Detection of Aflatoxigenic and Atoxigenic Mexican Aspergillus Strains by the Dichlorvos–Ammonia (DV–AM) Method


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
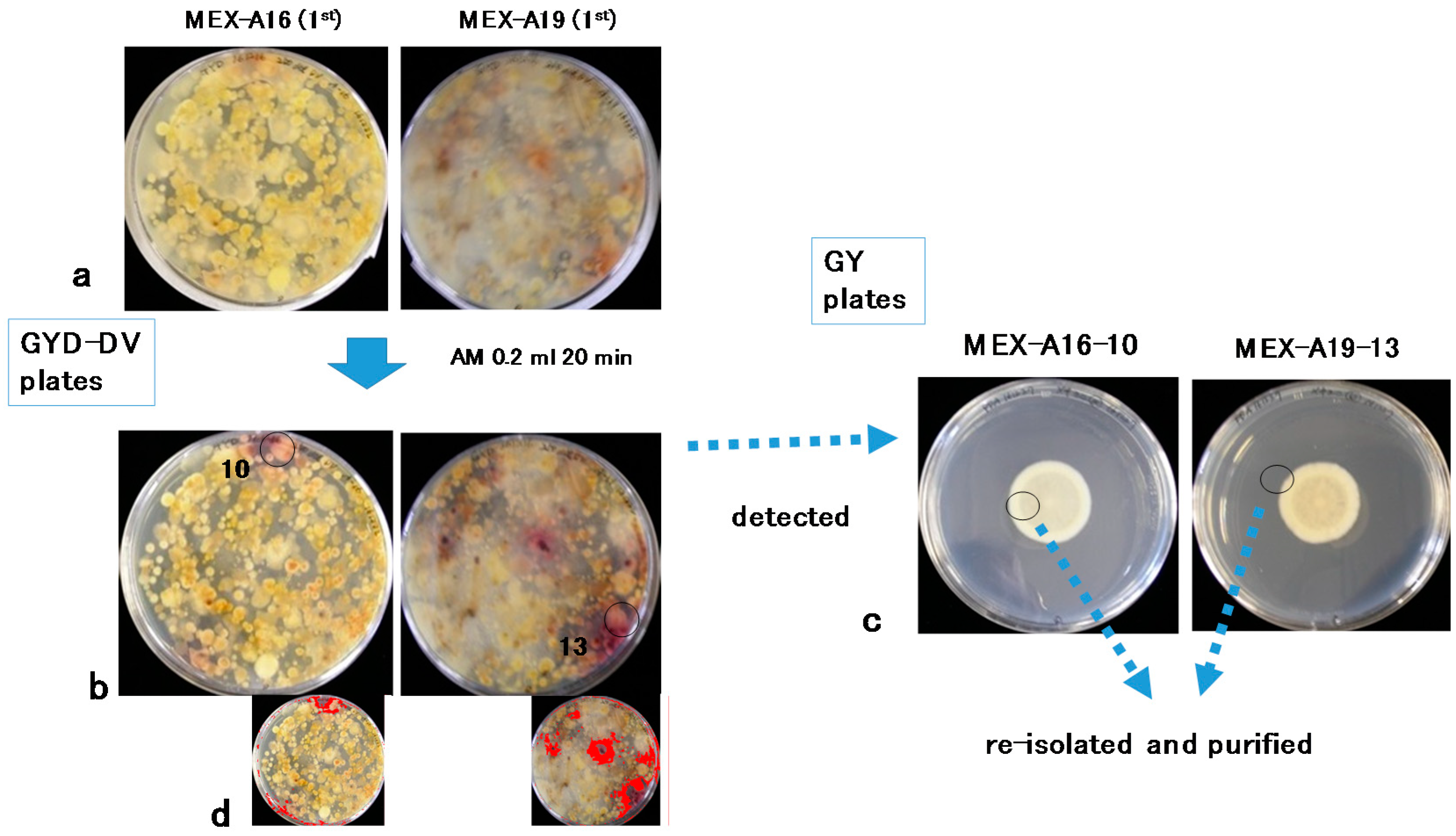
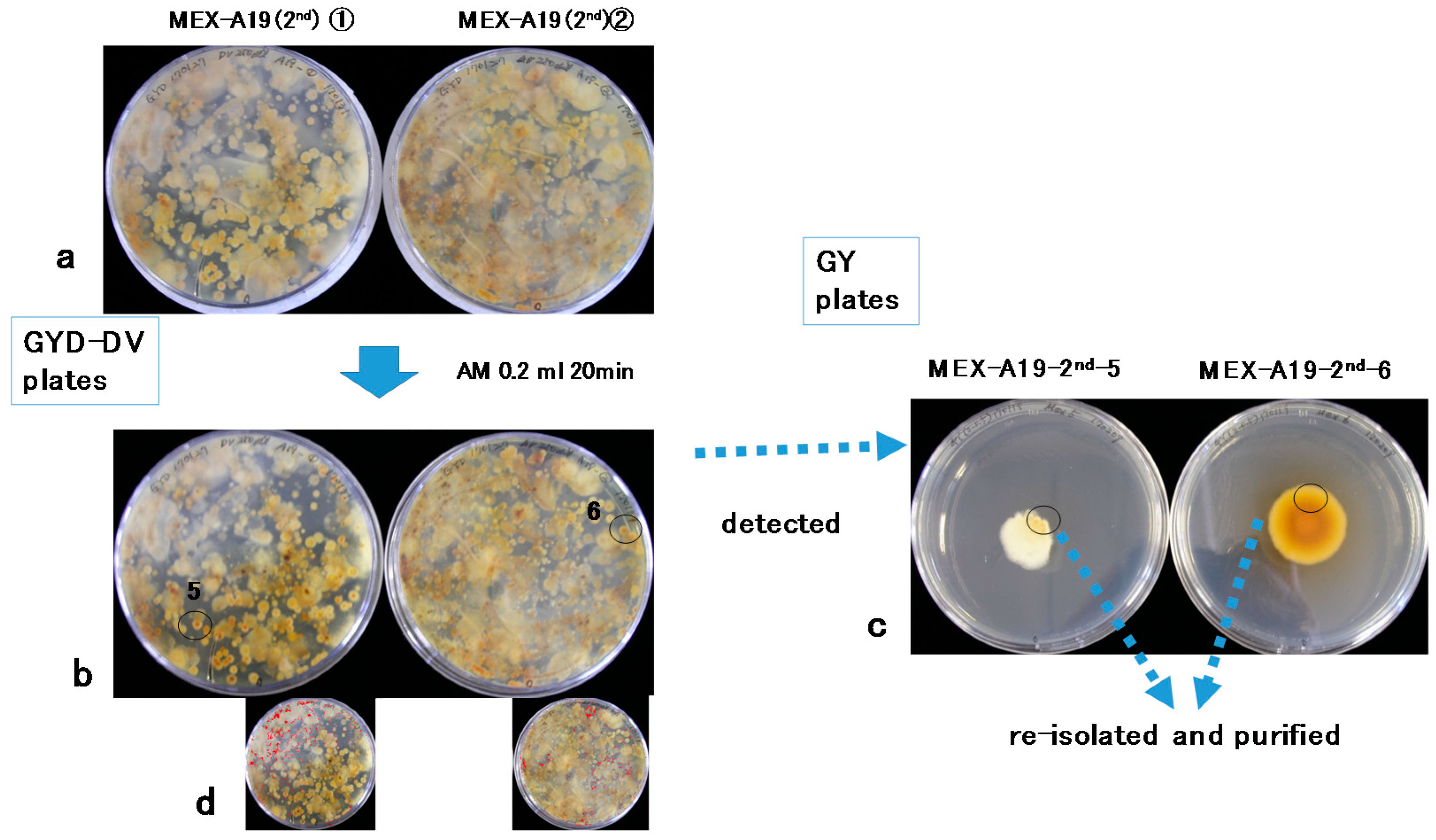
2.1. Isolation of Aspergilli from Soil Subsamples
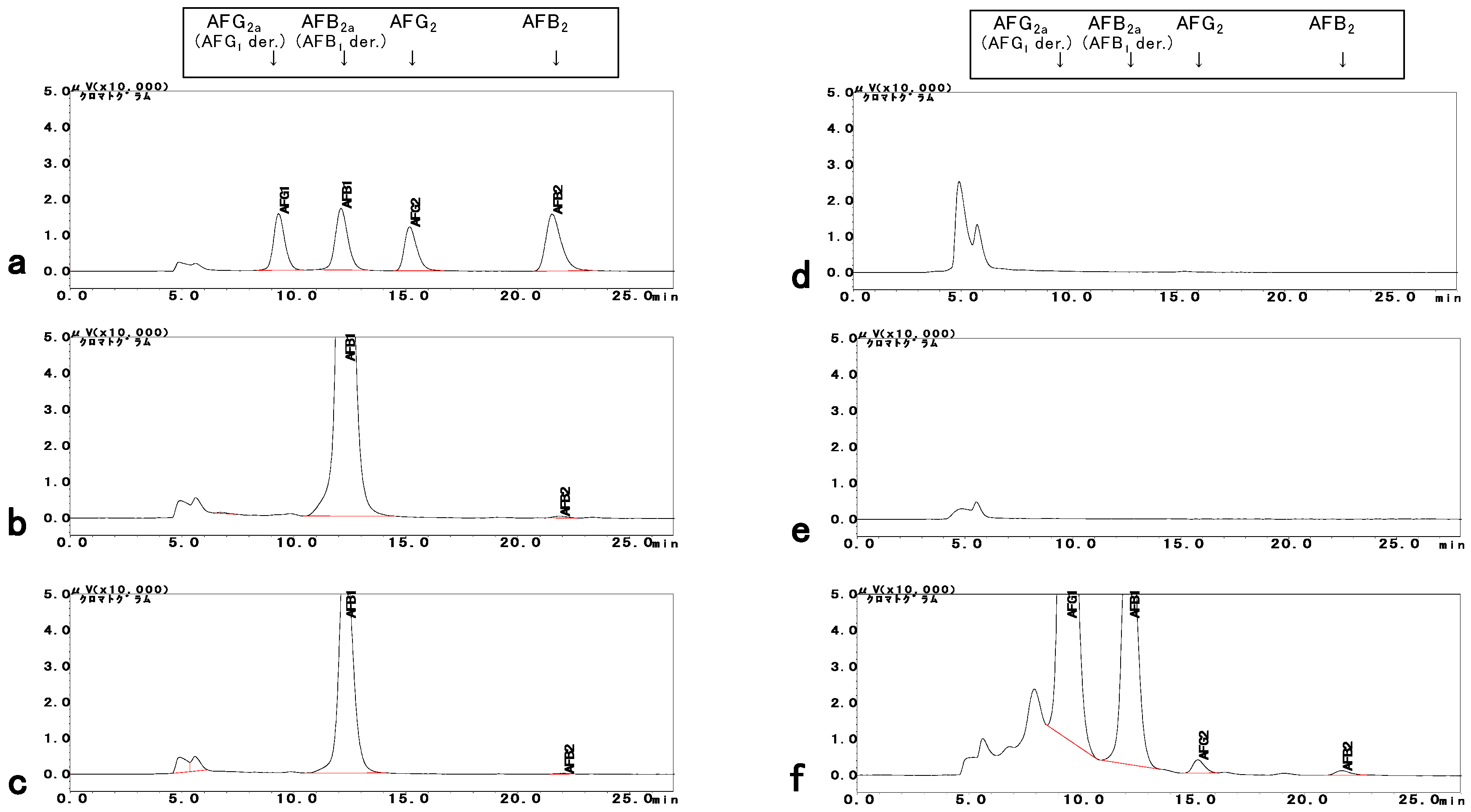
2.2. Determination of AFs
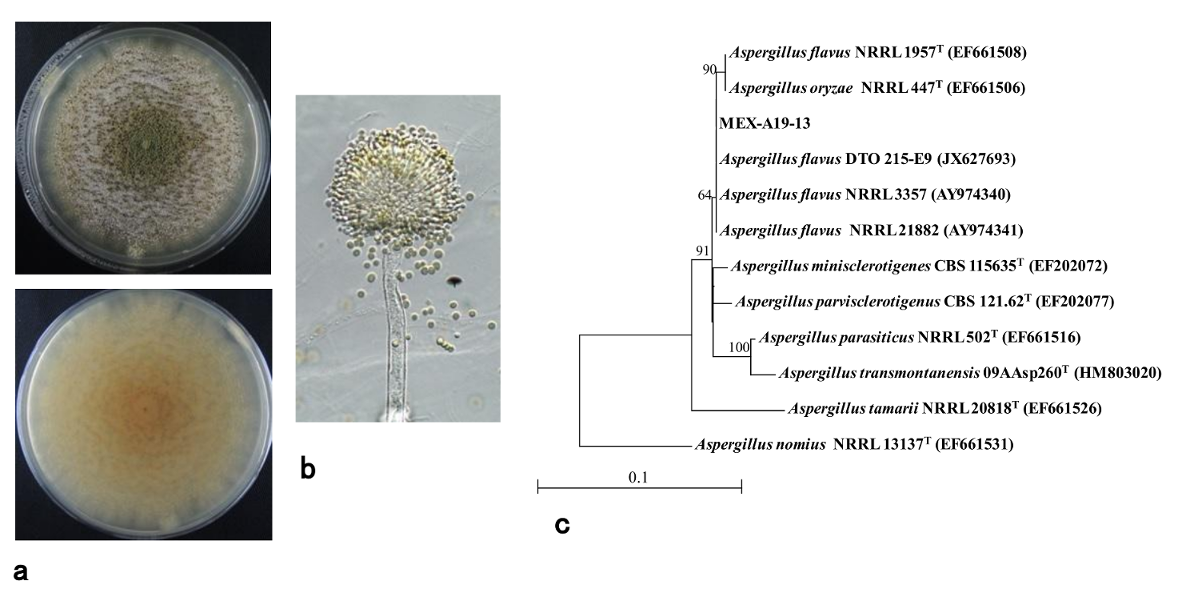
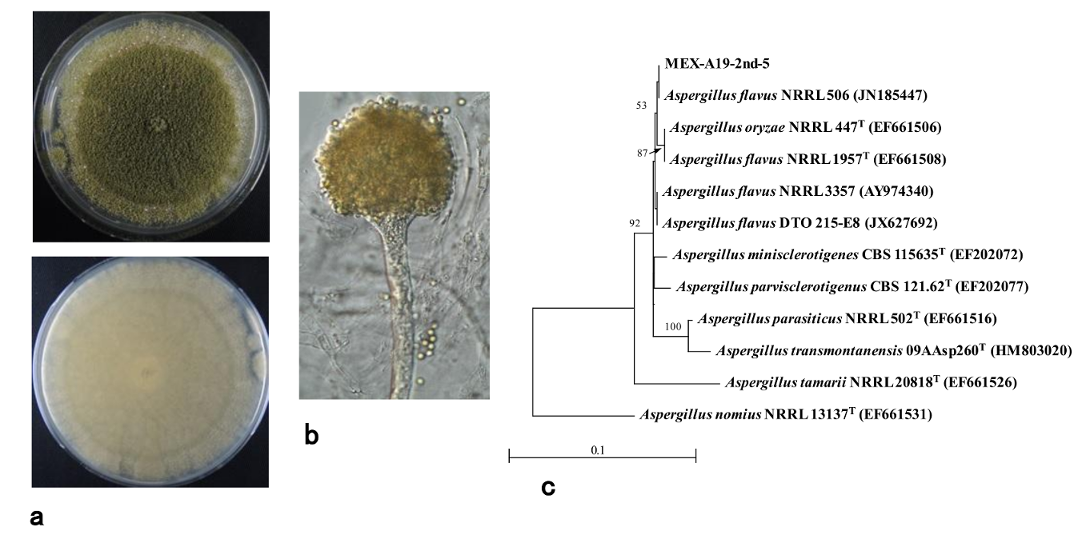
2.3. Phylogenetic Analysis of Isolates
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Media
5.2. Strains Used
5.3. Soil Sampling
5.4. DV–AM Method
5.5. AF Production Analysis
5.6. Identification of Fungal Species
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DV-AM | dichlorvos-ammonia |
| AF | aflatoxin |
| DV | dichlorvos |
| VOAc | versiconol acetate |
| VHA | versiconal hemiacetal acetate |
| AFPA | Aspergillus flavus and parasiticus agar |
| TFA | trifluoroacetic acid |
| HPLC | High-performance liquid chromatography |
| DNA | deoxyribonucleic acid |
| PCR | polymerase chain reaction |
| DDBJ | DNA Data Bank of Japan |
| EMBL | European Molecular Biology Laboratory |
References
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2016, 7, 2070. [Google Scholar] [CrossRef] [PubMed]
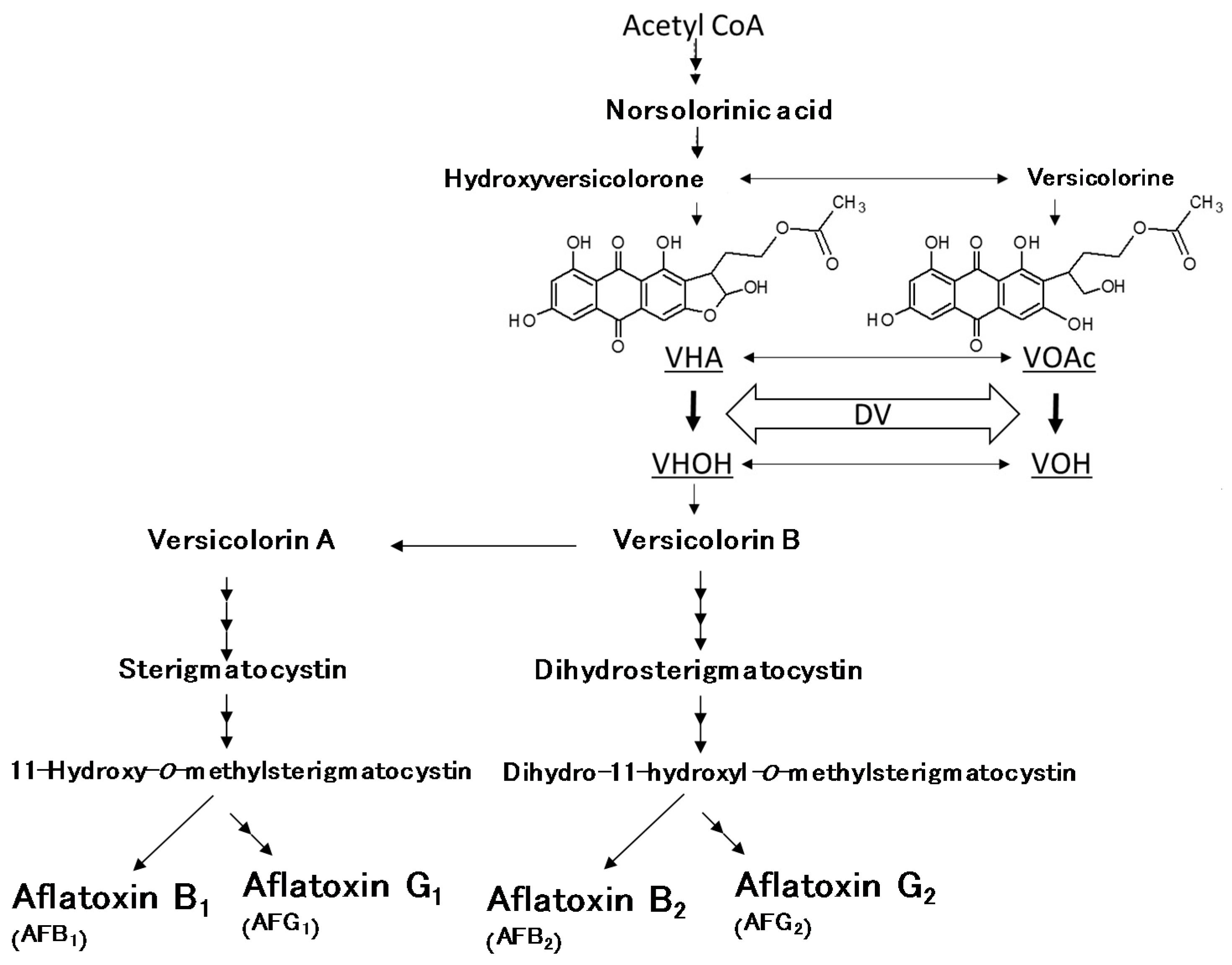
- Yabe, K.; Nakajima, H. Enzyme reactions and genes in aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2004, 64, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Yabe, K.; Chihaya, N.; Hatabayashi, H.; Kito, M.; Hoshino, S.; Zeng, H.; Cai, J.; Nakajima, H. Production of M-/GM-group aflatoxins catalyzed by the OrdA enzyme in aflatoxin biosynthesis. Fungal Genet. Biol. 2012, 49, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. Some Naturally Occurring Substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; WHO Press: Lyon, France, 1993; Volume 56, pp. 41–487. [Google Scholar]
- Guzmán-de-Peña, D.; Peña-Cabriales, J.J. Regulatory considerations of aflatoxin contamination of food in Mexico. Rev. Latinoam. Microbiol. 2005, 47, 160–164. [Google Scholar] [PubMed]
- Yabe, K.; Hatabayashi, H.; Ikehata, A.; Zheng, Y.; Kushiro, M. Development of the dichlorvos-ammonia (DV-AM) method for the visual detection of aflatoxigenic fungi. Appl. Microbiol. Biotechnol. 2015, 99, 10681–10694. [Google Scholar] [CrossRef] [PubMed]
- Yabe, K.; Chihaya, N.; Hamamatsu, S.; Sakuno, E.; Hamasaki, T.; Nakajima, H.; Bennett, J.W. Enzymatic conversion of averufin to hydroxyversicolorone and elucidation of a novel metabolic grid involved in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2003, 69, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Guclu, H. Aflatoxin regulations in a network of global maize trade. PLoS ONE 2012, 7, e45151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitt, J.I. Mycotoxins: Aflatoxins. In Encyclopedia for Food Safety; Motarjemi, Y., Gerald, M., Ewen, T., Eds.; Academic Press: Waltham, MA, USA, 2014; Volume 2, pp. 289–294. [Google Scholar]
- Horn, B.W.; Dorner, J.W. Regional differences in production of aflatoxin B1 and cyclopiazonic acid by soil isolates of Aspergillus flavus along a transect within the United States. Appl. Environ. Microbiol. 1999, 65, 1444–1449. [Google Scholar] [PubMed]
- Takahashi, H.; Kamimura, H.; Ichinoe, M. Distribution of aflatoxin-producing Aspergillus flavus and Aspergillus parasiticus in sugarcane fields in the southernmost islands of Japan. J. Food Prot. 2004, 67, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Beltran, A.; Jaime, R.; Cotty, P.J. Aflatoxin-producing fungi in maize field soils from sea level to over 2000 masl: A three year study in Sonora, Mexico. Fungal Biol. 2015, 119, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Magnussen, A.; Parsi, M.A. Aflatoxins, hepatocellular carcinoma and public health. World J. Gastroenterol. 2013, 19, 1508–1512. [Google Scholar] [CrossRef] [PubMed]
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect. 2005, 113, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.L.; Chen, Z.Y.; Cleveland, T.E.; Russin, J.S. Advances in the development of host resistance in corn to aflatoxin contamination by Aspergillus flavus. Phytopathology 1999, 82, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Munkvold, G.P. Cultural and genetic approaches to managing mycotoxins in maize. Annu. Rev. Phytopathol. 2003, 41, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Blandino, M.; Reyneri, A.; Vanara, F. Effect of sowing time on toxigenic fungal infection and mycotoxin contamination of maize kernels. J. Phytopathol. 2009, 157, 7–14. [Google Scholar] [CrossRef]
- Barros, G.; Magnoli, C.; Reynoso, M.; Ramirez, M.; Farnochi, M.; Torres, A.; Dalcero, M.; Sequeira, J.; Rubinstein, C.; Chulze, S. Fungal and mycotoxin contamination in Bt maize and non-Bt maize grown in Argentina. World Mycotoxin J. 2009, 2, 53–60. [Google Scholar] [CrossRef]
- Hawkins, L.K.; Windham, G.L.; Williams, W.P. Effect of different postharvest drying temperatures on Aspergillus flavus survival and aflatoxin content in five maize hybrids. J. Food Prot. 2005, 68, 1521–1524. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Brown, R.L.; Chen, Z.Y.; Menkir, A.; Yu, J.; Bhatnagar, D. Transcriptional profiles uncover Aspergillus flavus-induced resistance in maize kernels. Toxins 2011, 3, 766–786. [Google Scholar] [CrossRef] [PubMed]
- Cotty, P.J. Influence of field application of an atoxigenic strain of Aspergillus flavus on the populations of A. flavus infecting cotton bolls and on the aflatoxin content of cottonseed. Phytopathology 1994, 84, 1270–1277. [Google Scholar] [CrossRef]
- Dorner, J.W.; Cole, R.J.; Wicklow, D.T. Aflatoxin reduction in corn through field application of competitive fungi. J. Food Prot. 1999, 62, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, T.E.; Dowd, P.F.; Desjardins, A.E.; Bhatnagar, D.; Cotty, P.J. United States Department of Agriculture—Agricultural Research Service research on pre-harvest prevention of mycotoxins and mycotoxigenic fungi in US crops. Pest Manag. Sci. 2003, 59, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.K.; Shier, W.T.; Horn, B.W.; Weaver, M.A. Cultural methods for aflatoxin detection. J. Toxicol. Toxin Rev. 2004, 23, 295–315. [Google Scholar] [CrossRef]
- Suzuki, T.; Iwahashi, Y. Addition of carbon to the culture medium improves the detection efficiency of aflatoxin synthetic fungi. Toxins 2016, 8, 338. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I.; Hocking, A.D.; Glenn, D.R. An improved medium for the detection of Aspergillus flavus and A. parasiticus. J. Appl. Bacteriol. 1983, 54, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Bothast, R.J.; Fennell, D.I. A medium for rapid identification and enumeration of Aspergillus flavus and related organisms. Mycologia 1974, 66, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.C.; Hsieh, D.P.H. Step of dichlorvos inhibition in the pathway of aflatoxin biosynthesis. Appl. Microbiol. 1974, 28, 52–57. [Google Scholar] [PubMed]
- Kushiro, M.; Hatabayashi, H.; Nakagawa, H.; Yabe, K. Isolation of minor aflatoxigenic fungi using dichlorvos-ammonia (DV-AM) method. JSM Mycotoxins 2018, 68, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Kushiro, M.; Hatabayashi, H.; Zheng, Y.; Yabe, K. Application of newly-developed dichlorvos–ammonia (DV–AM) method to direct isolation of aflatoxigenic fungi from field soils. Mycoscience 2017, 58, 85–94. [Google Scholar] [CrossRef]
- Hong, S.B.; Go, S.J.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Aspergillus fumigatus and related species. Mycologia 2005, 97, 1316–1329. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kushiro, M.; Hatabayashi, H.; Yabe, K.; Loladze, A. Detection of Aflatoxigenic and Atoxigenic Mexican Aspergillus Strains by the Dichlorvos–Ammonia (DV–AM) Method. Toxins 2018, 10, 263. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10070263
Kushiro M, Hatabayashi H, Yabe K, Loladze A. Detection of Aflatoxigenic and Atoxigenic Mexican Aspergillus Strains by the Dichlorvos–Ammonia (DV–AM) Method. Toxins. 2018; 10(7):263. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10070263
Chicago/Turabian StyleKushiro, Masayo, Hidemi Hatabayashi, Kimiko Yabe, and Alexander Loladze. 2018. "Detection of Aflatoxigenic and Atoxigenic Mexican Aspergillus Strains by the Dichlorvos–Ammonia (DV–AM) Method" Toxins 10, no. 7: 263. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins10070263