
Zinc Oxide Nanoparticles Induce DNA Damage in Sand Dollar Scaphechinus mirabilis Sperm
by
 and
and
Sergey Petrovich Kukla
*, 
Victor Pavlovich Chelomin
,
Andrey Alexandrovich Mazur
and
Valentina Vladimirovna Slobodskova
Il’ichev Pacific Oceanological Institute, Far Eastern Branch, Russian Academy of Sciences, 690041 Vladivostok, Russia
*
Author to whom correspondence should be addressed.
Toxics 2022, 10(7), 348; https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10070348
Submission received: 25 May 2022
/
Revised: 14 June 2022
/
Accepted: 23 June 2022
/
Published: 24 June 2022
(This article belongs to the Special Issue Current Knowledge of Nanomaterial Ecotoxicology)
Abstract
:Products containing nanomaterials are becoming more and more common in everyday life. Zinc oxide nanoparticles (ZnO NPs), meanwhile, are among the most widely used NPs. However, their genotoxic effect on the germ products of marine organisms is poorly understood. Therefore, the effects of ZnO NPs and zinc ions (20, 50, 100, 200 µg/L) on the sperm of sand dollar Scaphechinus mirabilis were compared. Comet assay showed that both tested pollutants caused an increase in DNA damage to 6.57 ± 2.41 and 7.42 ± 0.88% DNA in the comet tail, for zinc ions and ZnO NPs, respectively. Additionally, a different pattern was shown by the increase in DNA damage, with increasing concentration of pollutants, in different experimental groups.
1. Introduction
A huge variety of nanomaterials have now been developed and are widely used in various industries (electronics, energy, construction, semiconductors, paint and varnish, household chemicals, etc.) and their number continues to grow rapidly [1,2,3]. The extent of nanomaterial penetration in consumer products widely used by humans in everyday life is particularly impressive: in medicine, clothing, detergents, perfumes, cosmetics, household items, as well as in food and personal care products [4,5,6].
The growing interest in nanoparticles (NPs) is due to the manifestation of new, unique physical and chemical properties in the transition of traditional materials to the nanodispersed state [7]. This makes scientists pay special attention to the ecotoxicological effects associated with NPs. It is these properties that make NPs highly reactive, forming bonds of a different nature with basic biostructures and cellular macromolecules, which may result in unique pathobiochemical effects. Current nanotoxicological research shows that NPs of a different nature are capable of crossing the body’s protective barriers: the gastric, placental and blood-brain barriers [8,9,10], penetrating biological membranes and accumulating in biological systems of different levels of organization [8,11,12,13]. This shows that complex ecotoxicological problems associated with the introduction of nanotechnology products into the biosphere and the danger of different composition NPs interfering with biochemical processes in the living organism are inevitable.
Products of nanotechnology inevitably enter the environment in different ways: through production, processing, transport, use and disposal. In doing so, they inevitably end up in coastal ecosystems due to various migration processes [14,15]. Therefore, not only humans, but also the environment with all of its biodiversity, are becoming increasingly influenced by NPs.
The penetration of NPs into the marine environment is fraught with numerous consequences which, due to lack of information, cannot yet be predicted. Despite the low water solubility and tendency for aggregation of NPs, particularly metal oxides [16], which to some extent limits their accessibility to marine organisms, experimental data indicate high levels of NPs accumulation in various species of marine organisms, such as crustaceans, echinoderms and mollusks [14,17,18].
Among a wide range of metal oxide NPs, ZnO NPs are considered to be among the most common used [19]. Due to their unique characteristics, ZnO NPs are widely used in instrumentation (including bioelectronics and biomedical devices), cosmetics, pharmaceuticals, UV filters, biomaterials and food packaging materials.
In the literature, there are sufficiently convincing data demonstrating the effect of ZnO NPs on various functional aspects of marine organisms [20,21,22]. A number of studies have revealed a higher toxicity of ZnO NPs, not only in relation to those that are identical in size to particles of oxides of other metals [23,24], but also in comparison with zinc ions [20,25]. Of increased concern to ecotoxicologists is the ability of ZnO NPs to induce genome damage, manifested in impaired gene expression in mussels [26], and in chromosomal aberrations and morphological changes in the gametes and embryos of sea urchins Paracentrotus lividus [20,27]. In addition, in the example of the sea urchin Scaphechinus mirabilis, the exposure of adults to ZnO NPs has been shown to affect the reproductive system, causing DNA damage in gametes and abnormalities in larval development [25,28].
The increased ecotoxicological interest in the early stages of development is due to the generally accepted view in the literature that gametes, embryos and larvae are more sensitive than adults and that they represent a critical period in the life cycle of an organism [29,30]. Spermatozoa are highly specialized cells and, unlike somatic cells, they are potentially more susceptible to damage by substances exhibiting genotoxic properties, as they contain highly condensed DNA, have weak antioxidant protection and very limited ability to repair DNA damage [31,32,33,34]. Therefore, spermatozoa are very vulnerable to oxidative stress, which is known to be one of the main mechanisms involved in DNA damage.
S. mirabilis, as a typical member of the sea urchin class, releases sexual gametes directly into the sea water during spawning, where fertilization and further development of embryos and larvae occurs. At this stage, spermatozoa are the least protected and their outer membranes and receptors are directly exposed to a wide range of chemicals [35,36]. In this respect, our experiments mimic environmental conditions in terms of Zn2+ and ZnO NPs interaction with sea dollar gametes. Our studies were carried out under controlled laboratory conditions with no exposure to other concomitant stressors typical of the marine environment. Zn2+ and ZnO NPs concentrations, which are widely used in ecotoxicological experiments, were used [37]. This approach, which belongs to the category of “acute” experiments, aims to identify likely “targets”, i.e., most vulnerable cell structures, and associated toxicity mechanisms. In order to assess the potential danger posed by the penetration of nanoparticles into the marine environment, we investigated the genotoxic properties of NPs in male sea urchin gametes.
To determine genotoxicity, we applied the comet assay method, a sensitive method for the early detection of DNA damage [38], which is widely used in research on the toxicity of NPs [39].
The relevance of assessing DNA damage in sperm cells increases dramatically, given that DNA damage to these cells typically has deleterious effects on species reproduction (reproductive success), which is an important indicator of long-term ecotoxicological consequences [31].
Based on the above, the aim of this work was to investigate, using the spermatozoa of the sea urchin S. mirabilis, the potential risk posed by ZnO NPs to the genome integrity of marine invertebrate gametes.
2. Materials and Methods
2.1. Preparation of Working Solutions
2.2. Description of the Experiment
Adult sea dollars were collected in Peter the Great Gulf, Japan Sea and delivered to the laboratory within one hour. After delivery, the urchins were acclimatized for 2 days in water filtered by a three-stage gravel filter and sterilized by ultraviolet (pH 8.2; salinity 32.75 ppm, O2 concentration 7.5 ± 0.3 mg/L, T = 17–18 °C).
Semen from 4 males S. mirabilis in 2 replicates was used in an experiment. Spermatozoa were obtained by stimulating spawning with 0.5 M potassium chloride solution. Semen was collected immediately before the experiment and diluted with pure seawater. Both experimental and control groups used semen from the same males.
To study genotoxicity, experimental solutions were added to the diluted semen aliquot to final concentrations of 20, 50, 100 and 200 µg Zn/L and incubated for 1 h [41]. After that, it was used in the comet assay.
2.3. Comet Assay
An alkaline version of the comet assay adapted for marine organisms was used [42].
First, 50 µL of semen suspension was added to 100 µL of 1% fusible agarose in 0.04 M phosphate buffer (pH 7.4) at 37 °C, thoroughly mixed, and applied on a slide coated with 1% agarose solution for better adhesion and covered with a coverslip. The sample was placed in the refrigerator for 3 min to form a gel. The coverslip was carefully removed and the slide was submerged into the lysis solution (2.5M NaCl; 0.1M EDTA-Na2, 1% Triton X-100; 10% DMSO; 0.02 M Tris, pH 10) for 1 h in the dark at 4 °C. After washing with distilled water, the slides were placed in electrophoresis buffer (300 mM NaOH, 1 mM EDTA-Na2) and incubated for 40 min. Electrophoresis was performed at 2 V/cm for 15 min. After neutralization (0.4 M Tris-HCl, pH 7.4), the slides were stained with SYBR Green fluorescent dye.
DNA comets were visualized and recorded using a fluorescence microscope (Zeiss, Axio Imager A1) equipped with an AxioCam MRc digital camera. For digital image processing, the CaspLab computer program was used to calculate various comet parameters indicating the degree of cellular DNA damage. At least 50 comets were analyzed for each glass. For each comet, the proportion of DNA in the comet tail (% of DNA in tail) was determined.
2.4. Statistical Analysis
Statistical processing of the results was performed using STATISTICA 8 software (StatSoft, Tulsa, OK, USA). The significance of the differences between the control and experimental groups was assessed by means of a one-factor analysis of variance using Dunnett’s test (at p ≤ 0.05).
3. Results and Discussion
Evaluation of DNA damage after exposure of experimental solutions gave the following results.
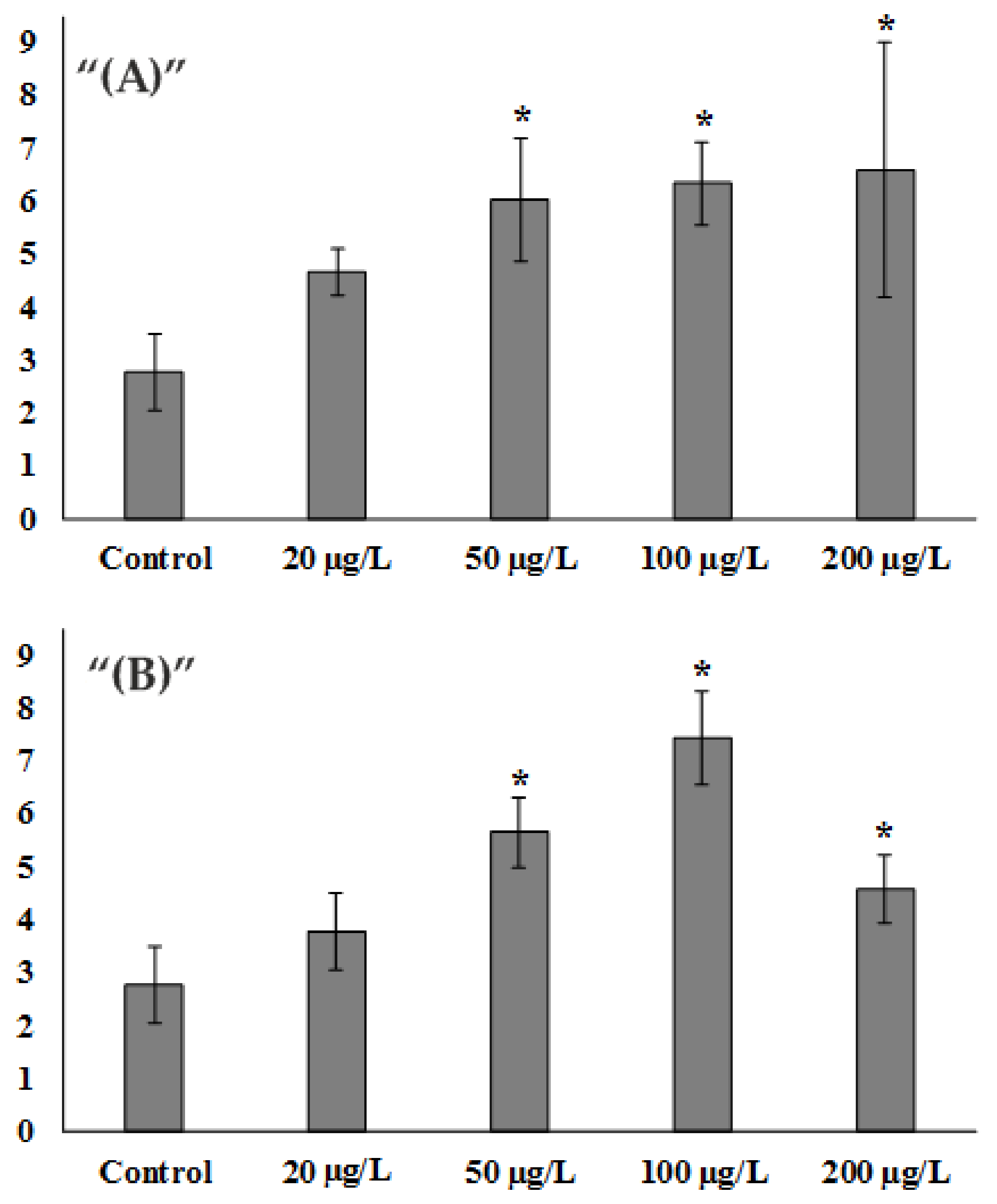
After experimental exposure to Zn2+, already at the concentration of 20 μg/L, there was an increase in DNA damage by about 1.6 times which amounted to 4.68 ± 0.44% DNA in the tail, compared to the control group, where the damage percentage was 2.77 ± 0.73% DNA in the tail. At concentrations of 50 µg/L and higher, a statistically significant 2.1-fold increase in DNA damage was observed, corresponding to 6.04 ± 1.17% DNA in the tail. At the same time, a further increase in zinc ions in water up to the values of 100 µg/L and 200 µg/L did not lead to a further sharp increase in DNA damage and exceeded the control value by 2.2- and 2.3-fold, respectively (6.33 ± 0.78 6.57 ± 2.41% DNA in the tail) (Figure 1A).
When exposed to low concentrations of ZnO NPs, a similar pattern to that of zinc ions was observed. Exposure to 20 µg/L resulted in a slight increase in DNA damage of approximately 1.3-fold and amounted to 3.77 ± 0.74% DNA in the tail. At 50 µg/L, the difference was more than 2-fold and amounted to 5.65 ± 0.66% DNA in the tail. At 100 µg/L there was a further 2.6-fold increase in DNA damage to values of 7.42 ± 0.88% DNA in the tail. Nevertheless, at the NPs concentration of 200 μg/L, there was a decrease in the extent of DNA damage. It was only 1.6-fold higher than the control and amounted to 4.59 ± 0.66% DNA in the tail (Figure 1B).
In order to estimate the size of the detected destructive changes in sperm exposed to ZnO NPs, one can refer to the results of Lacaze et al. (2011) [32]. Using healthy amphipods of Gammarus fossarum as an example, they determined the reference and threshold values of sperm genome damage with regard to seasonal variations. The reference value of DNA damage in sperm was found to be 3.1%, while the minimum and maximum threshold values were 2.6% and 3.5%, respectively [32].
Comparison of these data with our results obtained on sand dollar sperm suggests that exposure to ZnO NPs leads to severe damage in the sperm DNA molecule at all concentrations studied.
Literature data on the genotoxic properties of NPs are still scarce. Similar studies using sea urchin sperm of Paracentrotus lividus have also revealed genotoxic properties of ZnO and CuO NPs [27,43]. In addition, the genome sensitivity of male gametes to various nanoparticles has been shown in other marine invertebrates, including the bivalve Tegillarca granosa [44], the polychaete Hydroides elegans [45] and the ascidia Ciona intestinalis [46].
In explaining the mechanisms of toxicity, the view has developed and become popular that the toxicity of NPs is directly related to Zn2+, which is formed due to the instability of NPs in the aqueous medium [47]. However, given the short-term nature of experiments in this research, it is logical to assume that most of the ZnO NPs retain their structure and the Zn2+ concentration is very low [47]. Thus, the main contribution to DNA degradation is made by NPs themselves. In addition, several previous studies have revealed differences in ecotoxicological effects initiated by NPs and zinc ions [20,28]. Oliviero et al. (2019) drew attention to the lack of a direct correlation between levels of sperm DNA damage and NPs concentration [27]. They noted that, in a certain range of changes in ZnO NP concentrations, a gradual increase in the number of damaged “comets” was observed, reaching a maximum value (3 μM, in their case), but with a further increase in concentration, this rate began to decrease gradually, in contrast to exposure to dissolved zinc. Additionally, a similar dependence was observed in the analysis of the number of skeletal anomalies in sea urchin larvae of P. lividus after exposure to different concentrations of TiO2 NPs [48]. This pattern in our results and those reported in the literature can be explained by the fact that the aggregation rate of NPs depends on their concentration in the aqueous medium [49]. At higher concentrations, nanoparticles, particularly ZnO NPs, tend to form large aggregates, which become less bioavailable and, consequently, cause less effect.
The mechanisms underlying the genotoxic effects of ZnO nanoparticles on male gametes are not clear. Most researchers, in explaining the causes of genotoxicity, draw attention to the ability of ZnO to induce increased generation of reactive oxygen species (ROS), thereby causing oxidative stress. Highly reactive oxyradicals are thought to be the main cause of oxidative damage and DNA strand breaks [11]. Additionally, the possibility of NPs directly affecting gene structures and their regulation mechanisms cannot be ruled out [50].
It has been established that the main modes of entry of NPs into a living cell are variants of endocytosis, in particular through receptor-mediated pinocytosis [51]. At present, there are no data on the penetration of NPs into sperm cells. Given the adsorption characteristics of NPs and their ability to interact with cellular receptors responsible for the transmission of stress signals [6], the following suggestion can be made. In the framework of oxidative stress, it can be suggested that ZnO NPs, when located at different sites of the outer sperm membrane, can disorganize the receptor-signaling system to some extent and induce the formation of ROS. As shown earlier, sea urchin spermatozoa are capable of generating several types of ROS, such as H2O2 and O2− [52]. It is likely that a similar mechanism of ROS formation was initiated through the exposure of P. lividus sea urchin sperm to CuO NPs, causing significant DNA fragmentation [46]. Additionally, was showed that TiO2 NPs, when interacting with gills, caused oxidative stress without penetrating cells [53].
In the context of the problem at hand, it must be taken into account that spermatozoa and oocytes have a unique and essential biological function in forming the genome for the development of the next generation. Therefore, the integrity of gamete genomes is of paramount importance for the development of viable offspring.
However, the biochemical response of the organism, such as DNA damage, most often outpaces the cytological response. For example, short-term exposure of bivalve spermatozoa to benz[a]pyrene and diuron resulted in significant and dose-dependent DNA damage, but the sperm retained fertilizing capacity [31,54]. Additionally, in other studies, despite the high level of DNA damage in fish sperm induced by exposure to genotoxicants, fertilization success was maintained at a high level [34,55,56].
It is thought that, after fertilization, damaged paternal DNA may be partially repaired by the embryo repair system [34,57], or serve as a signal to trigger various developmental anomalies and embryonic deaths.
Indirect support for this assumption is provided by numerous laboratory studies in recent years that have demonstrated a relationship between sperm DNA integrity and offspring quality in aquatic organisms [31,32,55,56,57]. For example, rainbow trout eggs fertilized with sperm with varying degrees of fragmented DNA formed embryos with lower chances of survival [57]. Numerous malformations, mainly in skeletal development, were observed in the hatching larvae of the three-legged stickleback, grown from eggs fertilized with methylmethanesulfonate-treated spermatozoa [56].
In conclusion, we believe that despite the relatively low degree of DNA molecule destruction in spermatozoa detected in our experiments, the risk of further initiation of destructive processes and manifestation of long-term undesirable effects remains. Further research in this area should focus on a detailed study of the biochemical mechanisms involved in sperm DNA damage.
Author Contributions
Conceptualization, V.P.C., V.V.S.; methodology, A.A.M., V.V.S. and S.P.K.; soft-ware, S.P.K.; validation, A.A.M. and V.V.S.; formal analysis, S.P.K.; investigation, A.A.M., V.V.S. and S.P.K.; resources, V.P.C.; data curation, V.P.C. and V.V.S. verified; writing—original draft preparation, V.P.C.; writing—review and editing, V.P.C., A.A.M. and V.V.S.; visualization, A.A.M.; supervision, V.P.C.; project administration, V.P.C.; funding acquisition, V.P.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the State assignment for research work of V.I. Il’ichev Pacific Oceanological Institute, FEB RAS (No. 121-21500052-9).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Elena Vladimirovna Zhuravel, at the Far Eastern Federal University, for her help in keeping and spawning the sand dollar.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giese, B.; Klaessig, F.; Park, B.; Kaegi, R.; Steinfeldt, M.; Wigger, H.; von Gleich, A.; Gottschalk, F. Risks, release and concentrations of engineered nanomaterial in the environment. Sci. Rep. 2018, 8, 1565. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, H.M.R.; Dawood, M.A.O.; Menanteau-Ledouble, S.; El-Matbouli, M. Environmental transformation of n-TiO 2 in the aquatic systems and their ecotoxicity in bivalve mollusks: A systematic review. Ecotoxicol. Environ. Saf. 2020, 200, 110776. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, A.; Liang, L.; Lee, M.Y.; Bobacka, J.; Lisak, G. Too small to matter? Physicochemical transformation and toxicity of engineered nTiO 2, nSiO 2, nZnO, carbon nanotubes, and nAg. J. Hazard. Mater. 2021, 404 Pt A, 124107. [Google Scholar] [CrossRef]
- Mueller, N.; Nowack, B. Exposure modeling of engineered nanoparticles in the environment. Environ. Sci. Technol. 2008, 42, 4447–4453. [Google Scholar] [CrossRef]
- Garcia, C.V.; Shin, G.H.; Kim, J.T. Metal oxide-based nanocomposites in food packaging: Applications, migration, and regulations. Trends Food Sci. Technol. 2018, 82, 21–31. [Google Scholar] [CrossRef]
- Sruthi, S.; Ashtami, J.; Mohanan, P.V. Biomedical application and hidden toxicity of Zinc oxide nanoparticles. Mater. Today Chem. 2018, 10, 175–186. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Suh, W.H.; Suslick, K.S.; Stucky, G.D.; Suh, Y.H. Nanotechnology, nanotoxicology, and neuroscience. Prog. Neurobiol. 2009, 87, 133–170. [Google Scholar] [CrossRef] [Green Version]
- Sawicki, K.; Czajka, M.; Matysiak-Kucharek, M.; Fal, B.; Drop, B.; Męczyńska-Wielgosz, S.; Sikorska, K.; Kruszewski, M.; Kapka-Skrzypczak, L. Toxicity of metallic nanoparticles in the central nervous system. Nanotechnol. Rev. 2019, 8, 175–200. [Google Scholar] [CrossRef] [Green Version]
- Bongaerts, E.; Nawrot, T.S.; van Pee, T.; Ameloot, M.; Bové, H. Translocation of (ultra)fine particles and nanoparticles across the placenta; a systematic review on the evidence of in vitro, ex vivo, and in vivo studies. Part. Fibre Toxicol. 2020, 17, 56. [Google Scholar] [CrossRef]
- Arora, S.; Rajwade, J.M.; Paknikar, K.M. Nanotoxicology and in vitro studies: The need of the hour. Toxicol. Appl. Pharmacol. 2012, 258, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, O.; Juganson, K.; Ivask, A.; Kasemets, K.; Mortimer, M.; Kahru, A. Toxicity of Ag, CuO and ZnO nanoparticles to selected environmentally relevant test organisms and mammalian cells in vitro: A critical review. Arch. Toxicol. 2013, 87, 1181–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, M.N. Do nanoparticles present ecotoxicological risks for the health of the aquatic environment? Environ. Int. 2006, 32, 967–976. [Google Scholar] [CrossRef]
- Matranga, V.; Corsi, I. Toxic effects of engineered nanoparticles in the marine environment: Model organisms and molecular approaches. Mar. Environ. Res. 2012, 76, 32–40. [Google Scholar] [CrossRef]
- Keller, A.A.; Wang, H.; Zhou, D.; Lenihan, H.S.; Cherr, G.; Cardinale, B.J.; Miller, R.; Ji, Z. Stability and aggregation of metal oxide nanoparticles in natural aqueous matrices. Environ. Sci. Technol. 2010, 44, 1962–1967. [Google Scholar] [CrossRef]
- Canesi, L.; Ciacci, C.; Fabbri, R.; Marcomini, A.; Pojana, G.; Gallo, G. Bivalve mollusks as a unique target group for nanotoxity. Mar. Environ. Res. 2012, 76, 16–21. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Z.; Zhao, J.; Lin, M.; Xing, B. Accumulation of metal-based nanoparticles in marine bivalve mollusks from offshore aquaculture as detected by single particle ICP-MS. Environ. Pollut. 2020, 260, 114043. [Google Scholar] [CrossRef]
- Paul, S.K.; Dutta, H.; Sarkar, S.; Sethi, L.N.; Ghosh, S.K. Nanosized zinc oxide: Super-functionalities, present scenario of application, safety issues, and future prospects in food processing and allied industries. Food Rev. Int. 2019, 35, 505–535. [Google Scholar] [CrossRef]
- Manzo, S.; Miglietta, M.; Rametta, G.; Buono, S.; Francia, G. Embryotoxicity and spermiotoxicity of nanosized ZnO for Mediterranean sea urchin Paracentrotus lividus. J. Hazard. Mater. 2013, 254, 1–9. [Google Scholar] [CrossRef]
- Trevisan, R.; Delapedra, G.; Mello, D.F.; Arl, M.; Schmidt, É.C.; Meder, F.; Monopoli, M.; Cargnin-Ferreira, E.; Bouzon, Z.L.; Fisher, A.S.; et al. Gills are an initial target of zinc oxide nanoparticles in oysters Crassostrea gigas, leading to mitochondrial disruption and oxidative stress. Aquat. Toxicol. 2014, 153, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Prato, E.; Fabbrocini, A.; Libralato, G.; Migliore, L.; Parlapiano, I.; D’Adamo, R.; Rotini, A.; Manfra, L.; Lofrano, G.; Carraturo, F.; et al. Comparative toxicity of ionic and nanoparticulate zinc in the species Cymodoce truncata, Gammarus aequicauda and Paracentrotus lividus. Environ. Sci. Pollut. Res. Int. 2021, 28, 42891–42900. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhu, L.; Duan, Z. Comparative toxicity of several metal oxide nanoparticle aqueous suspensions to Zebrafish (Danio rerio) early developmental stage. J. Environ. Sci. Health. A Tox. Hazard. Subst. Environ. Eng. 2008, 43, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Minetto, D.; Ghirardini, A.V.; Libralato, G. Saltwater ecotoxicology of Ag, Au, CuO, TiO2, ZnO and C60 engineered nanoparticles: An overview. Environ. Int. 2016, 92–93, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, A.A.; Zhuravel, E.V.; Slobodskova, V.V.; Mazur, M.A. Assessment of the toxic effect of zinc ions and nano-sized zinc oxide on the early development of the sand dollar Scaphechinus mirabilis (Agassiz, 1864) (Echinodermata: Echinoidea). Russ. J. Mar. Bio. 2020, 46, 49–55. [Google Scholar] [CrossRef]
- Li, J.; Schiavo, S.; Xiangli, D.; Rametta, G.; Miglietta, M.L.; Oliviero, M.; Changwen, W.; Manzo, S. Early ecotoxic effects of ZnO nanoparticle chronic exposure in Mytilus galloprovincialis revealed by transcription of apoptosis and antioxidant-related genes. Ecotoxicology 2018, 27, 369–384. [Google Scholar] [CrossRef]
- Oliviero, M.; Schiavo, S.; Dumontet, S.; Manzo, S. DNA damages and offspring quality in sea urchin Paracentrotus lividus sperms exposed to ZnO nanoparticles. Sci. Total Environ. 2019, 651, 756–765. [Google Scholar] [CrossRef]
- Mazur, A.A.; Zhuravel, E.V.; Slobodskova, V.V.; Mazur, M.A.; Kukla, S.P.; Chelomin, V.P. Waterborne exposure of adult sand dollar, Scaphechinus mirabilis (Agassiz, 1864), to zinc ions and zinc oxide nanoparticles affects early development of its offspring. Water Air Soil Pollut. 2020, 231, 115. [Google Scholar] [CrossRef]
- Beiras, R.; Durán, I.; Bellas, J.; Sánchez-Marín, P. Biological Effects of Contaminants: Paracentrotus Lividus Sea Urchin Embryo Test with Marine Sediment Elutriates; International Council for the Exploration of the Sea (ICES): Copenhagen, Denmark, 2012; p. 13. [CrossRef]
- Nobre, C.R.; Santana, M.F.M.; Maluf, A.; Cortez, F.S.; Cesar, A.; Pereira, C.D.S.; Turra, A. Assessment of microplastic toxicity to embryonic development of the sea urchin Lytechinus variegatus (Echinodermata: Echinoidea). Mar. Pollut. Bull. 2015, 92, 99–104. [Google Scholar] [CrossRef]
- Lewis, C.; Galloway, T.S. Genotoxic damage in Polychaetes: A study of species and cell-type sensitivities. Mutat. Res. Genet. Toxicol. Environ. Mutat. 2008, 654, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Lacaze, E.; Geffard, O.; Goyet, D.; Bony, S.; Devaux, A. Linking genotoxic responses in Gammarus fossarum germ cells with reproduction impairment, using the Comet assay. Environ. Res. 2011, 111, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Lacaze, E.; Geffard, O.; Bony, S.; Devaux, A. Genotoxicity assessment in the amphipod Gammarus fossarum by use of the alkaline Comet assay. Mutat. Res. 2010, 700, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Devaux, Y.; Zangrando, J.; Schroen, B.; Creemers, E.E.; Pedrazzini, T.; Chang, C.P.; Dorn, G.W.; Thum, T.; Heymans, S. Cardiolinc Network. Long noncoding RNAs in cardiac development and ageing. Nat. Rev. Cardiol. 2015, 12, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Hamlin, H.J.; Marciano, K.; Downs, C.A. Migration of nonylphenol from food-grade plastic is toxic to the coralreef fish species Pseudochromis fridmani. Chemosphere 2015, 139, 223–228. [Google Scholar] [CrossRef]
- Messinetti, S.; Mercurio, S.; Parolini, M.; Sugni, M.; Pennati, R. Effects of polystyrene microplastics on early stages of two marine invertebrates with different feeding strategies. Environ. Pollut. 2018, 237, 1080–1087. [Google Scholar] [CrossRef]
- Balbi, T.; Camisassi, G.; Montagna, M.; Fabbri, R.; Franzellitti, S.; Carbone, C. Impact of cationic polystyrene nanoparticles (PS-NH2) on early embryo development of Mytilus galloprovincialis: Effects on shell formation. Chemosphere 2017, 186, 1–9. [Google Scholar] [CrossRef]
- Smith, M.A.; Fernandez-Triana, J.; Roughley, R.; Hebert, D.N. DNA barcode accumulation curves for understudied taxa and areas. Mol. Ecol. Resour. 2009, 9, 208–216. [Google Scholar] [CrossRef]
- Mahaye, N.; Thwala, M.; Cowan, D.A.; Musee, N. Genotoxicity of metal based engineered nanoparticles in aquatic organisms: A review. Mutat. Res. 2017, 773, 134–160. [Google Scholar] [CrossRef]
- Tang, Y.; Xin, H.; Yang, S.; Guo, M.; Malkoske, T.; Yin, D.; Xia, S. Environmental risks of ZnO nanoparticle exposure on Microcystis aeruginosa: Toxic effects and environmental feedback. Aquat. Toxicol. 2018, 204, 19–26. [Google Scholar] [CrossRef]
- Dinnel, P.A.; Stober, Q.J.; Crumley, S.C.; Nakatani, R.E. Development of a sperm cell toxicity test for marine water. Aquat. Toxicol. Haz. Asses. 1982, 1, 82–98. [Google Scholar] [CrossRef]
- Mitchelmore, C.L.; Birmelin, C.; Livingstone, D.R.; Chipman, J.K. Detection of DNA strand breaks in isolated mussels (Mytilus edulis) digestive gland cells using the “comet” assay. Ecotoxicol. Environ. Saf. 1998, 41, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Boni, R.; Buttino, I.; Tosti, E. Spermiotoxicity of nickel nanoparticles in the marine invertebrate Ciona intestinalis (ascidians). Nanotoxicology 2016, 10, 1096–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Shi, W.; Rong, J.; Zha, S.; Guan, X.; Sun, H.; Liu, G. Exposure to waterborne nTiO2 reduces fertilization success and increases polyspermy in a bivalve mollusc: A threat to population recruitment. Environ. Sci. Technol. 2019, 53, 12754–12763. [Google Scholar] [CrossRef] [PubMed]
- Devakumar, C.; Gopalakrishnan, H.; Chinnasamy, A.; Subramanian, B.; Durai, P. Toxicity of silver nanoparticles on fertilization success and early development of the marine polychaete Hydroides elegans (Haswell, 1883). J. Basic Appl. Zool. 2017, 78, 1. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Manfra, L.; Boni, R.; Rotini, A.; Migliore, L.; Tosti, E. Cytotoxicity and genotoxicity of CuO nanoparticles in sea urchin spermatozoa through oxidative stress. Environ. Int. 2018, 118, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Hanna, S.K.; Miller, R.J.; Zhou, D.; Keller, A.A.; Lenihan, H.S. Accumulation and toxicity of metal oxide nanoparticles in a soft-sediment estuarine amphipod. Aquat. Toxicol. 2013, 142–143, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Gambardella, C.; Morgana, S.; Bari, G.D.; Ramoino, P.; Bramini, M.; Diaspro, A.; Falugi, C.; Faimali, M. Multidisciplinary screening of toxicity induced by silica nanoparticles during sea urchin development. Chemosphere 2015, 139, 486–495. [Google Scholar] [CrossRef]
- Mwaanga, P.; Carraway, E.R.; van den Hurk, P. The induction of biochemical changes in Daphnia magna by CuO and ZnO nanoparticles. Aquat. Toxicol. 2014, 150, 201–209. [Google Scholar] [CrossRef]
- Rim, K.T.; Song, S.W.; Kim, H.Y. Oxidative DNA damage from nanoparticle exposure and its application to workers’ health: A literature review. Saf. Health Work 2013, 4, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Huerta-García, E.; Márquez-Ramírez, S.G.; Ramos-Godinez, M.P.; López-Saavedra, A.; Herrera, L.A.; Parra, A.; Alfaro-Moreno, E.; Gómez, E.O.; López-Marure, R. Internalization of titanium dioxide nanoparticles by glial cells is given at short times and is mainly mediated by actin reorganization-dependent endocytosis. NeuroToxicology 2015, 51, 27–37. [Google Scholar] [CrossRef]
- Kazama, M.; Hino, A. Sea urchin spermatozoa generate at least two reactive oxygen species; the type of reactive oxygen species changes under different conditions. Mol. Reprod. Dev. 2012, 79, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Kukla, S.; Slobodskova, V.; Mazur, A.; Chelomin, V.; Kamenev, Y. Genotoxic testing of titanium dioxide nanoparticles in Far Eastern mussels, Mytilus trossulus. Pollution 2021, 7, 129–140. [Google Scholar] [CrossRef]
- Akcha, F.; Spagnol, C.; Rouxel, J. Genotoxicity of diuron and glyphosate in oyster spermatozoa and embryos. Aquat. Toxicol. 2012, 106–107, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaux, A.; Fiat, L.; Gillet, C.; Bony, S. Reproduction impairment following paternal genotoxin exposure in brown trout (Salmo trutta) and Arctic charr (Salvelinus alpinus). Aquat. Toxicol. 2011, 101, 405–411. [Google Scholar] [CrossRef]
- Santos, R.; Palos-Ladeiro, M.; Besnard, A.; Porcher, J.M.; Bony, S.; Sanchez, W.; Devaux, A. Relationship between DNA damage in sperm after ex vivo exposure and abnormal embryo development in the progeny of the three-spined stickleback. Reprod. Toxicol. 2013, 36, 6–11. [Google Scholar] [CrossRef]
- Pérez-Cerezales, S.; Martínez-Páramo, S.; Beirão, J.; Herráez, M.P. Evaluation of DNA damage as a quality marker for rainbow trout sperm cryopreservation and use of LDL as cryoprotectant. Theriogenology 2010, 74, 282–289. [Google Scholar] [CrossRef]
Figure 1.
Assessment of S.mirabilis sperm DNA damage from control and experimental groups. (A) Zn2+ exposure. (B) ZnO NPs exposure. (Mean ± standard deviation, N = 8, n = 400.) * Difference from the control is significant (p < 0.05).
Figure 1.
Assessment of S.mirabilis sperm DNA damage from control and experimental groups. (A) Zn2+ exposure. (B) ZnO NPs exposure. (Mean ± standard deviation, N = 8, n = 400.) * Difference from the control is significant (p < 0.05).


{kind=link}
Table 1.
The basic properties of ZnO NPs.
| Purity, % | Zeta Potential, mV | Particle Size, nm | Hydraulic Radius, nm | BET, m2/g |
|---|---|---|---|---|
| ≥99.5 | −39.4 | 40–50 | 200 | 58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kukla, S.P.; Chelomin, V.P.; Mazur, A.A.; Slobodskova, V.V. Zinc Oxide Nanoparticles Induce DNA Damage in Sand Dollar Scaphechinus mirabilis Sperm. Toxics 2022, 10, 348. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10070348
AMA Style
Kukla SP, Chelomin VP, Mazur AA, Slobodskova VV. Zinc Oxide Nanoparticles Induce DNA Damage in Sand Dollar Scaphechinus mirabilis Sperm. Toxics. 2022; 10(7):348. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10070348
Chicago/Turabian StyleKukla, Sergey Petrovich, Victor Pavlovich Chelomin, Andrey Alexandrovich Mazur, and Valentina Vladimirovna Slobodskova. 2022. "Zinc Oxide Nanoparticles Induce DNA Damage in Sand Dollar Scaphechinus mirabilis Sperm" Toxics 10, no. 7: 348. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10070348
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.