Dynamics of Laterality in Lake Tanganyika Scale-Eaters Driven by Cross-Predation

1
Animal Ecology, Department of Zoology, Graduate School of Science, Kyoto University, Kitashirakawa-Oiwakecho, Sakyo 606-8502, Japan
2
Animal Sociology, Department of Biology and Geosciences, Graduate School of Science, Osaka City University, 3-3-138 Sugimoto, Sumiyoshi, Osaka 558-8585, Japan
3
Graduate School of Humanities and Sciences, Nara Women’s University, Nara 630-8506, Japan
*
Author to whom correspondence should be addressed.
Symmetry 2019, 11(1), 119; https://0-doi-org.brum.beds.ac.uk/10.3390/sym11010119
Submission received: 24 November 2018
/
Revised: 13 January 2019
/
Accepted: 14 January 2019
/
Published: 20 January 2019
(This article belongs to the Special Issue Left Versus Right Asymmetries of Brain and Behaviour)
Abstract
:Scale-eating cichlid fishes, Perissodus spp., in Lake Tanganyika have laterally asymmetrical bodies, and each population is composed of righty and lefty morphs. Righty morphs attack the right side of prey and lefty morphs do the opposite. This anti-symmetric dimorphism has a genetic basis. Temporal changes in the frequencies of morphs in two cohabiting scale-eating species (Perissodus microlepis and P. straeleni) were investigated over a 31-year period on a rocky shore at the southern end of the lake. Dimorphism was maintained dynamically during the period in both species, and the frequencies oscillated with a period of about four years in a semi-synchronized manner. Recent studies have indicated that this type of anti-symmetric dimorphism is shared widely among fishes, and is maintained by frequency-dependent selection between predator and prey species. The combinations of laterality in each scale-eater and its victim were surveyed. The results showed that “cross-predation”, in which righty predators catch lefty prey and lefty predators catch righty prey, occurred more frequently than the reverse combination (“parallel-predation”). The cause of the predominance of cross-predation is discussed from the viewpoint of the physical and sensory abilities of fishes.
1. Introduction
Lateral bias of behavior has been observed in various animals, particularly vertebrates. Among vertebrates, fish are expected to exhibit primordial forms of vertebrate laterality, as they are the most ancestral group. Numerous studies have used fish to elucidate the causes of behavioral laterality, and many have demonstrated an association between behavioral laterality and brain lateralization [1,2,3,4,5,6,7,8]. Many studies on morphological asymmetry have been conducted in both invertebrates and vertebrates including fish, from which three types of asymmetry have been categorized based on the frequency distribution of measured laterality: fluctuating asymmetry (unimodal and symmetrical distribution), directional asymmetry (unimodal distribution shifted from symmetry) and anti-symmetry (bimodal distribution) [9]. The relationship between behavioral laterality and morphological asymmetry, especially for anti-symmetry, had rarely been investigated aside from asymmetry of the brain [6,10,11].
Some theoretical and/or empirical studies have investigated the mechanism that is responsible for maintaining the lateralization in one population from the viewpoint of cerebral lateralization [12,13,14], but few studies have been done to analyze the mechanism that maintains the anti-symmetric dimorphism in one population. In particular, the population in which the dimorphism is maintained dynamically, i.e., the frequency of righty and lefty morphs changes temporally with in a fixed range, has not been investigated except for scale-eating cichlid fish in Lake Tanganyika [15]. Note that, if the frequency of righty and lefty morphs is changeable, it is not the laterality at the population level, even though the laterality is obvious at the individual level.
The scale-eating cichlid fishes, Perissodus spp., in Lake Tanganyika have laterally asymmetric bodies, and each population is composed of righty and lefty individuals or morphs [15,16,17]. Righty individuals attack the right side of prey and lefty fish do the opposite [15,18,19]. Similar morphological and behavioral laterality was also observed in an Amazonian characid scale-eater, Exodon sp. [20]. This type of dimorphism has a genetic basis [15,21,22,23,24]. The ratio of laterality (frequency of righty morphs in each population) of P. microlepis oscillates around 0.5 with a period of five years, and this balance appears to be maintained by frequency-dependent selection (minority advantage) mediated by the vigilance of prey [15,25,26].
A field study of the balance of polymorphism in P. microlepis was conducted on rocky shores at the northern end of the lake (near Uvira, Democratic Republic of the Congo), where P. microlepis was essentially the only species of scale-eater in the fish community [15]. From this study, further questions arose, such as how laterality interacts in a locality where two species of scale-eater coexist, and whether polymorphism is maintained in the two species.
In order to know the temporal change of the ratio of laterality where two species of scale-eater cohabit, we monitored the ratios of their laterality on a rocky shore at the southern end of the lake (near Mpulungu, Republic of Zambia) for 31 years from 1988 to 2018, where P. microlepis and P. straeleni cohabited [27,28]. The two species (Figure 1) are the sister species in phylogeny [29], and occupy the same feeding niche, i.e., they attack nearly the same species of cichlid fishes (mainly algal-feeders with high body depth such as Petrochromis spp., Cyathopharynx furcifer and Tropheus moorii) as prey of their scale-eating [17,30].
Another purpose of the present study was to investigate the relation between the laterality of scale-eater and that of their prey fish. Recently Hori et al. [31] demonstrated that almost all fishes have the same kind of laterality at various intensities in a similar way to that of scale-eaters, which had been suggested for various types of fish [21,32,33,34,35,36]. Given the findings that all fishes have laterality, we have to reconsider the mechanism responsible for maintaining that laterality and for driving the oscillation of the ratio of laterality. The most probable mechanism may still be the prey—predator interactions, but it has come to light that the mechanism is embedded in the genetic ability of both predator and prey as well as the prey’s switching of vigilance toward the majority morph of predator. Therefore, we have to examine the combination of laterality between each predator and its prey.
In the situation that all the fishes have laterality, two types of predation incident can be distinguished: A predator catches a prey of the same morph of laterality (righty predator catches righty prey and lefty one lefty prey) or a predator catches a prey of the opposite morph (righty predator catches lefty prey and lefty one righty prey). The former type can be called “parallel-predation” and the latter “cross-predation” [32,36,37,38]. Then, provided that the ratios of laterality of both predator and prey are maintained in any pattern, we can predict that the predation incidents in a community as a whole at any one time are biased toward an excess of one type of predation over the other, i.e., either that cross-predations are prominent over parallel-predation (predominance of cross-predation) or the reverse (predominance of parallel-predation), but not that both types of predation occur at a similar frequency (random-predation).
To verify this prediction, during the latter half of the field work (from 2006 to 2018) we observed the scale-eating in water and tried to collect both the scale-eater and its victim fish just after the scale-eating took placed, and examined the frequencies of predation types among the sampled pairs.
In short, the purpose of this study is to demonstrate the long-term dynamics of laterality in the two co-habiting species of scale-eater in one locality, and also to detect the predominance of either cross predation or parallel-predation in the events of scale-eating. Some discussion is made on the meaning of the pattern found.
2. Materials and Methods
2.1. Sampling of Scale-Eaters to Assess the Ratios of Laterality
We conducted a long-term survey on the ratios of laterality in the populations of the two species of scale-eater on a rocky shore at Kasenga Point in the southern end of Lake Tanganyika (near Mpulungu, Northern District, Republic of Zambia). It has been shown that the fish community in the littoral rocky shore of this lake is mainly composed of cichlid fishes and is very stable with the densities of most species being unchanged over many years [17,28,39]. Then, in order to know the effect of laterality of one species on another, it may be enough to know the ratios of laterality of the two species of scale-eater. In the same season (from September to November) in every year, fish samples of the two species were purchased from fish hauls by village fishermen at Kasenga Point. The hauls had been caught as their livelihood mainly by gill-net and sometimes by sein-net and angling every day. In every season, several individuals of each species (ranging from 10 to 200, the average was 132 for P. microlepis and 57 for P. straeleni) were collected in order to gain reliable ratios of laterality for each population at each time. This survey was conducted almost every year from 1988 to 2018, though we could not make the survey in two years in the beginning of this survey (1989 and 1991).
The laterality of each fish can be defined from the direction of the mouth opening [17,15,21]. Morphologically, the asymmetric mouth opening is due to either side of the joint, say, the right joint, between mandible and suspensorium taking a position frontward, ventrally, and outside compared to the opposite side of the joint [16]. The bending rightwards should mean that the right side of its head and flank have developed more compared to the left side [21]. This relation is seemingly held in other fishes [35,37]. The functional morphology and the quantitative measurement of the asymmetric mouth opening have been developed with separate works [31,36,40]. Note that the definition of laterality used in the present and recent studies differs from that used in earlier papers [15,16,17,33]. In the earlier papers, individuals with the mouth opening to the right were defined as “right-handed” or “dextral”. In the present and recent studies, usage of “lefty” reflects the fact that the left mandible of such ‘right-handed’ fish is larger than the right mandible [21,31], and the left eye is dominant [34,41].
Temporal change in the ratios of laterality (frequency of righty morphs in each population) in each species was determined using the samples mentioned above for the whole period, and the periodicity in the ratio of laterality, if any, was analyzed using the Fourier transform with the same method formalized in the Supplementary Material in the work of Yasughi and Hori [36].
2.2. Sampling of Pairs of Hunting Scale-Eaters and Their Victim Fish to Assess the Combination of Laterality at Each Predation Event
This sampling was carried out at Kasenga Point mainly in November of 2007, and supplementary samplings were added at the site in the same season from 2008 to 2018. In every survey, we used a scuba to trace each fish of either species of scale-eater in water 3–10 m deep. When observing the incidence of scale-eating, we immediately tried to catch both the scale-eater and its victim by spreading a short gill-net around them. Then, in the event that both the scale-eater and its victim were caught, the fishes were unloaded, fixed in water with ice, and kept in 10% formol. If only one fish was caught, it was released, and another tracing was started. Success in catching both fishes was achieved about once every 3 or 4 trials. In the laboratory, the laterality of the fishes was determined based on the direction of the mouth opening, and the combinations of their laterality were examined. These treatments were performed according to the Regulations on Animal Experimentation at Kyoto University. To know whether cross-predation or parallel-predation was predominant in each species of scale-eater, a statistical test was performed using odds ratios with a 95% confidential limit.
3. Results
3.1. Temporal Changes in the Laterality of Two Scale-Eaters
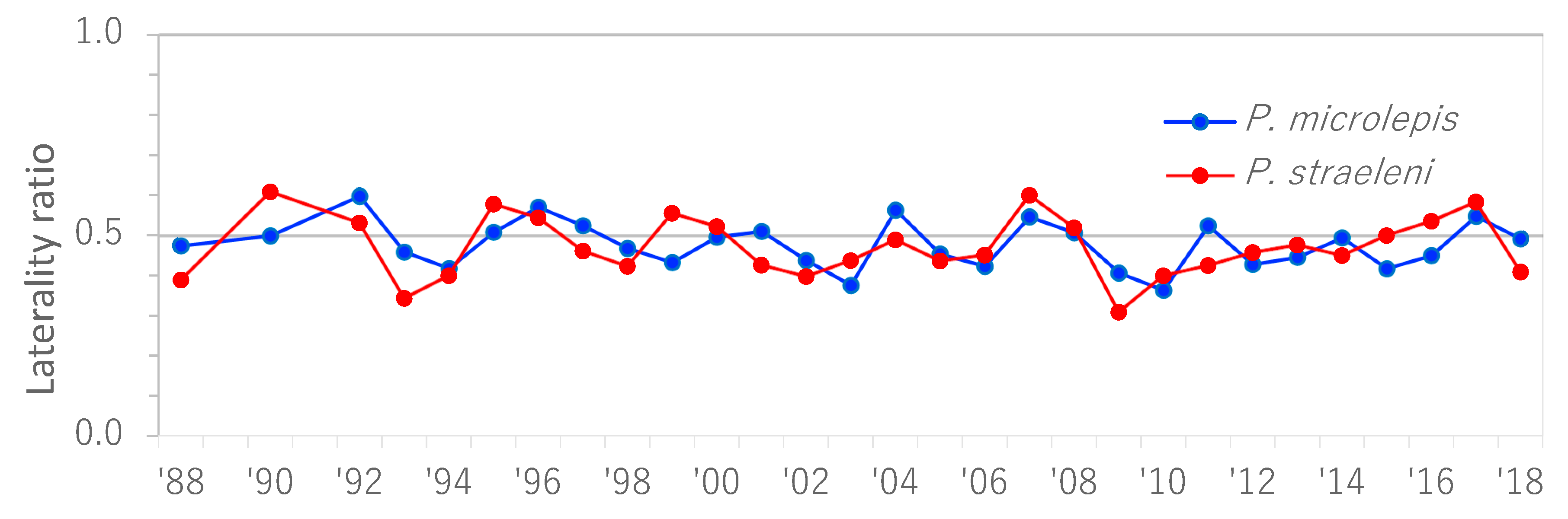
In Kasenga point, both lateral morphs of the two species of scale-eater were maintained for 31 years. The ratios of laterality of both species periodically and dynamically changed around a value of 0.5 but were almost always maintained within a range of 0.4 to 0.6 (Figure 2). The Fourier transform detected a significant period of cycle in the ratio of each species, i.e., 3.9 years (p-value < 0.001) for P. microlepis and 4.1 years (p-value < 0.01) for P. straeleni, indicating that the ratios of the two species oscillated with almost the same period.
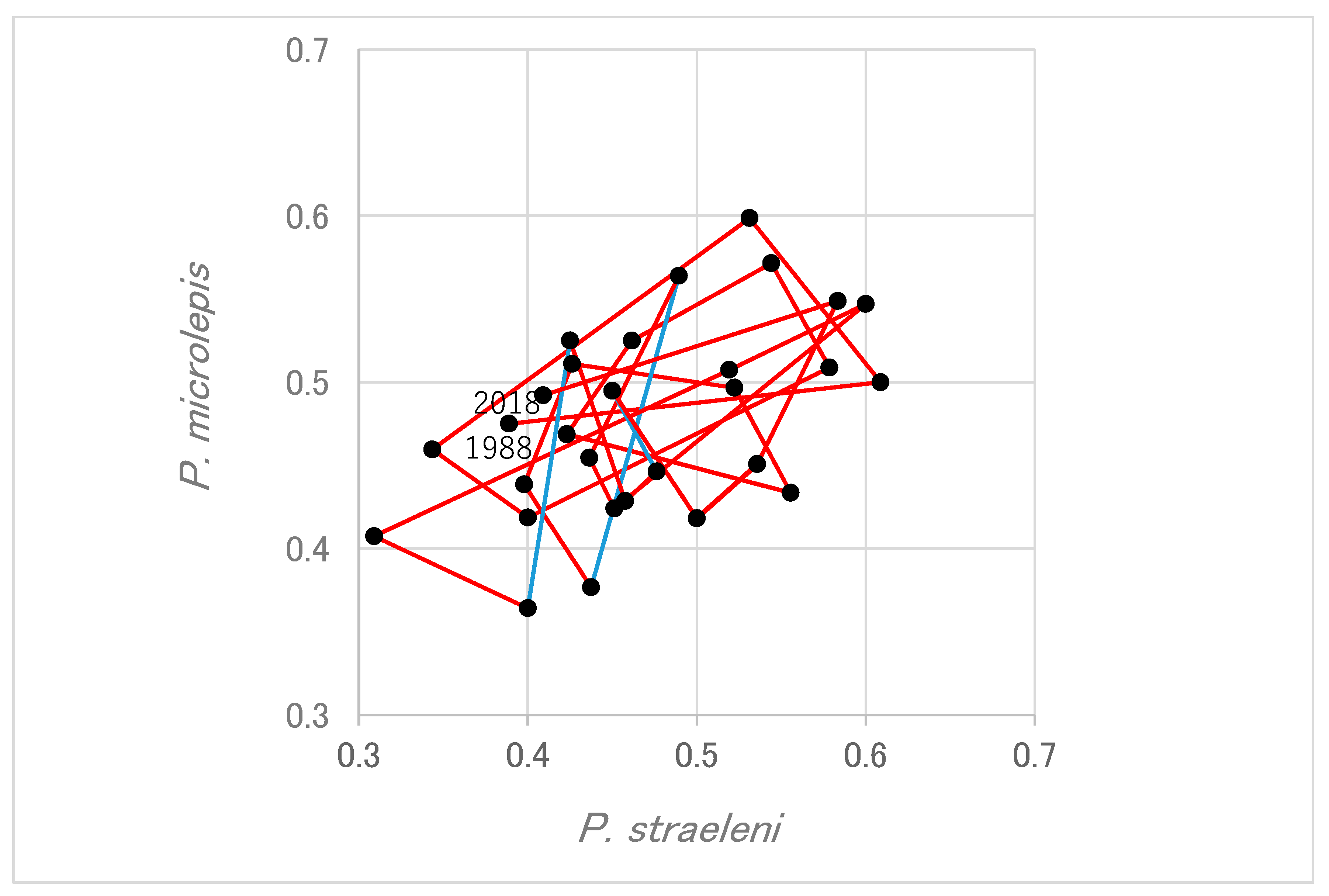
Relation of oscillation of laterality between the two species of scale-eater was examined by directly plotting the ratio of P. microlepis against that of P. straeleni (Figure 3). The plots scattered in a counterclockwise rotation around the equilibrium point in both species (coordinates (0.5, 0.5)). It suggests that the ratio of P. microlepis followed the periodic change of P. straeleni nearly one year later.
3.2. Correspondence of Fish Laterality between Individual Scale-Eater and Its Prey
We were able to collect 53 pairs of individual scale-eaters and prey fish for P. microlepis, and 51 pairs for P. straeleni. The “victim fish” (fish that have their scales eaten) were composed of Interochromis loocki, Petrochromis spp., Tropheus moorii, and Lamprologus callipterus, and they looked to be of little difference to the two species of scale-eater. The combination between the laterality of scale-eater and its prey exhibited a significant bias toward cross-predation in both species (Table 1). The odds ratio was 26.4 for P. microlepis (95% confidential limit; 24.9–27.9) and 28.88 for P. straeleni (27.4–30.4), indicating that cross-predation significantly occurred more frequently than parallel-predation (Mantel–Haenszel test; p < 0.01 for both species).
4. Discussion
4.1. Relation between the Two Species of Scale-Eater
Results showed that the lateral dimorphism of two species of scale-eater cohabiting in a rocky shore was maintained just as it was where only one species (P. microlepis) inhabited [15]. The ratios of the two species oscillated during the 30 years with nearly the same period (four years) and the same amplitude (0.2). These patterns indicate that the ratios of laterality of the two species interacted with each other. The change in ratio of P. microlepis appeared to follow that of P. straeleni.
This semi-synchronized pattern may be interpreted by the frequency-dependent selection (minority advantage) in scale-eating and also by the sharing of advantage between the minor morphs in frequency of the two species. A long-term census carried out in the same site (Takeuchi et al., 2010) [28] indicated that the density of the two species of scale-eater has remained rather constant with that of P. microlepis being 8.4 times higher than that of P. straeleni (the mean density of P. microlepis was 11.73/10 m2 (S.D., 28.76) and that of P. straeleni 1.39 (3.98)). If the minority advantage operates in this system, the minor morph at a time of P. straeleni, say, the righty morph, will get the higher fitness, and then the advantage should be shared by the righty morph of P. microlepis. This sharing of minority advantage may be the most responsible factor for the ratio of the laterality of majority scale-eaters in number, P. microlepis, following the oscillation of the minority one, P. straeleni. Using a theoretical model, Takahashi and Hori [26,42] examined the factor that generates an oscillation of polymorphism in two competing predators and found that the strong frequency-dependent selection was crucial for the polymorphism being dynamically maintained. In the next section, we examine the strength of the frequency-dependent selection between scale-eaters and their prey.
4.2. Predator and Prey Relationship
The results showed that righty scale-eaters predominantly succeeded in attacking lefty prey over righty prey, and vice versa for lefty scale-eaters in both P. microlepis and P. straeleni. A statistical test based on the common odds ratio was highly significant (Mantel–Haenszel test; p < 0.01), suggesting that “cross-predation” was 26 to 29 times more frequent than “parallel-predation”. Cross-predation should be crucially responsible for the oscillations being generated. Using a theoretical model, Takahashi and Hori [25] examined the factor that could generate an oscillation of polymorphism in a scale-eater and found that the strong frequency-dependent selection was essential for the oscillation around the equilibrium. The high level of predominance of cross-predation found in both scale-eaters seemed to account enough for the oscillation of laterality observed in their populations. In such a situation, the majority morph of scale-eater at any one time, say, righty scale-eaters, would exploit the lefty prey fish. Then the righty prey would be at an advantage in fitness and increase its own frequency after a period of time had elapsed, which would then favor the lefty morph of scale-eater. Furthermore, Nakajima et al. [43], using mathematical analyses and computer simulations, ascertained that under the predominance of cross-predation and high predation efficiency, the dimorphism was dynamically maintained in a one-predator–two-prey system as well as a three-trophic-levels system.
Yasugi and Hori [36] investigated the relationship between largemouth bass, Micropterus salmoides, and freshwater gobies, Rhinogobisu spp., in regards to their laterality, and found a significant bias toward cross-predation. Then, Yasugi and Hori [37] studied experimentally the kinematic causation of predominance of cross-predation. They found that every morph of laterality has a dominant side (either right or left) of the body in sensory and locomotion ability which may function deferentially in detecting and attacking a prey and making an attack in predator fish, and detecting and evading enemy and escaping in prey fish. Then, in the system where a predator stalks and attacks prey fish from behind, predation is more successful when lefty (righty) predator meets a righty (lefty) prey, and less successful when a lefty (righty) predator meets a lefty (righty) prey. Though no quantitative data were taken, we have the impression that scale-eaters can detect the laterality of prey that they targeted at an early stage of their pursuit, because the scale-eaters were often observed to pursue for a long time in cases of cross-predation. We think that they can detect the prey’s laterality judging by any delicate gesture in the prey’s movement, even though the laterality of each individual seems to be concealed from appearance [21].
Yasugi and Hori [38] also demonstrated that the dominance of cross-predation is characteristic of predators who attack prey from behind. A similar result was found in the investigation of stomach content analysis of the relation between piscivorous fishes belonging to Lamprologus (sensu lato) and their prey in regards to their laterality (Hori, unpublish data). Inversely, parallel-predation is dominant in predators who attack the benthic prey fish from ahead such as anglerfish, Lophinomus setigerus [38].
In this study we could not analyze the relation between ratios of predator and prey. The littoral fish community in Lake Tanganyika is so speciose and the interspecific interactions are highly complicated [17,27,30]. The interaction between piscivorous fishes and their prey and that between benthos-eating fishes and their prey may also be involved. Therefore, to analyze the temporal relation of laterality between predator and prey in the natural system, a more simple system such as one-predator–one-prey may be suitable. The fish community in a pelagic area in a temperate zone might provide such a system.
Furthermore, cross-predation may have a large effect on the structure of a fish community itself. Using a mathematical model and a computer simulation applied to a fish community of three trophic levels, Nakajima et al. [44] predicted that, when only one type of lateral morph exists in a species, the other type can invade, which suggests that dimorphism is maintained in all directly and indirectly interacting species of a community. Takahashi and Hori [42] theoretically showed that oscillation in laterality of morphs affects the coexistence of competing species. For an investigation of the relation between laterality and community structure in a natural system, however, the fish community in Lake Tanganyika may be suitable in spite of the complexity of its structure. The reason is that, as every population was very stable [17,28], the effect of frequency of each morph of a species on other fishes could be assessed only with periodic fish samplings from the community. This type of research is now in progress at the same site of the present study.
At present, it is difficult to evaluate the effect of frequency-dependent vigilance of prey fish on oscillation. Hori [15] demonstrated that prey fish actually showed such differential vigilance responding to the ambient ratio of laterality in scale-eaters. However, the result was reached in circumstances where the laterality of prey fish had not yet been recognized, and thus the two kinds of effects on oscillation, i.e., cross-predation and differential vigilance, were mingled. As the theoretical model [42] predicts that natural selection should be strong to generate the oscillation of the ratios of laterality, frequency-dependent vigilance of prey fish may have a large effect on the oscillation, as well as the predominance of cross-predation. The evaluation of the relative importance of the vigilance of prey fish remains for future investigation.
Author Contributions
Conceptualization, M.H. and S.T.; methodology, M.H. and S.T.; statistical analysis, S.T.; investigation, M.H., M.K. and S.A.; data collecting, M.H., M.K. and S.A.; data curation, M.H.; writing—original draft preparation, M.H.; writing—review and editing, M.K., S.A., S.T. and M.H.; project administration, M.H.; resources, M.H. and M.K.
Funding
This study was supported by grants from the Ministry of Education, Culture, Science and Technology, Japan (21st Century COE Program (A14), Global Coe Program (A06), Priority Area (14087203), and Scientific Research (B, 21370010, 15H05230, 16H05773, 17K14934) for MH and (B, 16H05773) for MK).
Acknowledgments
We are grateful to members of the Maneno Team (Tanganyika Research Project Team) and staff of LTRU, Mpulungu, Zambia, for their support. The research presented here was conducted with permission for fish research in Lake Tanganyika from the Zambian Ministry of Agriculture, Food, and Fisheries and complied with the current law in Zambia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bisazza, A.; Rogers, L.J.; Vallortigara, G. The origins of cerebral asymmetry: A review of evidence of behavioral and brain lateralization in fishes, reptiles and amphibians. Neurosci. Biobehav. Rev. 1998, 22, 411–426. [Google Scholar] [CrossRef]
- Vallortigara, G.; Rogers, L.J.; Bisazza, A. Possible evolutionary origins of cognitive brain lateralization. Brain Res. Rev. 1999, 30, 164–175. [Google Scholar] [CrossRef]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Vallortigara, G.; Versace, E. Laterality at the neural, cognitive, and behavioral levels. In APA Handbooks in Psychology. APA Handbook of Comparative Psychology: Basic Concepts, Methods, Neural Substrate, and Behavior; Call, J., Burghardt, G.M., Pepperberg, I.M., Snowdon, C.T., Zentall, T., Eds.; American Psychological Association: Washington, DC, USA, 2017; pp. 557–577. [Google Scholar]
- MacNeilage, P.; Rogers, L.J.; Vallortigara, G. Evolutionary origins of your left and right brain. Sci. Am. 2009, 301, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Rogers, L.J. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain Sci. 2005, 28, 575–633. [Google Scholar] [CrossRef]
- Vallortigara, G.; Chiandetti, C.; Sovrano, V.A. Brain asymmetry (animal). WIREs Cogn. Sci. 2011, 2, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J.; Vallortigara, G. When and why did brains break symmetry? Symmetry 2015, 7, 2181–2194. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry as a measure of development stability: Implications of non-normal distributions and power of statistical tests. Acta Zool. Fennica 1992, 191, 57–72. [Google Scholar]
- Rogers, L.J.; Andrew, R.J. Comparative Vertebrate Lateralization; Cambridge University Press: London, UK, 2002; ISBN 0-521-78161-2. [Google Scholar]
- Ströckens, F.; Gunturkün, O.; Ocklenburg, S. Limb preferences in non-human vertebrates. Laterality Asymmetries Body Brain Cogn. 2013, 18, 536–575. [Google Scholar] [CrossRef]
- Ghirlanda, S.; Vallortigara, G. The evolution of brain lateralization: A game-theoretical analysis of population structure. Proc. R. Soc. Lond. B 2004, 271, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Ghirlanda, S.; Frasnelli, E.; Vallortigara, G. Intraspecific competition and coordination in the evolution of lateralization. Philos. Tranzactions R. Soc. B 2009, 364, 861–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallortigara, G. The Evolutionary psychology of left and right: Costs and benefits of lateralization. Dev. Phychobiol. 2006, 48, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Hori, M. Frequency-dependent natural selection in the handedness of scale-eating cichlid fish. Science 1993, 260, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Liem, K.F.; Stewart, D.J. Evolution of the scale-eating cichlid fishes of Lake Tanganyika: A generic revision with a description of a new species. Bull. Mus. Comp. Zool. 1976, 147, 319–350. [Google Scholar]
- Hori, M. Feeding relationships among cichlid fishes in Lake Tanganyika: Effects of intra- and interspecific variation of feeding behavior on their coexistence. Ecol. Int. Bull. 1991, 19, 89–101. [Google Scholar]
- Takahashi, R.; Moriwaki, T.; Hori, M. Foraging behaviour and functional morphology of two scale-eating cichlids from Lake Tanganyika. J. Fish Biol. 2007, 70, 1458–1469. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Hori, M.; Oda, Y. Lateralized kinematics of predation behavior in a Lake Tanganyika scale-eating cichlid fish. PLoS ONE 2012, 7, e29272. [Google Scholar] [CrossRef] [PubMed]
- Hata, H.; Yasugi, M.; Hori, M. Jaw laterality and related handedness in the hunting behavior of a scale-eating characin, Exodon paradoxus. PLoS ONE 2011, 6, e29349. [Google Scholar] [CrossRef]
- Hori, M.; Ochi, H.; Kohda, M. Inheritance pattern of lateral dimorphism in two cichlids (a scale eater, Perissodus microlepis, and an herbivore, Neolamprologus moorii) in Lake Tanganyika. Zool. Sci. 2007, 24, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Stewart, T.A.; Albertson, R.C. Evolution of a unique predatory feeding apparatus: Functional anatomy, development and a genetic locus for jaw laterality in Lake Tanganyika scale-eating cichlids. BMC Biol. 2010, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Hata, H.; Takahashi, R.; Ashiwa, H.; Awata, S.; Takeyama, T.; Kohda, M.; Hori, M. Inheritance patterns of lateral dimorphism examined by breeding experiments with the Tanganyikan cichlid (Julidochromis transcriptus) and the Japanese medaka (Oryzias latipes). Zool. Sci. 2012, 29, 49–53. [Google Scholar] [CrossRef]
- Hata, H.; Hori, M. Inheritance patterns of morphological laterality in mouth opening of zebrafish, Danio rerio. Laterality 2012, 17, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Hori, M. Unstable evolutionarily stable strategy and oscillation: A model on lateral asymmetry in scale-eating cichlids. Am. Nat. 1994, 144, 1001–1020. [Google Scholar] [CrossRef]
- Takahashi, S.; Hori, M. Oscillation maintains polymorphisms—A model of lateral asymmetry in two competing scale-eating cichlids. J. Theor. Biol. 1998, 195, 1–12. [Google Scholar] [CrossRef]
- Hori, M. Structure of littoral fish communities organized by their feeding activities. In Fish Communities in Lake Tanganyika; Kawanabe, H., Hori, M., Nagoshi, M., Eds.; Kyoto University Press: Kyoto, Japan, 1997; pp. 275–298. ISBN 4-87698-042-X. [Google Scholar]
- Takeuchi, Y.; Ochi, H.; Kohda, M.; Sinyinza, D.; Hori, M. A 20-year census of a rocky littoral fish community in Lake Tanganyika. Ecol. Freshw. Fish 2010, 19, 239–248. [Google Scholar] [CrossRef]
- Takahashi, R.; Watanabe, K.; Nishida, M.; Hori, M. Evolution of feeding secialization in Tanganyikan scale-eating cichlids: A molecular phylogenetic approach. BMC Evol. 2007, 7, 195. [Google Scholar]
- Hori, M. Mutualism and commensalism in the fish community of Lake Tanganyika. In Evolution and Coadaptation in Biotic Communities; Kawano, S., Connell, J.H., Hidaka, T., Eds.; Tokyo University Press: Tokyo, Japan, 1987; pp. 219–239. [Google Scholar]
- Hori, M.; Nakajima, M.; Hata, H.; Yasugi, M.; Takahashi, S.; Nakae, M.; Yamaoka, K.; Kohda, M.; Kitamura, J.; Maehata, M.; et al. Laterality is universal among fishes but increasingly cryptic among derived groups. Zool. Sci. 2017, 34, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Yodo, T.; Katano, O. Righty fish are hooked on the right side of their mouths—Observations from an angling experiment with largemouth bass, Microperus salmoides. Zool. Sci. 2007, 24, 855–859. [Google Scholar] [CrossRef]
- Seki, S.; Kohda, M.; Hori, M. Asymmetry of mouth morph of a freshwater goby, Rhinogobius flumineus. Zool. Sci. 2000, 17, 1321–1325. [Google Scholar] [CrossRef]
- Matsui, S.; Takeuchi, Y.; Hori, M. Relation between morphological antisymmetry and behavioral laterality in a Poeciliid fish. Zool. Sci. 2013, 30, 613–618. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Hori, M. Behavioural laterality in the shrimp-eating cichlid fish, Neolamprologus fasciatus, in Lake Tanganyika. Anim. Behav. 2008, 75, 1359–1366. [Google Scholar] [CrossRef]
- Yasugi, M.; Hori, M. Predominance of cross-predation between lateral morphs in a largemouth bass and a freshwater goby. Zool. Sci. 2011, 28, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Yasugi, M.; Hori, M. Lateralized behavior in the attacks of largemouth bass on Rhinogobius gobies corresponding to their morphological antisymmetry. J. Exp. Biol. 2012, 215, 2390–2398. [Google Scholar] [CrossRef] [PubMed]
- Yasugi, M.; Hori, M. Predominance of parallel- and cross-predation in anglerfish. Mar. Ecol. 2016, 37, 576–587. [Google Scholar] [CrossRef]
- Hori, M.; Yamaoka, K.; Takamura, K. Abundance and micro-distribution of cichlid fishes on a rocky shore of Lake Tanganyika. Afr. Study Monogr. 1983, 3, 25–38. [Google Scholar]
- Hata, H.; Yasugi, M.; Takeuchi, Y.; Hori, M. Distinct lateral dimorphism in the jaw morphology of the scale-eating cichlids, Perissodus microlepis and P. straeleni. Ecol. Evol. 2013, 3, 4641–4647. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Hori, M.; Myint, O.; Kohda, M. Lateral bias of agonistic responses to mirror images and morphological asymmetry in the Siamese fighting fish (Betta splendens). Behav. Brain Res. 2010, 208, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Hori, M. Coexistence of competing species by the oscillation polymorphism. J. Theor. Biol. 2005, 235, 591–596. [Google Scholar] [CrossRef]
- Nakajima, M.; Matsuda, H.; Hori, M. A population genetic model for lateral dimorphism frequency in fishes. Am. Nat. 2005, 163, 692–698. [Google Scholar] [CrossRef]
- Nakajima, M.; Matsuda, H.; Hori, M. Persistence and fluctuation of lateral dimorphism in fishes. Am. Nat. 2004, 163, 692–698. [Google Scholar] [CrossRef]
Figure 1.
The laterality of two scale-eating cichlids, Perissodus straeleni (top) and Perissodus microlepis (bottom) in Lake Tanganyika. A lefty morph of the former and righty morph of the latter species are shown from both sides.
Figure 1.
The laterality of two scale-eating cichlids, Perissodus straeleni (top) and Perissodus microlepis (bottom) in Lake Tanganyika. A lefty morph of the former and righty morph of the latter species are shown from both sides.

Figure 2.
The temporal changes in the ratios of laterality (frequency of righty morphs) in two scale-eaters, P. microlepis and P. straeleni, on the southernmost shoreline of Lake Tanganyika over a 30-year period from 1988 to 2017.
Figure 2.
The temporal changes in the ratios of laterality (frequency of righty morphs) in two scale-eaters, P. microlepis and P. straeleni, on the southernmost shoreline of Lake Tanganyika over a 30-year period from 1988 to 2017.

Figure 3.
The relationship of laterality ratios between P. microlepis and P. straeleni. Red lines indicate that the change proceeds anti-clockwise, thus the change in P. microlepis follows that in P. straeleni. Blue lines proceed clockwise, indicating the opposite pattern.
Figure 3.
The relationship of laterality ratios between P. microlepis and P. straeleni. Red lines indicate that the change proceeds anti-clockwise, thus the change in P. microlepis follows that in P. straeleni. Blue lines proceed clockwise, indicating the opposite pattern.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The combination of morph types between each scale-eater and its victim fish. R and L represent the righty morph and lefty morph, respectively.
Table 1.
The combination of morph types between each scale-eater and its victim fish. R and L represent the righty morph and lefty morph, respectively.
| Species | Scale-Eater | Victim Fish | Species | Scale-Eater | Victim Fish | ||
|---|---|---|---|---|---|---|---|
| R | L | R | L | ||||
| P. microlepis | R | 3 | 19 | P. straeleni | R | 4 | 21 |
| L | 25 | 6 | L | 22 | 4 | ||
| Total: 53 | Total: 51 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hori, M.; Kohda, M.; Awata, S.; Takahashi, S. Dynamics of Laterality in Lake Tanganyika Scale-Eaters Driven by Cross-Predation. Symmetry 2019, 11, 119. https://0-doi-org.brum.beds.ac.uk/10.3390/sym11010119
AMA Style
Hori M, Kohda M, Awata S, Takahashi S. Dynamics of Laterality in Lake Tanganyika Scale-Eaters Driven by Cross-Predation. Symmetry. 2019; 11(1):119. https://0-doi-org.brum.beds.ac.uk/10.3390/sym11010119
Chicago/Turabian StyleHori, Michio, Masanori Kohda, Satoshi Awata, and Satoshi Takahashi. 2019. "Dynamics of Laterality in Lake Tanganyika Scale-Eaters Driven by Cross-Predation" Symmetry 11, no. 1: 119. https://0-doi-org.brum.beds.ac.uk/10.3390/sym11010119
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.