Effects of MP Polyethylene Microparticles on Microbiome and Inflammatory Response of Larval Zebrafish


Abstract
:
1. Introduction
2. Methods
2.1. Caretaking
2.2. Respirometry
2.3. RNA Extraction, Reverse Transcription (RT), and qPCR
2.4. qPCR
2.5. Respirometry and Gene Expression Statistical Analysis
2.6. Microbiome Analysis
2.6.1. Sampling, DNA Extraction, Sequencing, and Pre-Processing
2.6.2. CLC Analysis
3. Results
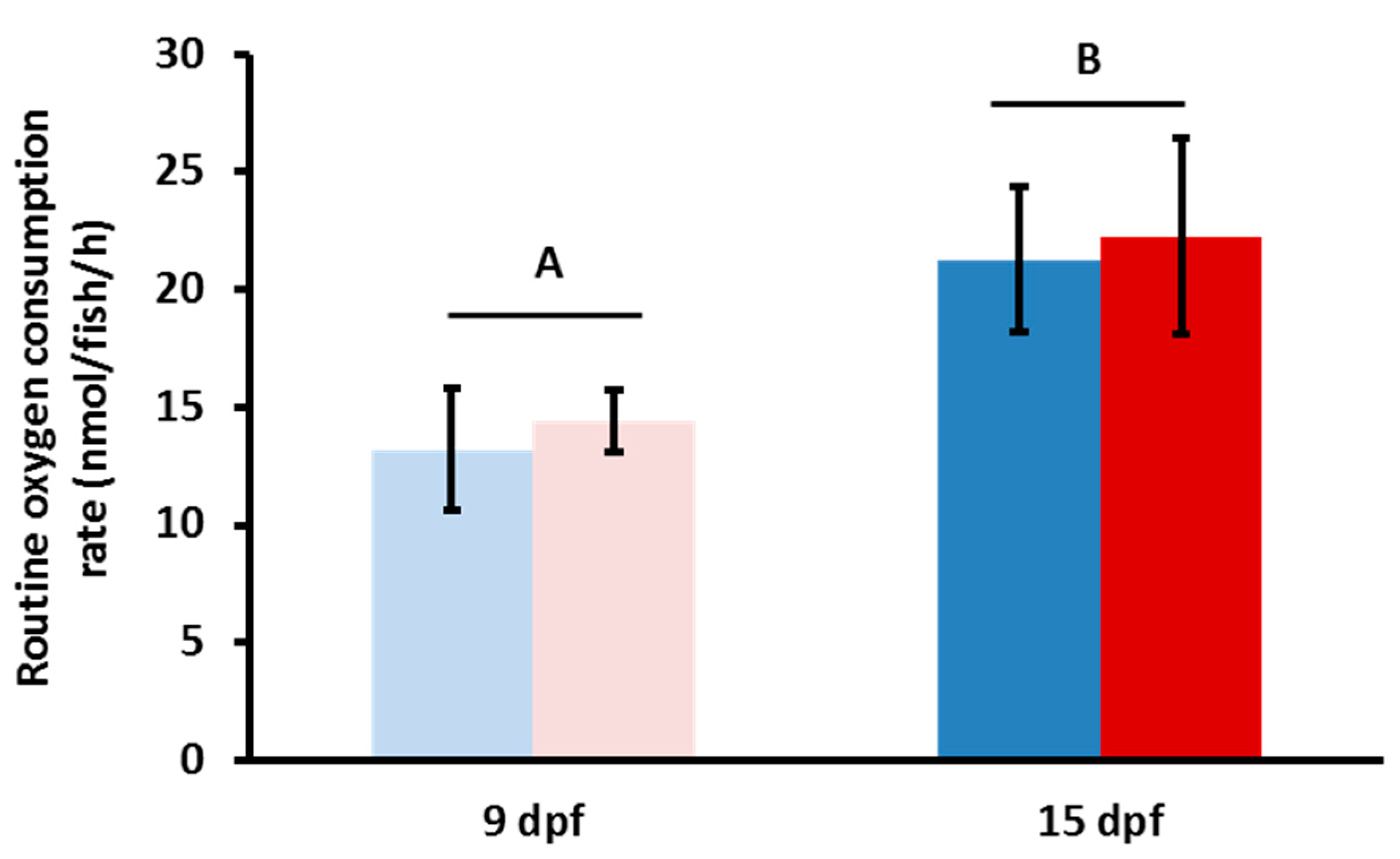
3.1. Respirometry
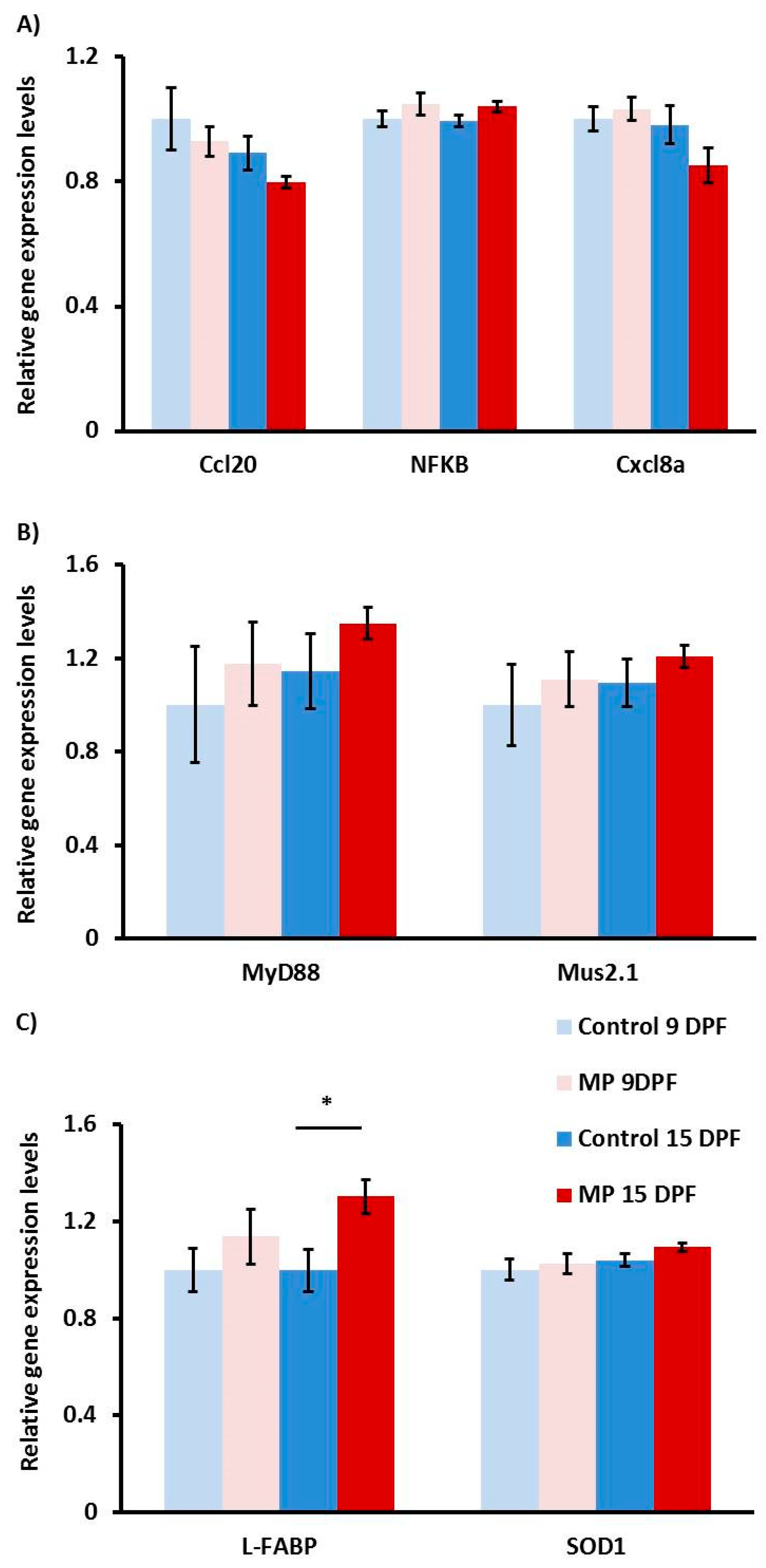
3.2. Gene Expression Analysis
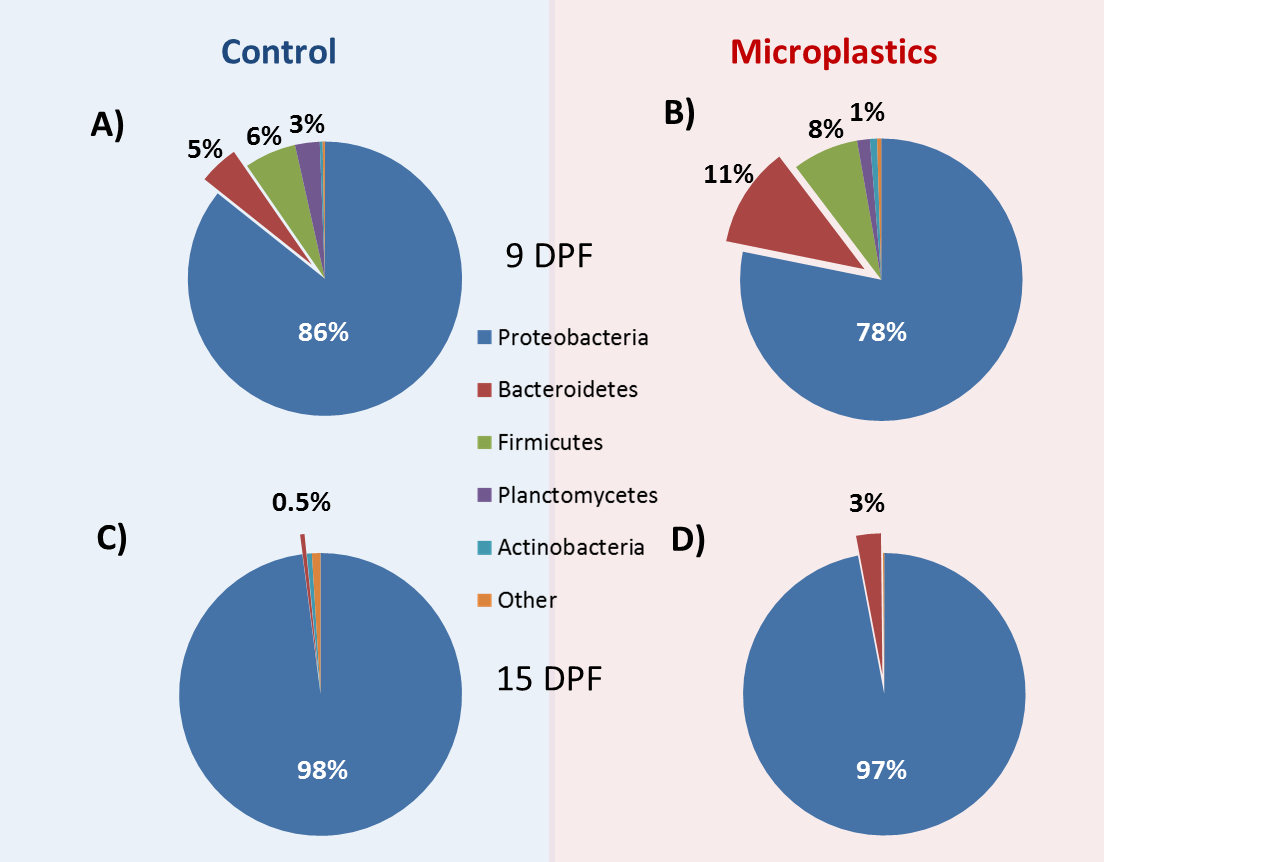
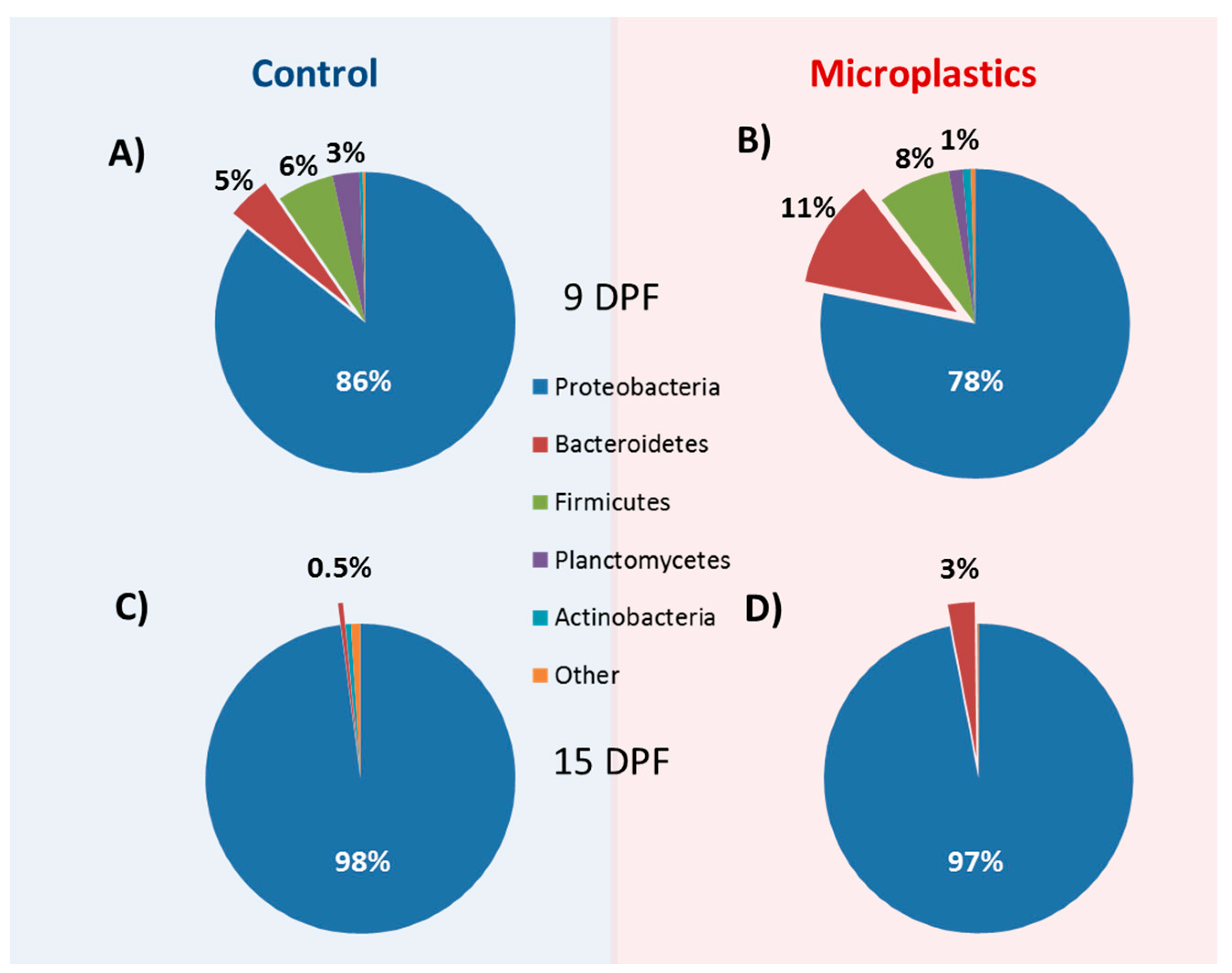
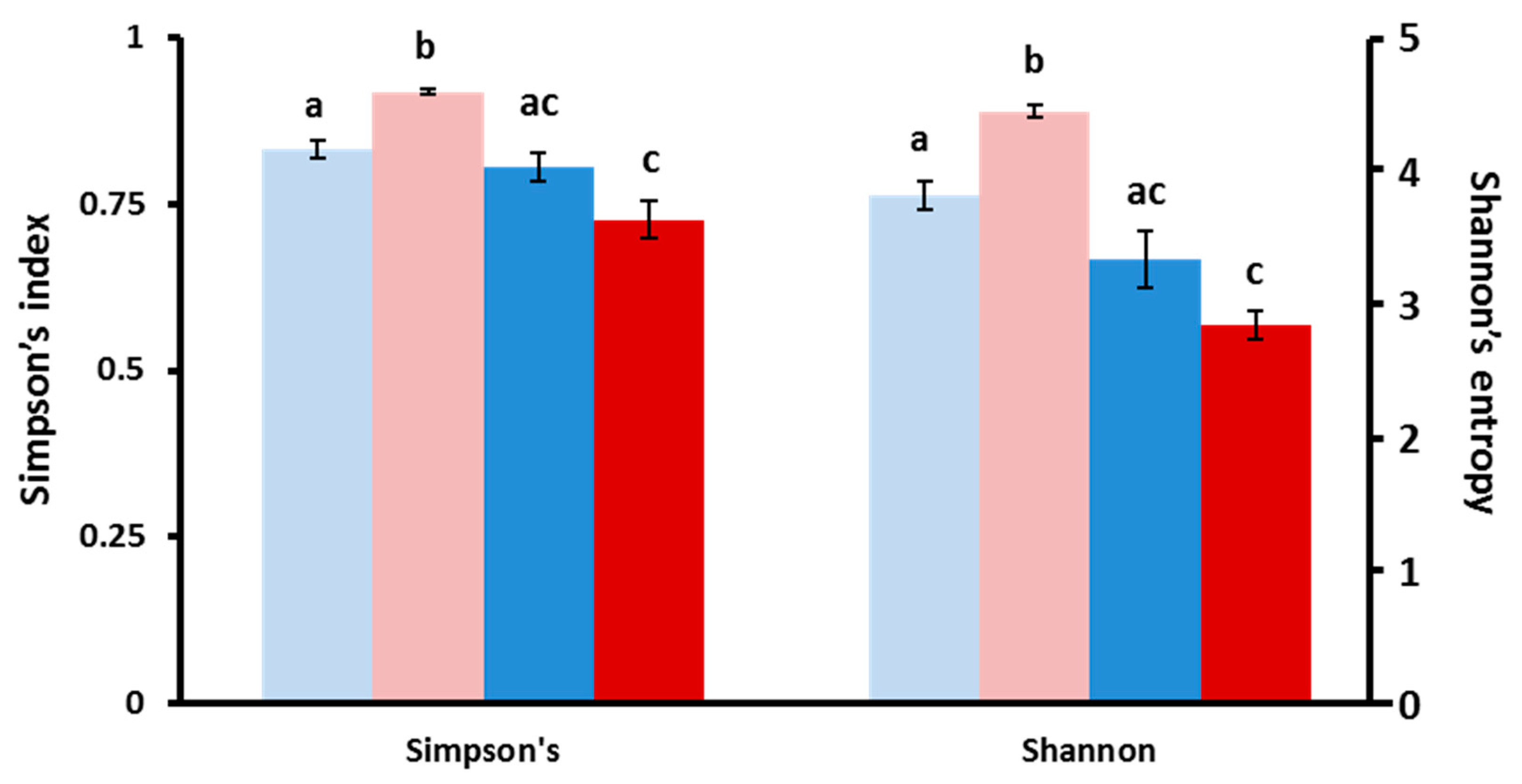
3.3. Structure of Gut Bacteria Communities
3.4. Core Microbial Composition
3.5. Microbial Community Changes through Ontogeny
3.6. MP Exposure Disturbs the Microbiome
4. Discussion
Microbiome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, L.K. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, D.K.A.; Galgani, F.; Thompson, C.R.; Barlaz, M. Accumulation and Fragmentation of Plastic Debris in Global Environments. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009, 364, 1985–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, P.K.; Hakkarainen, M.; Varma, I.K.; Albertsson, A.C. Degradable Polyethylene: Fantasy or Reality. Environ. Sci. Technol. 2011, 45, 4217–4227. [Google Scholar] [CrossRef] [PubMed]
- Andrady, A.L. Microplastics in the Marine Environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- Cheshire, A.; Ellik, A.; Barbière, J.; Cohen, Y.; Evans, S.; Jarayabhand, S.; Jeftic, L.; Jung, R.T.; Kinsey, S.; Kusui, E.T.; et al. UNEP/IOC Guidelines on Survey and Monitoring of Marine Litter; Regional Seas Reports and Studies No. 186 IOC Technical Series No. 83. UNEP Regional Seas Reports and Studies; Intergovernmental Oceanographic Commission: Paris, France, 2009. [Google Scholar]
- Frias, J.P.; Nash, R. Microplastics: Finding a consensus on the definition. Mar. Pollut. Bull. 2019, 138, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, R.V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef]
- Obbard, R.W.; Sadri, S.; Wong, Q.Y.; Khitun, A.A.; Baker, I.; Thompson, R.C. Global Warming Releases Microplastic Legacy Frozen in Arctic Sea Ice. Earths Future 2014, 2, 315–320. [Google Scholar] [CrossRef]
- Sun, J.; Dai, X.; Wang, Q.; Loosdrecht, V.M.C.M.; Ni, B.J. Microplastics in Wastewater Treatment Plants: Detection, Occurrence and Removal. Water Res. 2019, 152, 21–37. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.; Chen, J.P. Microplastics in Freshwater Systems: A Review on Occurrence, Environmental Effects, and Methods for Microplastics Detection. Water Res. 2018, 137, 362–374. [Google Scholar] [CrossRef]
- Shahul, H.F.; Bhatti, M.S.; Anuar, N.; Anuar, N.; Mohan, P.; Periathamby, A. Worldwide Distribution and Abundance of Microplastic: How Dire Is the Situation? Waste Manag. Res. 2018, 36, 873–897. [Google Scholar] [CrossRef]
- Vermaire, C.J.; Pomeroy, C.; Herczegh, M.S.; Haggart, O.; Murphy, M. Microplastic Abundance and Distribution in the Open Water and Sediment of the Ottawa River, Canada, and Its Tributaries. FACETS. 2017, 2, 301–314. [Google Scholar] [CrossRef] [Green Version]
- LeMoine, C.M.R.; Kelleher, B.M.; Lagarde, R.; Northam, C.; Elebute, O.O.; Cassone, B.J. Transcriptional Effects of Polyethylene Microplastics Ingestion in Developing Zebrafish (Danio rerio). Environ. Pollut. 2018, 243 Pt A, 591–600. [Google Scholar] [CrossRef]
- Wright, S.L.; Thompson, R.C.; Galloway, T.S. The Physical Impacts of Microplastics on Marine Organisms: A Review. Environ. Pollut. 2013, 178, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.K.; Chen, Z.C.; Crawford, P.A.; Fu, X.; Burgess, S.C.; Lai, L.; Leone, T.C.; Kelly, D.P.; Finck, B.N. Liver-Specific PGC-1beta Deficiency Leads to Impaired Mitochondrial Function and Lipogenic Response to Fasting-Refeeding. PLoS ONE 2012, 7, e52645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef]
- Pitt, J.A.; Kozal, J.S.; Jayasundara, N.; Massarsky, A.; Trevisan, R.; Geitner, N.; Wiesner, M.; Levin, E.D.; Di Giulio, R.T. Uptake, Tissue Distribution, and Toxicity of Polystyrene Nanoparticles in Developing Zebrafish (Danio rerio). Aquat. Toxicol. 2018, 194, 185–194. [Google Scholar] [CrossRef]
- Pedà, C.; Leteria, C.; Fossi, C.M.; Gai, F.; Andaloro, F.; Genovese, L.; Perdichizzi, A.; Romeo, T.; Maricchiolo, G. Intestinal Alterations in European Sea Bass Dicentrarchus labrax (Linnaeus, 1758) Exposed to Microplastics: Preliminary Results. Environ. Pollut. 2016, 212, 251–256. [Google Scholar] [CrossRef]
- Jeong, C.B.; Won, E.J.; Kang, H.-M.; Lee, M.C.; Hwang, D.S.; Hwang, U.K.; Zhou, B.; Souissi, S.; Lee, S.J.; Lee, J.S. Microplastic Size-Dependent Toxicity, Oxidative Stress Induction, and p-JNK and p-P38 Activation in the Monogonont Rotifer (Brachionus koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS crosstalk in inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef]
- Duan, X.D.; Feng, L.; Jiang, W.D.; Wu, P.; Liu, Y.; Kuang, S.Y.; Tang, T.; Tang, W.N.; Zhang, Y.A.; Zhou, X.Q. Dietary soybean β-conglycinin suppresses growth performance and inconsistently triggers apoptosis in the intestine of juvenile grass carp (Ctenopharyngodon idella) in association with ROS-mediated MAPK signalling. Aquac. Nutr. 2019, 25, 770–782. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, C.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaser, A.; Tilg, H. “Metabolic Aspects” in Inflammatory Bowel Diseases. Curr. Drug Deliv. 2012, 9, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Weisshof, R.; Chermesh, I. Micronutrient Deficiencies in Inflammatory Bowel Disease. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Slyepchenko, A.; Maes, M.; Machado-Vieira, R.; Anderson, G.; Solmi, M.; Sanz, Y.; Berk, M.; Köhler, C.A.; Carvalho, A.F. Intestinal dysbiosis, gut hyperpermeability and bacterial translocation: Missing links between depression, obesity and type 2 diabetes. Curr. Pharm Des. 2016, 22, 6087–6106. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Kim, N.; Roh, J.H.; Chun, W.-K.; Ho, D.T.; Lee, Y.; Kim, D.-H. Administration of antibiotics can cause dysbiosis in fish gut. Aquaculture 2019, 512, 734330. [Google Scholar] [CrossRef]
- Qiao, R.; Cheng, S.; Lu, Y.; Zhang, Y.; Ren, H.; Lemos, B. Microplastics Induce Intestinal Inflammation, Oxidative Stress, and Disorders of Metabolome and Microbiome in Zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef]
- Wan, Z.; Wang, C.; Zhou, J.; Shen, M.; Wang, X.; Fu, Z.; Jin, Y. Effects of polystyrene microplastics on the composition of the microbiome and metabolism in larval zebrafish. Chemosphere 2019, 217, 646–658. [Google Scholar] [CrossRef]
- Fackelmann, G.; Sommer, S. Microplastics and the gut microbiome: How chronically exposed species may suffer from gut dysbiosis. Mar. Pollut. Bull. 2019, 143, 193–203. [Google Scholar] [CrossRef]
- Lu, L.; Luo, T.; Zhao, Y.; Cai, C.; Fu, Z.; Jin, Y. Interaction between microplastics and microorganism as well as gut microbiota: A consideration on environmental animal and human health. Sci. Total Environ. 2019, 667, 94–100. [Google Scholar] [CrossRef]
- Luo, T.; Wang, C.; Pan, Z.; Jin, C.; Fu, Z.; Jin, Y. Maternal polystyrene microplastic exposure during gestation and lactation altered metabolic homeostasis in the dams and their F1 and F2 offspring. Environ. Sci. Technol. 2019, 53, 10978–10992. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Chen, Q.-L.; An, X.-L.; Yang, X.-R.; Christie, P.; Ke, X.; Wu, L.H.; Zhu, Y.G. Exposure of soil collembolans to microplastics perturbs their gut microbiota and alters their isotopic composition. Soil Biol. Biochem. 2018, 116, 302–310. [Google Scholar] [CrossRef]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Nhieu, J.T.V.; Furet, J.P. Microbial Dysbiosis in Colorectal Cancer (CRC) Patients. PLoS ONE 2011, 6, e16393. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Sanderson, J.D.; Churcher, C.; Parkes, G.C.; Hudspith, B.N.; Rayment, N.; Brostoff, J.; Parkhill, J.; Gordon, D.; Petrovska, L. High-Throughput Clone Library Analysis of the Mucosa-Associated Microbiota Reveals Dysbiosis and Differences between Inflamed and Non-Inflamed Regions of the Intestine in Inflammatory Bowel Disease. BMC Microbiol. 2011, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Ma, C.; Han, L.; Nawaz, M.; Gao, F.; Zhang, X.; Yu, P.; Zhao, C.; Li, L.; Zhou, A.; et al. Molecular Characterisation of the Faecal Microbiota in Patients with Type II Diabetes. Curr. Microbiol. 2010, 61, 69–78. [Google Scholar] [CrossRef]
- Reese, A.T.; Cho, E.H.; Kiltzman, B.; Nichols, S.P.; Wisniewski, N.A.; Villa, M.M.; Durand, H.K.; Jiang, S.; Midani, F.S.; Nimmagadda, S.N.; et al. Antibiotic-Induced Changes in the Microbiota Disrupt Redox Dynamics in the Gut. eLife 2018, 7, e35987. [Google Scholar] [CrossRef]
- Gil, D.; Alfonso, I.S.; Pérez, R.L.; Muriel, J.; Monclús, R. Harsh Conditions during Early Development Influence Telomere Length in an Altricial Passerine: Links with Oxidative Stress and Corticosteroids. J. Evol. Biol. 2019, 32, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.M.; Cowan, M.; Moonah, S.N.; Petri, W.A. The Impact of Systemic Inflammation on Neurodevelopment. Trends Mol. Med. 2018, 24, 794–804. [Google Scholar] [CrossRef]
- Mead, K.S.; Denny, M.W. The Effects of Hydrodynamic Shear Stress on Fertilization and Early Development of the Purple Sea Urchin Strongylocentrotus purpuratus. Biol. Bull. 1995, 188, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Xia, J.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z. Polystyrene Microplastics Induce Microbiota Dysbiosis and Inflammation in the Gut of Adult Zebrafish. Environ. Pollut. 2018, 235, 322–329. [Google Scholar] [CrossRef]
- Song, Y.K.; Hong, H.S.; Jang, M.; Kang, J.-H.; Kwon, O.Y.; Han, G.M.; Shim, W.J. Large Accumulation of Micro-Sized Synthetic Polymer Particles in the Sea Surface Microlayer. Environ. Sci. Technol. 2014, 48, 9014–9021. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.J.R.; Lewis, C.; Goodhead, R.M.; Beckett, S.J.; Moger, J.; Tyler, C.R.; Galloway, T.S. Uptake and retention of microplastics by the shore crab Carcinus maenas. Environ. Sci. Technol. 2014, 48, 8823e8830. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Galloway, T.S. The Impact of Polystyrene Microplastics on Feeding, Function and Fecundity in the Marine Copepod Calanus Helgolandicus. Environ. Sci. Technol. 2015, 49, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.F.; Syberg, K.; Shashoua, Y. Influence of polyethylene microplastic beads on the uptake and localization of silver in zebrafish (Danio rerio). Environ. Pollut. 2015, 206, 73e79. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book; University of Oregon Press: Eugene, OR, USA, 1995. [Google Scholar]
- Harianto, J.; Carey, N.; Byrne, M. respR—An R package for the manipulation and analysis of respirometry data. Methods Ecol. Evol. 2019, 10, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Rombough, P.; Drader, H. Hemoglobin Enhances Oxygen Uptake in Larval Zebrafish (Danio rerio) but Only under Conditions of Extreme Hypoxia. J. Exp. Biol. 2009, 212 Pt 6, 778–784. [Google Scholar] [CrossRef] [Green Version]
- Northam, C.; LeMoine, C.M.R. Metabolic Regulation by the PGC-1α and PGC-1β Coactivators in Larval Zebrafish (Danio rerio). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 234, 60–67. [Google Scholar] [CrossRef]
- Untergasser, A.S.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- McCurley, A.T.; Callard, G.V. Characterization of housekeeping genes in zebrafish: Male-female differences and effects of tissue type, developmental stage and chemical treatment. BMC Mol. Biol. 2008, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Ibarra, M.S.; Etichetti, C.B.; Benedetto, C.D.; Rosano, G.L.; Margarit, E.; Sal, G.D.; Mione, M.; Girardini, J. Dynamic Regulation of Pin1 Expression and Function during Zebrafish Development. PLoS ONE 2017, 12, e0175939. [Google Scholar] [CrossRef]
- Vaart, V.D.M.; Soest, V.J.J.; Spaink, H.P.; Meijer, A.H. Functional analysis of a zebrafish myd88 mutant identifies key transcriptional components of the innate immune system. Dis. Model Mech. 2013, 6, 841–854. [Google Scholar] [CrossRef] [Green Version]
- Rotman, J.; Gils, V.W.; Butler, D.; Spaink, H.P.; Meijer, A.H. Rapid screening of innate immune gene expression in zebrafish using reverse transcription-multiplex ligation-dependent probe amplification. BMC Res. Notes 2011, 4, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Zhang, Y.; Han, L.; He, Q.; Hou, H.; Han, J.; Wang, X.; Li, C.; Cen, J.; Liu, K. Oxidative stress-mediated developmental toxicity induced by isoniazide in zebrafish embryos and larvae. J. Appl. Toxicol. 2017, 37, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Kim, C.Y.; Oh, H.; Kim, U.; Kim, J.; Jung, C.R.; Lee, B.H.; Lee, S.; Chang, S.N.; Lee, J.M.; et al. Development of an alternative zebrafish model for drug-induced intestinal toxicity. J. Appl. Toxicol. 2018, 38, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Brugman, S.; Witte, M.; Scholman, R.C.; Klein, M.R.; Boes, M.; Nieuwenhuis, E.E. T Lymphocyte–Dependent and–Independent Regulation of Cxcl8 Expression in Zebrafish Intestines. J. Immunol. 2014, 192, 484–491. [Google Scholar] [CrossRef] [Green Version]
- Livak, J.K.; Schmittgen, D.T. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Cassone, B.J.; Grove, H.C.; Elebute, O.; Villanueva, S.M.; LeMoine, C.M. Role of the intestinal microbiome in low-density polyethylene degradation by caterpillar larvae of the greater wax moth, Galleria mellonella. Proc. Biol. Sci. 2020, 287, 20200112. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Gallo, F.; Fossi, C.; Weber, R.; Santillo, D.; Sousa, J.; Ingram, I.; Nadal, A.; Romano, D. Marine Litter Plastics and Microplastics and Their Toxic Chemicals Components: The Need for Urgent Preventive Measures. Environ. Sci. Eur. 2018, 30. [Google Scholar] [CrossRef]
- Galloway, S.T.; Lewis, C.N. Marine Microplastics Spell Big Problems for Future Generations. Proc. Natl. Acad. Sci. USA 2016, 113, 2331–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Chatterjee, S. Microplastic Pollution, a Threat to Marine Ecosystem and Human Health: A Short Review. Environ. Sci. Pollut. Res. Int. 2017, 24, 21530–21547. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gundlach, M.; Shouye, Y.; Jiang, J.; Velki, M.; Yin, D.; Hollert, H. Quantitative Investigation of the Mechanisms of Microplastics and Nanoplastics toward Zebrafish Larvae Locomotor Activity. Sci. Total. Environ. 2017, 584–585, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Mazurais, D.; Ernande, B.; Quazuguel, P.; Severe, A.; Huelvan, C.; Madec, L.; Mouchel, O.; Soudant, P.; Robbens, J.; Huvet, A.; et al. Evaluation of the Impact of Polyethylene Microbeads Ingestion in European Sea Bass (Dicentrarchus labrax) Larvae. Mar. Environ. Res. 2015, 112 Pt A, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a Model Vertebrate for Investigating Chemical Toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, L.; Cheng, J. Exposure to Microplastics Decreases Swimming Competence in Larval Zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2019, 176, 226–233. [Google Scholar] [CrossRef]
- Reniere, M.L. Reduce, Induce, Thrive: Bacterial Redox Sensing during Pathogenesis. J. Bacteriol. 2018, 200, e00128-18. [Google Scholar] [CrossRef] [Green Version]
- Bonneaud, C.; Wilson, R.S.; Seebacher, F. Immune-Challenged Fish Up-Regulate Their Metabolic Scope to Support Locomotion. PLoS ONE 2016, 11, e0166028. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Yan, C.; Ying, Y.H.; Lu, W. Behavioral and Metabolic Phenotype Indicate Personality in Zebrafish (Danio rerio). Front. Physiol. 2018, 9, 653. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.S.; Cho, H.-J.; Kim, E.; Huh, Y.H.; Kim, H.-J.; Kim, B.; Kang, T.; Lee, J.-S.; Jeong, J. Bioaccumulation of Polystyrene Nanoplastics and Their Effect on the Toxicity of Au Ions in Zebrafish Embryos. Nanoscale 2019, 11, 3200–3207. [Google Scholar] [CrossRef]
- Novoa, B.; Figueras, A. Zebrafish: Model for the Study of Inflammation and the Innate Immune Response to Infectious Diseases. Adv. Exp. Med. Biol. 2012, 946, 253–275. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Hoshino, K. Myeloid Differentiation Factor 88–Dependent and –Independent Pathways in Toll-Like Receptor Signaling. J. Infect. Dis. 2003, 187 (Suppl. 2), S356–S363. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target Ther. 2017, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sar, A.M.; Stockhammer, O.W.; Laan, C.; Spaink, H.P.; Bitter, W.; Meijer, A.H. MyD88 Innate Immune Function in a Zebrafish Embryo Infection Model. Infect. Immun. 2006, 74, 2436–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugman, S. The Zebrafish as a Model to Study Intestinal Inflammation. Dev. Comp. Immunol. 2016, 64, 82–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanyang, L.; Liu, X.; Chen, X.; Yujia, Q.; Jiarong, F.; Jun, S.; Zhihua, R. Application of Zebrafish Models in Inflammatory Bowel Disease. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Bonkovsky, H.L.; de Lemos, A.; Burczynski, F.J. Recent Insights into the Biological Functions of Liver Fatty Acid Binding Protein 1. J. Lipid Res. 2015, 56, 2238–2247. [Google Scholar] [CrossRef] [Green Version]
- Maragakis, N.J.; Rothstein, J.D. Motor Neuron Disease: Amyotrophic Lateral Sclerosis. Mol. Neurobiol. 2007, 307–319. [Google Scholar] [CrossRef]
- Jeong, C.B.; Kang, H.M.; Lee, M.C.; Kim, D.H.; Han, J.; Hwang, S.; Souissi, S.; Lee, S.J.; Shin, K.H.; Park, H.G.; et al. Adverse Effects of Microplastics and Oxidative Stress-Induced MAPK/Nrf2 Pathway-Mediated Defense Mechanisms in the Marine Copepod Paracyclopina nana. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, P.R. The Gut Microbiota of Marine Fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef]
- Roeselers, G.; Mittge, E.K.; Stephens, Z.W.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a Core Gut Microbiota in the Zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, W.Z.; Burns, R.A.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J. The Composition of the Zebrafish Intestinal Microbial Community Varies across Development. ISME J. 2016, 10, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phelps, D.; Brinkman, N.E.; Keely, S.P.; Annken, E.M.; Catron, T.R.; Betancourt, D.; Wood, C.E.; Espenschied, S.T.; Rawls, J.F.; Tal, T. Microbial Colonization Is Required for Normal Neurobehavioral Development in Zebrafish. Sci. Rep. 2017, 7, 11244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.P.; Sapp, M.; Schratzberger, M.; Osborn, M.A. Interactions Between Microorganisms and Marine Microplastics: A Call for Research. Mar. Technol. Soc. J. 2011, 45, 12–20. [Google Scholar] [CrossRef]
- Lu, K.; Ruxia, Q.; Hao, A.; Zhang, Y. Influence of Microplastics on the Accumulation and Chronic Toxic Effects of Cadmium in Zebrafish (Danio rerio). Chemosphere 2018, 202, 514–520. [Google Scholar] [CrossRef]
- Prakash, S.; Rodes, L.; Coussa-Charley, M.; Tomaro-Duchnesneau, C. Gut Microbiota: Next Frontier in Understanding Human Health and Development of Biotherapeutics. Biologics 2011, 5, 71–86. [Google Scholar] [CrossRef] [Green Version]
- Vael, C.; Nelen, V.; Verhulst, L.S.; Goossens, H.; Desager, K.N. Early Intestinal Bacteroides Fragilis Colonisation and Development of Asthma. BMC Pulm. Med. 2008, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Cadet, J.; Wagner, R.J. DNA Base Damage by Reactive Oxygen Species, Oxidizing Agents, and UV Radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef]
- Dumitrescu, L.; Popescu-Olaru, I.; Cozma, L.; Tulbă, D.; Eugen, M.H.; Ceafalan, L.C.; Gherghiceanu, M.; Popescu, B.O. Oxidative Stress and the Microbiota-Gut-Brain Axis. Oxid. Med. Cell Longev. 2018, 406594. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.A.; Hug, L.A. Distribution of reactive oxygen species defense mechanisms across domain bacteria. Free Radic. Biol. Med. 2019, 140, 93–102. [Google Scholar] [CrossRef]
- Machado, M.V.; Cortez-Pinto, H. Gut Microbiota and Nonalcoholic Fatty Liver Disease. Ann. Hepatol. 2012, 11, 440–449. [Google Scholar] [CrossRef]
- Suzuki, A.; Ito, M.; Hamaguchi, T.; Mori, H.; Takeda, Y.; Baba, R.; Watanabe, T.; Kurokawa, K.; Asakawa, S.; Hirayama, M.; et al. Quantification of Hydrogen Production by Intestinal Bacteria That Are Specifically Dysregulated in Parkinson’s Disease. PLoS ONE 2018, 13, e0208313. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Accession Number |
| ef1α51 | CTTCTCAGGCTGACTGTGC | CCGCTAGCATTACCCTCC | AY422992 |
| Rpl-13a52 | TCTGGAGGACTGTAAGAGGTATGC | AGACGCACAATCTTGAGAGCAG | NM_212784.1 |
| MyD8853 | GAGGCGATTCCAGTAACAGC | GAAAGCATCAAAGGTCTCAGGTG | NM_212814.2 |
| Ccl2054 | ATCAATCTGCGCTAATCCATCAC | TGGTGAACATGCTCATCGTCTT | NM_001113595.1 |
| L-FABP55 | ACGTGGCAGGTTTACGCTCAG | TTGGAGGTGATGGTGAAGTCG | BC164928.1 |
| SOD156 | GTCGTCTGGCTTGTGGAGTG | TGTCAGCGGGCTAGTGCTT | AY324390.1 |
| NF-κβ56 | CCAAATCCCAAAAGGTTAGAGATTT | CCTCTTAGGGCTGAGCGAATT | XM_005156814.2 |
| Cxcl8a57 | TGTTTTCCTGGCATTTCTGAC | TTTACAGTGTGGGCTTGGAGGG | XM_009306855.3 |
| Mus2.150 | TGGTGGACCAGTGTGAAAAA | GGTCCAAAACCCAGCTACAA | XM_021470771.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurchaba, N.; Cassone, B.J.; Northam, C.; Ardelli, B.F.; LeMoine, C.M.R. Effects of MP Polyethylene Microparticles on Microbiome and Inflammatory Response of Larval Zebrafish. Toxics 2020, 8, 55. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030055
Kurchaba N, Cassone BJ, Northam C, Ardelli BF, LeMoine CMR. Effects of MP Polyethylene Microparticles on Microbiome and Inflammatory Response of Larval Zebrafish. Toxics. 2020; 8(3):55. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030055
Chicago/Turabian StyleKurchaba, Nicholas, Bryan J. Cassone, Caleb Northam, Bernadette F. Ardelli, and Christophe M. R. LeMoine. 2020. "Effects of MP Polyethylene Microparticles on Microbiome and Inflammatory Response of Larval Zebrafish" Toxics 8, no. 3: 55. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics8030055