
Arsenic Soil Contamination and Its Effects on 5-Methylcytosine Levels in Onions and Arsenic Distribution and Speciation
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Analysis of Total Arsenic Content
2.3. Arsenic Speciation Analysis
2.4. Determination of Gas Exchange Parameters
2.5. Determination of Pigments
2.6. Determination of Fluorescence
2.7. 5-methylcytosine Levels Determination
2.8. Microscopy
2.9. Statistical Analyses
3. Results
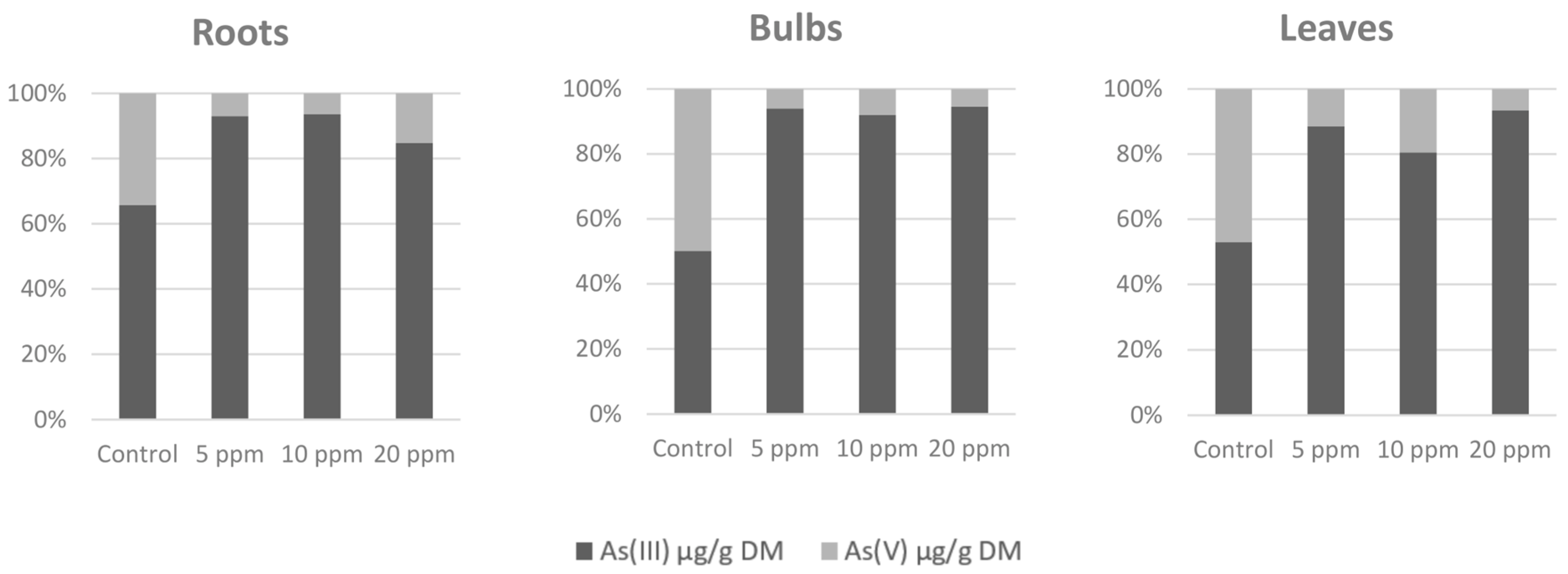
3.1. Arsenic Accumulation and Speciation in Different Parts of Plants
3.2. Physiological Responses to Arsenic Contamination
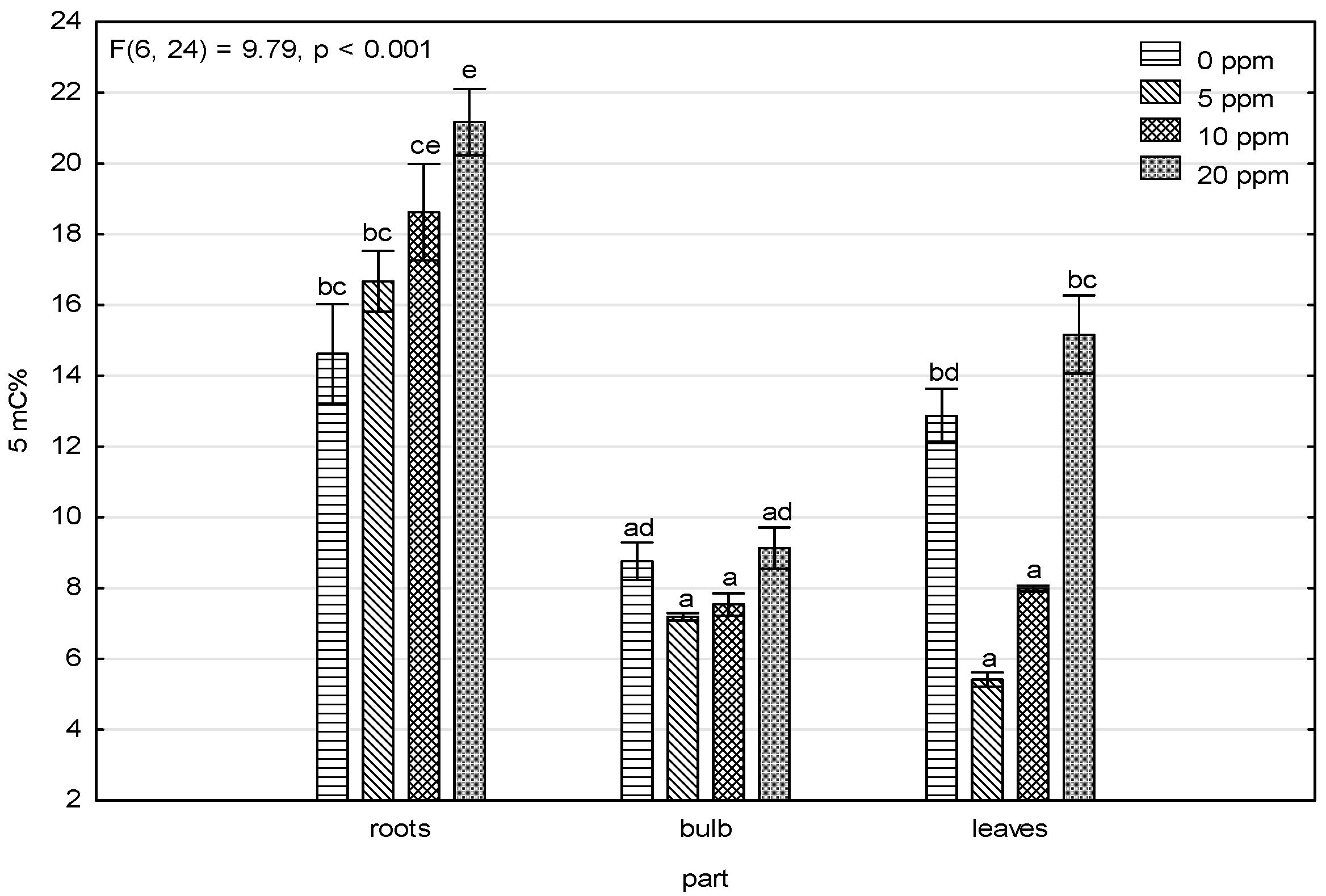
3.3. Changes in 5-Methylcytosine Level under Arsenic Exposure
3.4. Anatomic Changes in Tissues of Plants Exposed to Arsenic
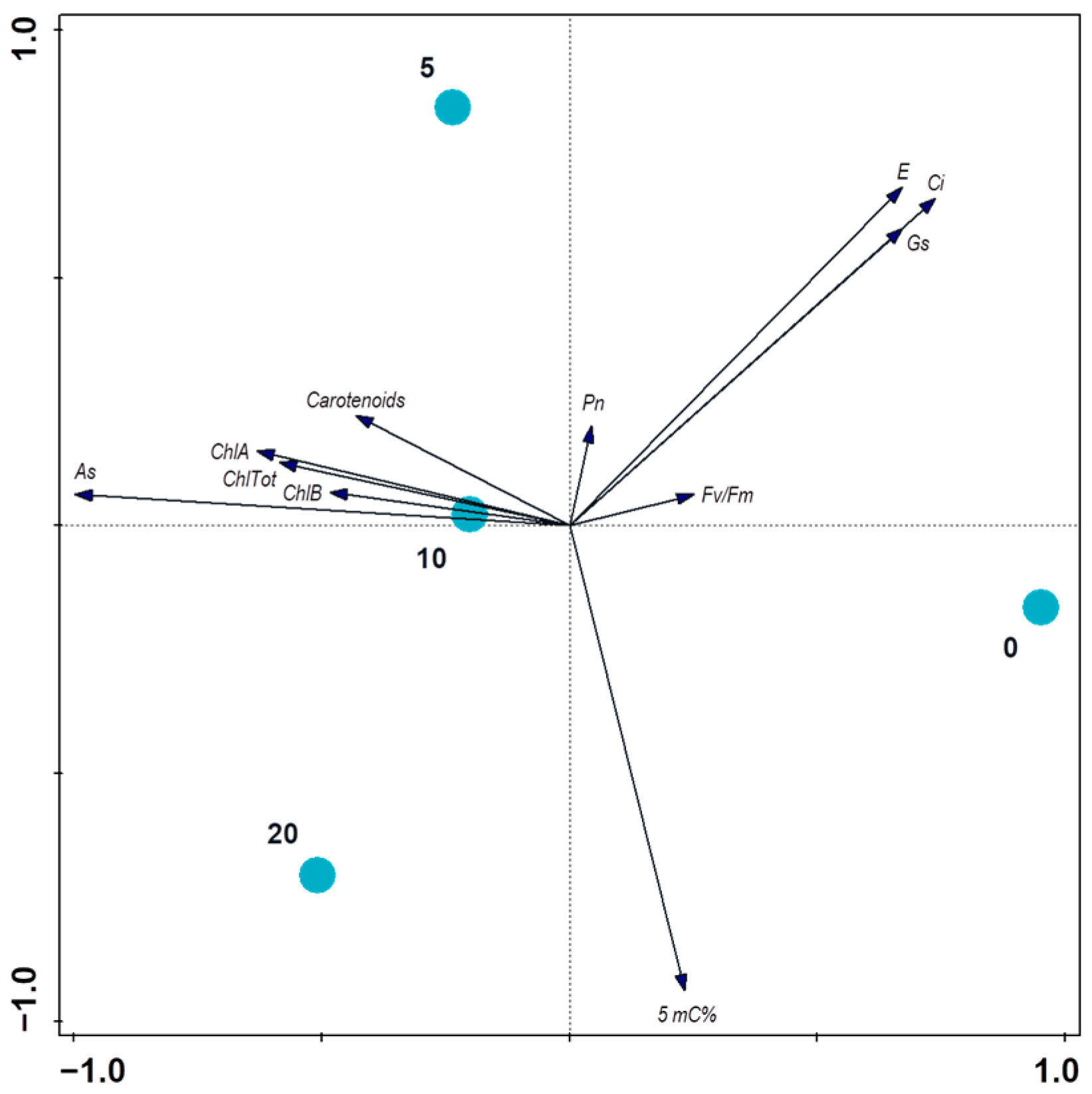
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, C.-Y.; Chen, T.-B. Arsenic accumulation by two brake ferns growing on an arsenic mine and their potential in phytoremediation. Chemosphere 2006, 63, 1048–1053. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.A.; Islam, F.; Ali, B.; Najeeb, U.; Mao, B.; Gill, R.A.; Siddique, K.H.M.; Zhou, W. Arsenic toxicity in plants: Cellular and molecular mechanisms of its transport and metabolism. Environ. Exp. Bot. 2016, 132, 42–52. [Google Scholar] [CrossRef]
- Muzaffar, S.; Khan, J.; Srivastava, R.; Gorbatyuk, M.S.; Athar, M. Mechanistic understanding of the toxic effects of arsenic and warfare arsenicals on human health and environment. Cell Biol. Toxicol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Ma, L.Q.; Santos, J.A.G. Three new arsenic hyperaccumulating ferns. Sci. Total Environ. 2006, 364, 24–31. [Google Scholar] [CrossRef]
- Samadder, S.R. Impact of Arsenic Pollution in Drinking Water on Life Expectancy: A GIS Study. KSCE J. Civ. Eng. 2010, 14, 681–691. [Google Scholar] [CrossRef]
- Samadder, S.R. Impact of Arsenic Pollution on Spatial Distribution of Human Development Index. KSCE J. Civ. Eng. 2010, 15, 975–982. [Google Scholar] [CrossRef]
- Samadder, S.R.; Nagesh Kumar, D.; Holden, N.M. An empirical model to predict arsenic pollution affected life expectancy. Popul. Environ. 2014, 36, 219–233. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, J.-W.; Han, Y.-H.; Rathinasabapathi, B.; Ma, L.Q. High As exposure induced substantial arsenite efflux in As-hyperaccumulator Pteris vittata. Chemosphere 2016, 144, 2189–2194. [Google Scholar] [CrossRef]
- Li, N.; Wang, J.; Song, W.-Y. Arsenic Uptake and Translocation in Plants. Plant Cell Physiol. 2016, 57, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Verma, P.K.; Pandeb, V.; Tripathic, R.D.; Chakrabartya, D. Transgenic Arabidopsis thaliana expressing fungal arsenic methyltransferase gene (WaarsM) showed enhanced arsenic tolerance via volatilization. Environ. Exp. Bot. 2016, 132, 113–120. [Google Scholar] [CrossRef]
- Oremland, R.S.; Stolz, J.F. The Ecology of Arsenic. Science 2003, 300, 939–944. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.J.; McGrath, S.P.; Meharg, A.A. Arsenic as a Food Chain Contaminant: Mechanisms of Plant Uptake and Metabolism and Mitigation Strategies. Annu. Rev. Plant Biol. 2010, 61, 535–559. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.; Lei, M.; Chen, T. Interaction of As and Sb in the hyperaccumulator Pteris vittata L.: Changes in As and Sb speciation by XANES. Environ. Sci. Pollut. Res. 2016, 23, 19137–19181. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses a current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Huang, Z.-C.; Chen, T.-B.; Lei, M.; Liu, Y.-R.; Hu, T.-D. Difference of Toxicity and Accumulation of Methylated and Inorganic Arsenic in Arsenic-Hyperaccumulating and -Hypertolerant Plants. Environ. Sci. Technol. 2008, 42, 5106–5111. [Google Scholar] [CrossRef]
- Campos, N.V.; Araújoa, T.O.; Arcanjo-Silva, S.; Freitas-Silva, L.; Azevedo, A.A.; Nunes-Nesia, A. Arsenic hyperaccumulation induces metabolic reprogramming in Pityrogramma calomelanos to reduce oxidative stress. Physiol. Plant. 2016, 157, 135–146. [Google Scholar] [CrossRef]
- Raab, A.; Feldmann, J.; Meharg, A.A. The Nature of Arsenic-Phytochelatin Complexes in Holcus lanatus and Pteris cretica. Plant Physiol. 2004, 134, 1113–1122. [Google Scholar] [CrossRef] [Green Version]
- Raab, A.; Ferreira, K.; Meharg, A.A.; Feldmann, J. Can arsenic-phytochelatin complex formation be used as an indicator for toxicity in Helianthus annuus? J. Exp. Bot. 2007, 58, 1333–1338. [Google Scholar] [CrossRef] [Green Version]
- Shoji, R.; Yajima, R.; Yano, Y. Arsenic speciation for the phytoremediation by the Chinese brake fern, Pteris vittata. J. Environ. Sci. 2008, 20, 1463–1468. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Dunham, S.J.; McGrath, S.P. Arsenic hyperaccumulation by different fern species. New Phytol. 2002, 156, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Souri, Z.; Karimi, N.; Sandalio, L.M. Arsenic Hyperaccumulation Strategies: An Overview. Front. Cell Dev. Biol. 2017, 5, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luongo, T.; Ma, L.Q. Characteristics of arsenic accumulation by Pteris and non-Pteris ferns. Plant Soil. 2004, 277, 117–126. [Google Scholar] [CrossRef]
- Chen, Y.; Hua, C.-Y.; Jia, M.-R.; Fu, J.-W.; Liu, X.; Han, Y.-H.; Liu, Y.; Rathinasabapathi, B.; Cao, Y.; Ma, L.Q. Heterologous Expression of Pteris vittata Arsenite Antiporter PvACR3;1 Reduces Arsenic Accumulation in Plant Shoots. Environ. Sci. Technol. 2017, 51, 10387–10395. [Google Scholar] [CrossRef] [PubMed]
- Maciaszczyk-Dziubinska, E.; Wawrzycka, D.; Wysocki, R. Arsenic and Antimony Transporters in Eukaryotes. Int. J. Mol. Sci. 2012, 13, 3527–3548. [Google Scholar] [CrossRef] [Green Version]
- Lukens, L.N.; Zhan, S. The plant genome’s methylation status and response to stress: Implications for plant improvement. Curr. Opin. Plant Biol. 2007, 10, 317–322. [Google Scholar] [CrossRef]
- Chen, M.; Lv, S.; Meng, Y. Epigenetic performers in plants. Dev. Growth Differ. 2010, 52, 555–566. [Google Scholar] [CrossRef]
- Grativol, C.; Hemerly, A.S.; Ferreira, P.C.G. Genetic and epigenetic regulation of stress responses in natural plant populations. Biochim. Biophys. Acta 2012, 1819, 176–185. [Google Scholar] [CrossRef]
- Kohli, R.; Zhang, Y. TET enzymes, TDG and the dynamics of DNA demethylation. Nature 2013, 502, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Wu, H.; Diep, D.; Yamaguchi, S.; D'Alessio, A.C.; Fung, H.L.; Zhang, K.; Zhang, Y. Genome-wide Analysis Reveals TET- and TDG-Dependent 5-Methylcytosine Oxidation Dynamics. Cell 2013, 153, 692–706. [Google Scholar] [CrossRef] [Green Version]
- Espinas, N.A.; Saze, H.; Saijo, Y. Epigenetic Control of Defense Signaling and Priming in Plants. Front. Plant Sci. 2016, 7, 1201. [Google Scholar] [CrossRef] [Green Version]
- Thiebaut, F.; Hemerly, A.S.; Ferreira, P.C.G. A Role for Epigenetic Regulation in the Adaptation and Stress Responses of Non-model Plants. Front. Plant Sci. 2019, 10, 246. [Google Scholar] [CrossRef] [Green Version]
- Meiqiong, T.; Jiao, Y.; Zhen, H.; Yali, H.; Zengqiang, L.; Dengjie, L.; Shan, C.; Hui, Z.; Jiao, P.; Xia, W.; et al. Physiological and DNA methylation analysis provides epigenetic insights into chromium tolerance in kenaf. J. Exp. Bot. 2022, 194, 104684. [Google Scholar] [CrossRef]
- Button, M.; Moriarty, M.M.; Watts, M.J.; Zhang, J.; Koch, I.; Reimer, K.J. Arsenic speciation in field-collected and laboratory-exposed earthworms Lumbricus terrestris. Chemosphere 2011, 85, 1277–1283. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Smilauer, P.; Leps, J. Multivariate Analysis of Ecological Data Using Canoco 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Dahal, B.M.; Fuerhacker, M.; Mentler, A.; Karki, K.B.; Shrestha, R.R.; Blum, W.E.H. Arsenic contamination of soils and agricultural plants through irrigation water in Nepal. Environ. Pollut. 2008, 155, 157–163. [Google Scholar] [CrossRef]
- Saha, N.; Zaman, M.R. Evaluation of possible health risks of heavy metals by consumption of foodstuffs available in the central market of Rajshahi City, Bangladesh. Environ. Monit. Assess 2013, 185, 3867–3878. [Google Scholar] [CrossRef]
- Rotting, T.S.; Mercado, M.; Garcia, M.E.; Quintanilla, J. Environmental distribution and health impacts of As and Pb in crops and soils near Vinto smelter, Oruro, Bolivia. Int. J. Environ. Sci. Technol. 2014, 11, 935–948. [Google Scholar] [CrossRef] [Green Version]
- Pinter, I.F.; Salomon, M.V.; Gil, R.; Mastrantonio, L.; Bottini, R.; Piccoli, P. Arsenic and trace elements in soil, water, grapevine and onion in Jachal, Argentina. Sci. Total Environ. 2018, 615, 1485–1498. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Sanchez, M.J.; Perez-Sirvent, C.; Martinez-Lopez, S.; Garcia-Lorenzo, M.L.; Agudo, I.; Martinez-Martinez, L.B.; Hernandez-Perez, C.; Bech, J. Uptake of potentially toxic elements by edible plants in experimental mining Technosols: Preliminary assessment. Environ. Geochem. Health 2022, 44, 1649–1665. [Google Scholar] [CrossRef] [PubMed]
- Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Gusman, G.S.; Oliveira, J.A.; Farnese, F.S.; Cambraia, J. Mineral nutrition and enzymatic adaptation induced by arsenate and arsenite exposure in lettuce plants. Plant Physiol. Biochem. 2013, 71, 307–314. [Google Scholar] [CrossRef]
- Anwar, S.; Khalilzadeh, R.; Khan, S.; Nisa, Z.; Bashir, R.; Pirzad, A.; Malik, A. Mitigation of drought stress and yield improvement in wheat by zinc foliar spray relates to enhanced water use efficiency and zinc contents. Int. J. Plant. Prod. 2021, 15, 377–389. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Ashraf, U.; Khan, I.; Wang, L. Alteration in Growth, Leaf Gas Exchange, and Photosynthetic Pigments of Maize Plants Under Combined Cadmium and Arsenic Stress. Water Air Soil Pollut. 2017, 228, 13. [Google Scholar] [CrossRef]
- Bolukbasi, E. Methylation Modelling and Epigenetic Analysis of Sunflower (Helianthus annuus L.) Seedlings Exposed to Cadmium Heavy Metal Stress. KSU J. Agric. Nat. 2022, 25, 467–475. [Google Scholar] [CrossRef]
- Aina, R.; Sgorbati, S.; Santagostino, A.; Labra, M.; Ghiani, A.; Citterio, S. Specific hypomethylation of DNA is induced by heavy metals in white clover and industrial hemp. Physiol. Plant. 2004, 121, 472–480. [Google Scholar] [CrossRef]
- Di, X.; Zheng, F.; Norton, G.J.; Beesley, L.; Zhang, Z.; Lin, H.; Zhi, S.; Liu, X.; Ding, Y. Physiological responses and transcriptome analyses of upland rice following exposure to arsenite and arsenate. Environ. Exp. Bot. 2021, 183, 104366. [Google Scholar] [CrossRef]
- Peralta, J.M.; Travaglia, C.; Romero-Puertas, M.C.; Molina-Moya, E.; Furlan, A.; Castro, S.; Bianucci, E. Decoding the antioxidant mechanisms underlying arsenic stress in roots of inoculated peanut plants. Plant Growth Regul. 2022, 97, 77–90. [Google Scholar] [CrossRef]
- Shi, S.; Wang, T.; Chen, Z.; Tang, Z.; Wu, Z.; Salt, D.E.; Chao, D.-Y.; Zhao, F.-J. OsHAC1;1 and OsHAC1;2 Function as Arsenate Reductases and Regulate Arsenic Accumulation. Plant Physiol. 2016, 172, 1708–1719. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Wu, L.; Jiang, L. Genotoxicity of arsenic evaluated by Allium-root micronucleus assay. Sci. Total Environ. 2007, 383, 232–236. [Google Scholar] [CrossRef]
- Wu, L.; Yi, H.; Min, Y. Assessment of arsenic toxicity using Allium/Vicia root tip micronucleus assays. J. Hazard. Mater. 2010, 176, 952–956. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ppm As(V) | Roots | Bulbs | Leaves |
|---|---|---|---|
| Control | 2.67 ± 0.11a | 0.60 ± 0.01b | 0.43 ± 0.03a |
| 5 | 1488.29 ± 23.31b | 10.39 ± 0.22c | 28.76 ± 2.06b |
| 10 | 1249.82 ± 41.75c | 26.94 ± 0.59a | 23.44 ± 0.13c |
| 20 | 1761.11 ± 101.84d | 28.06 ± 1.29a | 44.74 ± 1.27d |
| P | <0.001 | <0.001 | <0.001 |
| Parameter | Control | 5 ppm As(V) | 10 ppm As(V) | 20 ppm As(V) | p |
|---|---|---|---|---|---|
| Fv/Fm | 0.77 ± 0.00 | 0.75 ± 0.02 | 0.78 ± 0.02 | 0.75 ± 0.02 | 0.446 |
| Ci | 536.67 ± 50.63 b | 444.33 ± 26.52 b | 279.67 ± 17.57 a | 154.67 ± 10.27 a | <0.001 |
| E | 1.77 ± 0.11 b | 1.73 ± 0.05 b | 0.75 ± 0.03 a | 0.50 ± 0.02 a | <0.001 |
| gs | 0.07 ± 0.00 *b | 0.06 ± 0.00 *b | 0.02 ± 0.00 *a | 0.02 ± 0.00 *a | <0.001 |
| Pn | 6.77 ± 1.88 | 7.31 ± 0.69 | 5.54 ± 0.54 | 6.03 ± 0.60 | 0.677 |
| WUE | 3.99 ± 1.39 a | 4.25 ± 0.52 a | 7.37 ± 0.56 ab | 12.24 ± 1.58 b | 0.003 |
| Chl a | 0.83 ± 0.02 b | 1.45 ± 0.04 d | 0.80 ± 0.02 a | 1.42 ± 0.05 c | <0.001 |
| Chl b | 0.29 ± 0.01 b | 0.49 ± 0.02 c | 0.22 ± 0.01 a | 0.50 ± 0.02 d | <0.001 |
| Chlt | 1.12 ± 0.03 b | 1.94 ± 0.05 d | 1.02 ± 0.02 a | 1.92 ± 0.06 c | <0.001 |
| Car | 0.17 ± 0.01 b | 0.26 ± 0.02 d | 0.14 ± 0.01 a | 0.24 ± 0.01 c | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popov, M.; Kudrna, J.; Lhotská, M.; Hnilička, F.; Tunklová, B.; Zemanová, V.; Kubeš, J.; Vachová, P.; Česká, J.; Praus, L.; et al. Arsenic Soil Contamination and Its Effects on 5-Methylcytosine Levels in Onions and Arsenic Distribution and Speciation. Toxics 2023, 11, 237. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics11030237
Popov M, Kudrna J, Lhotská M, Hnilička F, Tunklová B, Zemanová V, Kubeš J, Vachová P, Česká J, Praus L, et al. Arsenic Soil Contamination and Its Effects on 5-Methylcytosine Levels in Onions and Arsenic Distribution and Speciation. Toxics. 2023; 11(3):237. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics11030237
Chicago/Turabian StylePopov, Marek, Jiří Kudrna, Marie Lhotská, František Hnilička, Barbora Tunklová, Veronika Zemanová, Jan Kubeš, Pavla Vachová, Jana Česká, Lukáš Praus, and et al. 2023. "Arsenic Soil Contamination and Its Effects on 5-Methylcytosine Levels in Onions and Arsenic Distribution and Speciation" Toxics 11, no. 3: 237. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics11030237