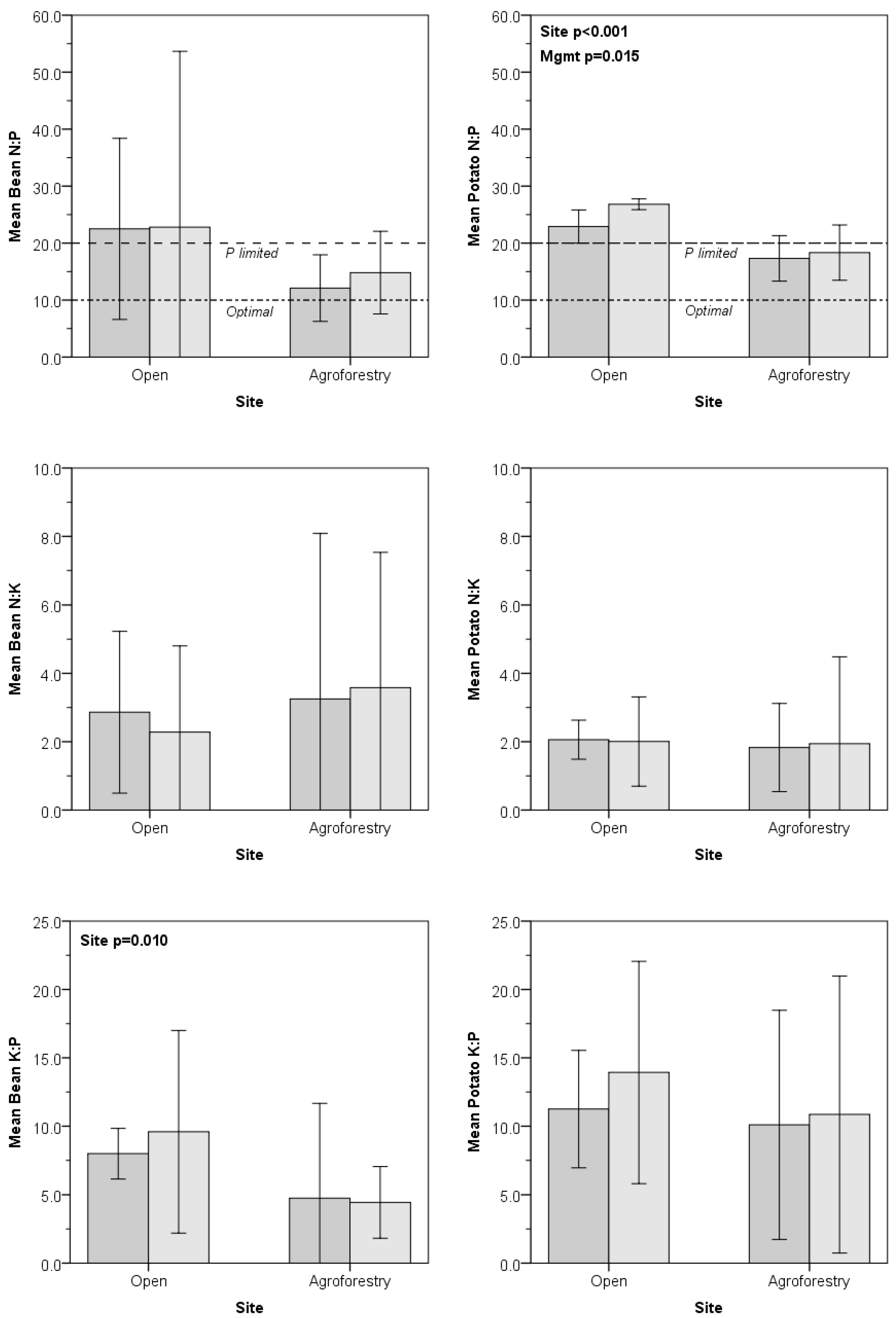
3.3. Determination of N, P, and K Limitations with Ternary Plots
4. Discussion
4.1. Nitrogen Mineralization Occuring in Subarctic Cultivated Soils
4.2. Agroforestry Sites Had More Available Phosphorus
4.3. Potassium Deficient Alkaline Soils Promote Nutrient Competition in Bean–Potato Intercropping Systems
4.4. Future Research on Local Phosphorus and Potassium Fertilizer in Northern Calcareous Soil
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Skinner, K.; Hanning, R.M.; Tsuji, L.J. Barriers and supports for healthy eating and physical activity for First Nation youths in Northern Canada. Int. J. Circumpolar Health 2006, 65, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.K.R. Climate change mitigation: A low-hanging fruit of agroforestry. In Agroforestry and the Future of Global Land Use; Nair, P.K.R., Garrity, D., Eds.; Springer Science and Business Media: Dordrecht, The Netherlands, 2012; pp. 31–67. ISBN 9789400746763. [Google Scholar]
- Tarasuk, V.; Mitchell, A.; Dachner, N. Household food insecurity in Canada, 2012. In Research to Identify Policy Options to Reduce Food Insecurity; PROOF: Toronto, ON, Canada, 2014. [Google Scholar]
- Schiff, R.; Brunger, F. Northern food networks: Building collaborative efforts for food security in remote Canadian aboriginal communities. J. Agric. Food Syst. Community Dev. 2013, 3, 31–45. [Google Scholar] [CrossRef]
- Skinner, K.; Burnett, K.; Williams, P.; Martin, D.; Stothart, C.; LeBlanc, J.; Veeraraghavan, G.; Sheedy, A. Challenges in assessing food environments in northern and remote communities in Canada. Can. J. Public Health 2016, 107, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Spiegelaar, N.F.; Tsuji, L.J.; Oelbermann, M. The Potential Use of Agroforestry Community Gardens as a Sustainable Import-Substitution Strategy for Enhancing Food Security in Subarctic Ontario, Canada. Sustainability 2013, 5, 4057–4075. [Google Scholar] [CrossRef]
- Leahey, A. Soil and agricultural problems in subarctic and arctic Canada. Arctic 1954, 7, 249–254. [Google Scholar] [CrossRef]
- Desmarais, A.A.; Wittman, H. Farmers, foodies and First Nations: Getting to food sovereignty in Canada. J. Peasant Stud. 2014, 41, 1153–1173. [Google Scholar] [CrossRef]
- Grünzweig, J.M.; Sparrow, S.D.; Chapin, F.S. Impact of forest conversion to agriculture on carbon and nitrogen mineralization in subarctic Alaska. Biogeochemistry 2003, 64, 271–296. [Google Scholar] [CrossRef]
- Alamenciak, T. How Climate Change Threatens Food Security in Northern Ontario. TVO. Available online: http://tvo.org/article/current-affairs/the-food-chain/how-climate-change-threatens-food-security-in-northern-ontario (accessed on 1 September 2017).
- Lukac, M.; Godbold, D.L. Soil Ecology in Northern Forests: A Belowground View of a Changing World; Cambridge University Press: Cambridge, UK, 2011; ISBN 9780521714211. [Google Scholar]
- Koenig, R.T.; Cochran, V.L. Decomposition and nitrogen mineralization from legume and non-legume crop residues in a subarctic agricultural soil. Biol. Fert. Soils 1994, 17, 269–275. [Google Scholar] [CrossRef]
- Stevenson, K.T.; Rader, H.B.; Alessa, L.; Kliskey, A.D.; Pantoja, A.; Clark, M.; Smeenk, J.; Giguère, N. Sustainable agriculture for Alaska and the circumpolar North: Part II. Environmental, geophysical, biological and socioeconomic challenges. Arctic 2014, 67, 296–319. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Ambus, P.; Jensen, E.S. Interspecific competition, N use and interference with weeds in pea–barley intercropping. Field Crop. Res. 2001, 70, 101–109. [Google Scholar] [CrossRef]
- Brandle, J.R.; Hodges, L.; Zhou, X.H. Windbreaks in North American agricultural systems. Agrofor. Syst. 2004, 61, 61–78. [Google Scholar]
- Brooker, R.W.; Bennett, A.E.; Cong, W.-C.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Lannetta, P.P.M.; Jones, H.G.; Karley, A.J.; et al. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altieri, M.A.; Nicholls, C.I.; Montalba, R. Technological Approaches to Sustainable Agriculture at a Crossroads: An Agroecological Perspective. Sustainability 2017, 9, 349. [Google Scholar] [CrossRef]
- Vandermeer, J.H. The Ecology of Intercropping; Cambridge University Press: New York, NY, USA, 1992; pp. 86–91. ISBN 0521346894. [Google Scholar]
- Nair, P.R. An Introduction to Agroforestry; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1993; pp. 272–333. ISBN 0792321340. [Google Scholar]
- Zhang, W.-P.; Liu, G.-C.; Sun, J.-H.; Fornara, D.; Zhang, L.-Z.; Zhang, F.-F.; Li, L. Temporal dynamics of nutrient uptake by neighbouring plant species: Evidence from intercropping. Funct. Ecol. 2017, 31, 469–479. [Google Scholar] [CrossRef]
- Gordon, A.; Newman, S. Temperate Agroforestry Systems; CAB International: Cambridge, MA, USA, 1997; pp. 263–269. ISBN 0851991475. [Google Scholar]
- Wang, X.; Deng, X.; Pu, T.; Song, C.; Yong, T.; Yang, F.; Sun, X.; Liu, W.; Yan, Y.; Du, J.; et al. Contribution of interspecific interactions and phosphorus application to increasing soil phosphorus availability in relay intercropping systems. Field Crop. Res. 2017, 204, 12–22. [Google Scholar] [CrossRef]
- Food and Agriculture Organizations of the United Nations (FAO). International Year of the Potato. Potato World, 2008. Available online: http://www.fao.org/potato-2008/en/world/index.html (accessed on 8 November 2014).
- Barbeau, C.D.; Oelbermann, M.; Karagatzides, J.D.; Tsuji, L.J. Sustainable Agriculture and Climate Change: Producing Potatoes (Solanum tuberosum L.) and Bush Beans (Phaseolus vulgaris L.) for Improved Food Security and Resilience in a Canadian Subarctic First Nations Community. Sustainability 2015, 7, 5664–5681. [Google Scholar] [CrossRef]
- Midmore, D.J. Scientific basis and scope for further improvement of intercropping with potato in the tropics. Field Crop. Res. 1990, 25, 3–24. [Google Scholar] [CrossRef]
- Sharaiha, R.K.; Battikhi, A. Potato-corn intercropping—Microclimate modification and yield advantages. Dirasat. Agric. Sci. 2002, 29, 97–108. [Google Scholar]
- Li, C.; He, X.; Zhu, S.; Zhou, H.; Wang, Y.; Li, Y.; Yang, J.; Fan, J.; Yang, J.; Wang, G.; et al. Crop diversity for yield increase. PLoS ONE 2009, 4, e8049. [Google Scholar] [CrossRef] [PubMed]
- Raei, Y.; Weisany, W.; Ghassemi-Golezani, K.; Torabian, S. Effects of additive intercropping on field performance of potato and green bean at different densities. BFIJ 2015, 7, 534–540. [Google Scholar]
- Fan, Z.; An, T.; Wu, K.; Zhou, F.; Zi, S.; Yang, Y.; Xue, G.; Wu, B. Effects of intercropping of maize and potato on sloping land on the water balance and surface runoff. Agric. Water Manag. 2016, 166, 9–16. [Google Scholar] [CrossRef]
- Dua, V.K.; Kumar, S.; Jatav, M.K. Effect of nitrogen application to intercrops on yield, competition, nutrient use efficiency and economics in potato (Solanum Tuberosum L.) + French bean (Phaseolus Vulgaris L.) system in north-western hills of India. Legume Res. 2017, 40, 698–703. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, G.; Zhang, E.; Zhang, B.; He, C.; Wang, Q.; Huang, G. Effect of phosphorus application and strip-intercropping on yield and some wheat-grain components in a wheat/maize/potato intercropping system. Afr. J. Agric. Res. 2011, 6, 5860–5869. [Google Scholar]
- Rezig, M.; Sahli, A.; Hachicha, M.; Jeddi, F.B.; Harbaoui, Y. Potato (Solanum tuberosum L.) and Bean (Phaseolus vulgaris L.) In Sole Intercropping: Effects on Light Interception and Radiation Use Efficiency. J. Agric. Sci. 2013, 5, 67–77. [Google Scholar]
- Barbeau, C.D.; Wilton, M.J.; Oelbermann, M.; Karagatzides, J.D.; Tsuji, T.J.S. Local food production in a Subarctic Indigenous Community: The use of willow (Salix spp.) windbreaks to increase the yield of intercropped potatoes (Solanum tuberosum L.) and Bush Bean (Phaseolus vulgaris L.) Intercrops. Int. J. Agric. Sustain. 2017, 1–11. [Google Scholar] [CrossRef]
- Hori, Y.; Tam, B.; Gough, W.A.; Ho-Foong, E.; Karagatzides, J.D.; Tsuji, L.T.S. Use of traditional environmental knowledge to assess the impact of climate change on subsistence fishing in the James Bay Region of Northern Ontario, Canada. Rural Remote Health 2012, 12, 1878. Available online: https://www.rrh.org.au/journal/article/1878 (accessed on 1 December 2017). [PubMed]
- Hanson, H.C. Muskeg as sharp-tailed grouse habitat. Wilson Bull. 1953, 65, 235–241. [Google Scholar]
- Minister of Supply and Services Canada (MSSC). A National Ecological Framework for Canada: Hudson Plains Ecozone and James Bay Lowland; Minister of Supply and Services Canada (MSSC): Ottawa, ON, Canada, 1996. [Google Scholar]
- Reuter, D.; Robinson, J.B. Plant Analysis: An Interpretation Manual; Inkata Press: Melbourne, Australia, 1986; pp. 151–175. ISBN 0909605416. [Google Scholar]
- Hack, H.; Gall, H.; Klemke, T.H.; Klose, R.; Meier, U.; Stauss, R.; Witzenberger, A. Phenological growth stages and BBCH-identification keys of potato (Solanum tuberosum L.). In Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph; Meier, U., Ed.; Blackwell Wissenschafts-Verlag: Berlin, Germany, 2001; pp. 44–52. Available online: http://www.reterurale.it/downloads/BBCH_engl_2001.pdf (accessed on 21 November 2017).
- Feller, C.; Blieholder, H.; Buhr, L.; Hack, M.; Hess, M.; Klose, R.; Meier, U.; Stauss, R.; van den Boom, T.; Weber, E. Phenological growth stages and BBCH-identification keys of Bean (Phaseolus vulgaris var. nanus L.). In Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph; Meier, U., Ed.; Blackwell Wissenschafts-Verlag: Berlin, Germany, 2001; pp. 141–144. Available online: http://www.reterurale.it/downloads/BBCH_engl_2001.pdf (accessed on 21 November 2017).
- Hochmuth, G.; Maynard, D.; Vavrina, C.; Hanlon, E.; Simonne, E. Plant Tissue Analysis and Interpretation for Vegetable Crops in Florida. IDAS Extension Special Series. 1991. Available online: http://www.soilwealth.com.au/imagesDB/wysiwyg/VegetableNutritionGuide-Florida.pdf (accessed on 1 September 2017).
- OMAFRA. Personal Communication. In Crops Specialists; Van Dyk, D., Roddy, E., Eds.; OMAFRA: Guelph, ON, Canada, 2016. [Google Scholar]
- Olde Venterink, H.; Wassen, M.J.; Verkroost, A.W.M.; De Ruiter, P.C. Species richness-productivity patterns differ between N-, P-, and K-limited Wetlands. Ecology 2003, 84, 2191–2199. [Google Scholar] [CrossRef]
- Güsewell, S. N: P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Lucas, R.E.; Davis, J. Relationships between pH values of organic soils and availabilities of 12 plant nutrients. Soil Sci. 1961, 92, 177–182. [Google Scholar] [CrossRef]
- Leidi, E.O.; Rodríguez-Navarro, D.N. Nitrogen and phosphorus availability limit N2 fixation in bean. New Phytol. 2000, 147, 337–346. [Google Scholar] [CrossRef]
- Rydin, H.; Jeglum, J.K.; Hooijer, A. The Biology of Peatlands; Oxford University Press: New York, NY, USA, 2006; pp. 155–169. ISBN 0198528728. [Google Scholar]
- Van Meeteren, M.J.M.; Tietema, A.; Westerveld, J.W. Regulation of microbial carbon, nitrogen, and phosphorus transformations by temperature and moisture during decomposition of Calluna vulgaris litter. Biol. Fert. Soils 2007, 44, 103–112. [Google Scholar] [CrossRef]
- Coombe, D.E. Notes on calcicolous communities and peat formation in Norwegian Lappland. J. Ecol. 1951, 39, 33–62. [Google Scholar] [CrossRef]
- Rytter, R.M.; Hansson, A.C. Seasonal amount, growth and depth distribution of fine roots in an irrigated and fertilized Salix viminalis L. plantation. Biomass Bioenergy 1996, 11, 129–137. [Google Scholar] [CrossRef]
- Radersma, S.; Grierson, P.F. Phosphorus mobilization in agroforestry: Organic anions, phosphatase activity and phosphorus fractions in the rhizosphere. Plant Soil 2004, 259, 209–219. [Google Scholar] [CrossRef]
- Hrynkiewicz, K.; Baum, C.; Leinweber, P. Mycorrhizal community structure, microbial biomass P and phosphatase activities under Salix polaris as influenced by nutrient availability. Eur. J. Soil Biol. 2009, 45, 168–175. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Thormann, M.N.; Currah, R.S.; Bayley, S.E. The mycorrhizal status of the dominant vegetation along a peatland gradient in southern boreal Alberta, Canada. Wetlands 1999, 19, 438–450. [Google Scholar] [CrossRef]
- Van der Heijden, E.W.; Kuyper, T.W. Ecological strategies of ectomycorrhizal fungi of Salix repens: Root manipulation versus root replacement. Oikos 2003, 103, 668–680. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Quigley, M.F. Willows beyond wetlands: Uses of Salix L. species for environmental projects. Water Air Soil Poll. 2005, 162, 183–204. [Google Scholar] [CrossRef]
- Thevathasan, N.V.; Gordon, A.M.; Bradley, R.; Cogliastro, A.; Folkard, P.; Grant, R.; Kort, J.; Liggins, L.; Njenga, F.; Olivier, A.; et al. Agroforestry research and development in Canada: The way forward. In Agroforestry and the Future of Global Land Use; Nair, P.K.R., Garrity, D., Eds.; Springer Science and Business Media: Dordrecht, The Netherlands, 2012; pp. 247–283. ISBN 9789400746763. [Google Scholar]
- Smith, J.; Pearce, B.D.; Wolfe, M.S. Reconciling productivity with protection of the environment: Is temperate agroforestry the answer? Renew. Agric. Food Syst. 2013, 28, 80–92. [Google Scholar] [CrossRef]
- Bird, P.R. Tree windbreaks and shelter benefits to pasture in temperate grazing systems. Agrofor. Syst. 1998, 41, 35–54. [Google Scholar] [CrossRef]
- Foereid, B.; Bro, R.; Mogensen, V.O.; Porter, J.R. Effects of windbreak strips of willow coppice—Modelling and field experiment on barley in Denmark. Agric. Ecosyst. Environ. 2002, 93, 25–32. [Google Scholar] [CrossRef]
- Jose, S.; Gillespie, A.R.; Pallardy, S.G. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 61, 237–255. [Google Scholar]
- Rao, M.R.; Nair, P.K.R.; Ong, C.K. Biophysical interactions in tropical agroforestry systems. Agrofor. Syst. 1997, 38, 3–50. [Google Scholar] [CrossRef]
- Shukla, A.; Kumar, A.; Jha, A.; Chaturvedi, O.P.; Prasad, R.; Gupta, A. Effects of shade on arbuscular mycorrhizal colonization and growth of crops and tree seedlings in Central India. Agrofor. Syst. 2009, 76, 95–109. [Google Scholar] [CrossRef]
- Sundström, E.; Magnusson, T.; Hånell, B. Nutrient conditions in drained Peatlands along a north-south gradient in Sweden. For. Ecol. Manag. 2000, 126, 149–161. [Google Scholar] [CrossRef]
- Harrap, F.E.G. Some aspects of the potash nutrition of the potato. J. Sci. Food Agric. 1960, 11, 293–298. [Google Scholar] [CrossRef]
- Overstreet, R.; Jacobson, L.; Handley, R. The effect of calcium on the absorption of potassium by barley roots. Plant Physiol. 1952, 27, 583. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, L.S.; Wilton, M.J.; Spiegelaar, N.F.; Oelbermann, M.; Barbeau, C.D.; Tsuji, C.J.D.; Liberda, E.N.; Meldrum, R.; Karagatzides, J.D. Enhancing Food Security in Subarctic Canada in the Context of Climate Change: The Harmonization of Indigenous Harvesting Pursuits and Agroforestry Activities to Form a Sustainable Import-substitution Strategy. In Sustainable Solutions for Food Security: Combating Climate Change by Adaptation; Sarkar, A., vanLoon, G.W., Sensarma, S.J., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; in press. [Google Scholar]
- Erich, M.S. Agronomic effectiveness of wood ash as a source of phosphorus and potassium. J. Environ. Qual. 1991, 20, 576–581. [Google Scholar] [CrossRef]
- Kuba, T.; Tschöll, A.; Partl, C.; Meyer, K.; Insam, H. Wood ash admixture to organic wastes improves compost and its performance. Agric. Ecosyst. Environ. 2008, 127, 43–49. [Google Scholar] [CrossRef]
- Guppy, C.N.; Menzies, N.W.; Moody, P.W.; Blamey, F.P.C. Competitive sorption reactions between phosphorus and organic matter in soil: A review. Soil Res. 2005, 43, 189–202. [Google Scholar] [CrossRef]
- Leytem, A.B.; Mikkelsen, R.L. The nature of phosphorus in calcareous soils. Better Crops 2005, 89, 11–13. [Google Scholar]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Creamer, W.; Christ, R.; Chruch, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: synthesis report. In Contribution of Working Groups I, II, III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; pp. 1–151. [Google Scholar]
- MacDonald, G.M.; Gajewski, K. The northern treeline of Canada. In Geographical Snapshots of North America: Commemorating the 27th Congress of the International Geographical Union and Assembly; Janelle, D.G., Ed.; Guilford Press: New York, NY, USA, 1992; pp. 34–37. ISBN 089862889X. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Agroforestry | Open | Boreal Forest |
|---|---|---|---|
| Management | Cultivated for 3 Years Enclosed with Willow Trees | Cultivated for 3 Years in an Open Field | Uncultivated Forest Soil |
| Soil Characteristics | n = 9 | n = 9 | n = 3 |
| pH * | 7.7 ± 0.08 | 7.6 ± 0.03 | 7.5 ± 0.11 |
| Bulk density (g cm−3) | 0.66 ± 0.07 | 0.66 ± 0.06 | 0.30 ± 0.12 |
| C/N * | 14.9 ± 0.48 | 15.7 ± 0.93 | 22.3 ± 3.29 |
| CEC (cmol+ kg−1) *† | 40.6 ± 4.21 | 41.0 ± 1.74 | 30.8 ± 12.77 |
| Soil Temperature (°C) | n/a | ||
| Mean ± SD § | 17.5 ± 2.80 | 15.4 ± 2.77 | |
| Minimum | 12.4 | 7.4 | |
| Maximum | 26.7 | 21.7 | |
| Soil Moisture (WFPS%) ‡ | n/a | ||
| Mean ± SD | 38.6 ± 0.017 | 51.9 ± 0.011 | |
| Minimum | 35.9 | 50.6 | |
| Maximum | 45.5 | 55.9 |
| Nutrient | Low | Critical | Adequate | Normal Max | High |
|---|---|---|---|---|---|
| Bush Bean (Phaseolus vulgaris L.) MRM * trifoliate, In bud to early flower | |||||
| N | 2.0 | 3.0 | 3.5 | 4.0 | 5.5 |
| P | 0.25 | 0.30 | 0.4 | 0.5 | 0.6 |
| K | 1.5 | 2.0 | 2.5 | 3.0 | 4.0 |
| Mg | - | 0.26 | - | - | - |
| Ca | - | 0.8 | - | - | - |
| Potato (Solanum tuberosum L.)—MRM leaf + petiole—In bud to early flower | |||||
| N | 2.0 | 3.0 | 4.0 | 5.0 | 6.0 |
| P | 0.15 | 0.20 | 0.40 | 0.50 | 0.60 |
| K | 2.0 | 3.0 | 4.0 | 5.0 | 6.0 |
| Mg | - | 0.25 | 0.5 | - | - |
| Ca | - | 0.6 | 1.0 | - | - |
| Nutrient | Bush Bean (Phaseolus vulgaris L.) | Potato (Solanum tuberosum L.) | ||||
|---|---|---|---|---|---|---|
| Site | Mgmt | Site × Mgmt | Site | Mgmt | Site × Mgmt | |
| N | 0.797 | 0.637 | 0.705 | 0.273 | 0.960 | 0.934 |
| P | 0.024 * | 0.694 | 0.605 | 0.002 | 0.083 | 0.792 |
| K | 0.046 | 0.240 | 0.294 | 0.639 | 0.781 | 0.906 |
| N:P | 0.054 | 0.782 | 0.739 | <0.001 | 0.016 | 0.184 |
| N:K | 0.339 | 0.887 | 0.595 | 0.700 | 0.938 | 0.828 |
| K:P | 0.010 | 0.689 | 0.502 | 0.315 | 0.394 | 0.676 |
| Mg | 0.264 | 0.963 | 0.628 | 0.504 | 0.558 | 0.824 |
| Ca | 0.736 | 0.978 | 0.457 | 0.070 | 0.391 | 0.606 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilton, M.J.; Karagatzides, J.D.; Tsuji, L.J.S. Nutrient Concentrations of Bush Bean (Phaseolus vulgaris L.) and Potato (Solanum tuberosum L.) Cultivated in Subarctic Soils Managed with Intercropping and Willow (Salix spp.) Agroforestry. Sustainability 2017, 9, 2294. https://0-doi-org.brum.beds.ac.uk/10.3390/su9122294
Wilton MJ, Karagatzides JD, Tsuji LJS. Nutrient Concentrations of Bush Bean (Phaseolus vulgaris L.) and Potato (Solanum tuberosum L.) Cultivated in Subarctic Soils Managed with Intercropping and Willow (Salix spp.) Agroforestry. Sustainability. 2017; 9(12):2294. https://0-doi-org.brum.beds.ac.uk/10.3390/su9122294
Chicago/Turabian StyleWilton, Meaghan J., Jim D. Karagatzides, and Leonard J. S. Tsuji. 2017. "Nutrient Concentrations of Bush Bean (Phaseolus vulgaris L.) and Potato (Solanum tuberosum L.) Cultivated in Subarctic Soils Managed with Intercropping and Willow (Salix spp.) Agroforestry" Sustainability 9, no. 12: 2294. https://0-doi-org.brum.beds.ac.uk/10.3390/su9122294