Clostridium botulinum and Clostridium perfringens Occurrence in Kazakh Honey Samples

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Culture Process
2.1.1. In-House Validation of Culture Enrichment Method
2.1.2. Culture Identification
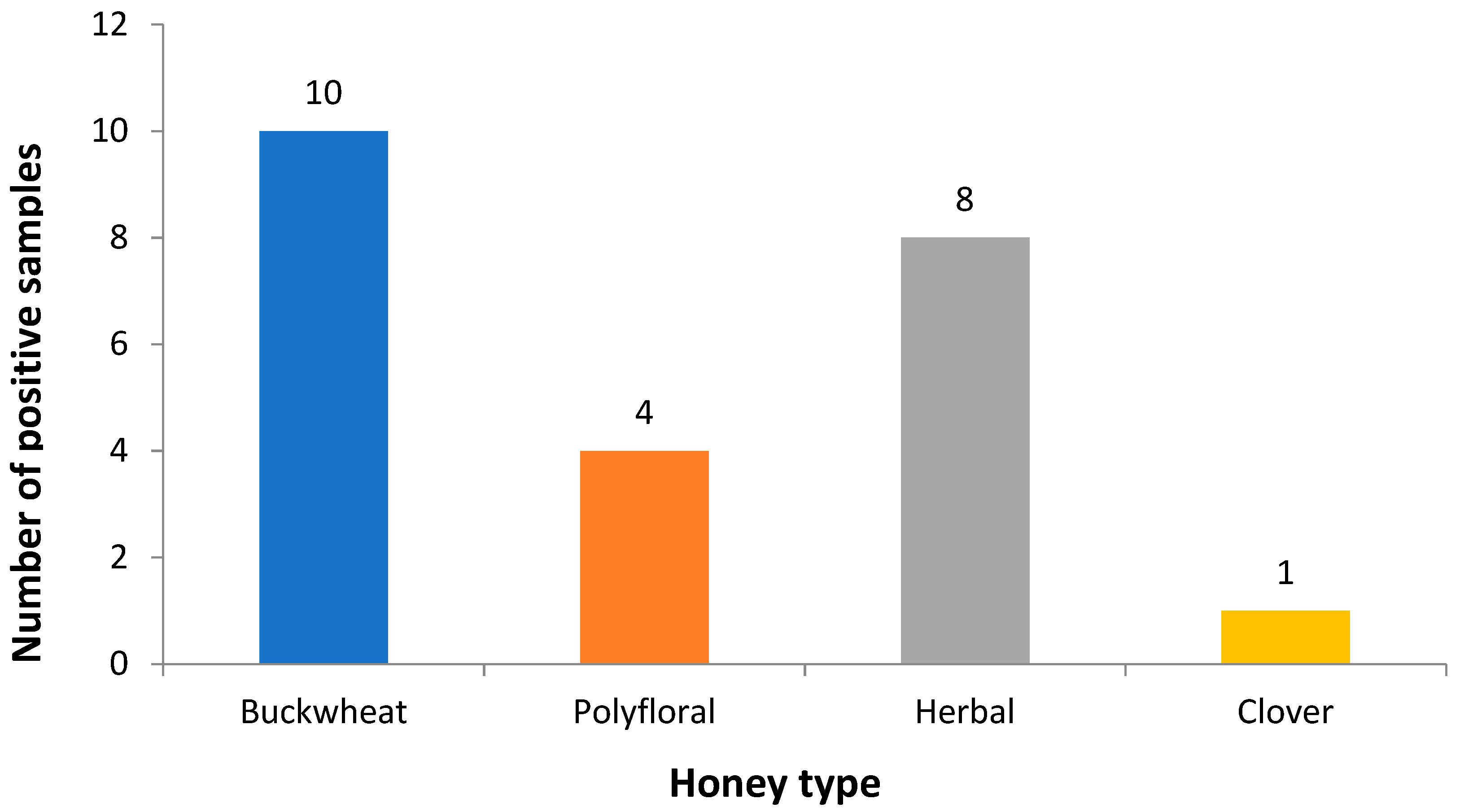
2.2. Clostridium Botulinum Detection
2.2.1. Real-Time PCR Analysis Results
2.2.2. mPCR for Bont Genes Detection
2.3. Clostridium Perfringens Detection
2.4. Detection of Other Spore-Forming Anaerobes by Sequence Analysis of 16S rDNA Amplicons
3. Discussion
4. Materials and Methods
4.1. Honey Samples
4.2. Enrichment Cultures
In-House Validation of Culture Enrichment Process
4.3. Nucleic Acids Isolation
DNA Isolation
4.4. Clostridium Botulinum Identification
4.4.1. ntnh Gene Detection by Real-Time PCR
4.4.2. Bont/A, Bont/B, Bont/E, and Bont/F GENES detection
4.5. Clostridium Perfringens Detection
4.6. Detection of Other Spore-Forming Anaerobes by Amplification and Sequencing of 16S rDNA
4.7. Electrophoresis of PCR Products
Author Contributions
Funding
Conflicts of Interest
References
- Nevas, M.; Lindström, M.; Hautamäki, K.; Puoskari, S.; Korkeala, H. Prevalence and diversity of Clostridium botulinum types A, B, E and F in honey produced in the Nordic countries. Int. J. Food Microbiol. 2005, 105, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, P.B.; Adeleke, O.E.; Ola, I.O. Honey: A reservoir for microorganisms and an inhibitory agent for microbes. Afr. Health Sci. 2007, 7, 159–165. [Google Scholar] [PubMed]
- Sereia, M.J.; Alves, E.M.; de Toledo, V.A.A.; Marchini, L.C.; Faquinello, P.; Sekine, E.S.; Wielewski, P. Microbial flora in organic honey samples of Africanized honey bees from Parana River islands. Food Sci. Tech.-Braz. 2017, 31, 462–466. [Google Scholar] [CrossRef]
- Tomassetti, F.; Milito, M.; Dell’Aira, E.; de Santis, L.; Migliore, G.; Formato, G. Microbiological comparison between honey in jar and honey in comb for human. Ital. J. Food Saf. 2009, 3, 65–66. [Google Scholar] [CrossRef]
- Wen, Y.; Wang, L.; Jin, Y.; Zhang, J.; Su, L.; Zhang, X.; Zhou, J.; Li, Y. The microbial community dynamics during the vitex honey ripening process in the honeycomb. Front. Microbiol. 2017, 8, 1649. [Google Scholar] [CrossRef] [PubMed]
- Aureli, P.; Franciosa, G.; Fenicia, L. Infant botulism and honey in Europe: A commentary. Pediatr. Infect. Dis. J. 2002, 21, 866–868. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, C.O.; Ayubi, A.; Zulfiquer, F.; Santhanam, G.; Ahmed, M.A.S.; Deeb, J. Infant botulism following honey ingestion. BMJ Case Rep. 2012. [Google Scholar] [CrossRef] [PubMed]
- Midura, T.F. Update: Infant Botulism. Clin. Microbiol. Rev. 1996, 9, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Midura, T.F.; Arnon, S.S. Infant botulism. Identification of Clostridium botulinum and its toxins in faeces. Lancet 1976, 2, 934–936. [Google Scholar] [CrossRef]
- Pickett, J.; Berg, B.; Chaplin, E.; Brunstetter-Shafer, M.A. Syndrome of botulism in infancy: Clinical and electrophysiologic study. N. Engl. J. Med. 1976, 295, 770–772. [Google Scholar] [CrossRef]
- Fenicia, L.; Anniballi, F. Infant botulism. Ann. Ist. Super. Sanità 2009, 45, 134–146. [Google Scholar] [PubMed]
- Weissenstein, A.; Villalon, G.; Luchter, E.; Bittmann, S. Medical honey and the role in pediatric emergency wound management. J. Pediatr. Dis. 2018, 2, 1–7. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 141. [Google Scholar] [CrossRef] [PubMed]
- McClane, B.A. Clostridium perfringens. In Food Microbiology: Fundamentals and Frontiers; Doyle, M.P., Beuchat, L.R., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 423–444. [Google Scholar]
- Brynestad, S.; Granum, P.E. Clostridium perfringens and foodborne infections. Int. J. Food Microbiol. 2002, 74, 195–202. [Google Scholar] [CrossRef]
- Labbe, R.G.; Juneja, V.K. Clostridium perfringens gastroenteritis. In Foodborne Infections and Intoxications; Riemann, H.P., Cliver, D.O., Eds.; Academic Press: San Diego, CA, USA, 2006; pp. 137–163. [Google Scholar]
- Grass, J.E.; Gould, L.H.; Mahon, B.E. Epidemiology of foodborne disease outbreaks caused by Clostridium perfringens, United States, 1998–2010. Foodborne Pathog. Dis. 2013, 10, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Saputo, J.; Sayeed, S.; Vidal, J.E.; Fisher, D.J.; Poon, R.; Adams, V.; Fernandez-Miyakawa, M.E.; Rood, J.I.; McClane, B.A. Development and application of new mouse models to study the pathogenesis of Clostridium perfringens type C Enterotoxemias. Infect. Immun. 2009, 77, 5291–5299. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Chen, F.; Leng, X.; Fei, R.; Wang, L. Toxinotyping of Clostridium perfringens fecal isolates of reintroduced P`ere David’s deer (Elaphurus davidianus) in China. J. Wildl. Dis. 2014, 50, 942–945. [Google Scholar] [CrossRef]
- Mustafina, R.; Maikanov, B.; Wiśniewski, J.; Tracz, M.; Anusz, K.; Grenda, T.; Kukier, E.; Goldsztejn, M.; Kwiatek, K. Contamination of honey produced in the Republic of Kazakhstan with Clostridium botulinum. Bull. Vet. Inst. Pulawy 2015, 59, 241–246. [Google Scholar] [CrossRef]
- Nakano, H.; Sakaguchi, G. An unusually heavy contamination of honey products by Clostridium botulinum type F and Bacillus alvei. FEMS Microbiol. Lett. 1991, 63, 171–177. [Google Scholar] [CrossRef]
- Küplülü, Ö.; Göncüoğlü, M.; Özdemir, H.; Koluman, A. Incidence of Clostridium botulinum spores in honey in Turkey. Food Control 2006, 17, 222–224. [Google Scholar] [CrossRef]
- Grenda, T.; Grabczak, M.; Sieradzki, Z.; Kwiatek, K.; Pohorecka, K.; Skubida, M.; Bober, A. Clostridium botulinum spores in Polish honey samples. J. Vet. Sci. 2018, 19, 635–642. [Google Scholar] [CrossRef]
- Wojtacka, J.; Wysok, B.; Lipiński, Z.; Gomółka-Pawlicka, M.; Rybak-Chmielewska, H.; Wiszniewska-Łaszczych, A. Clostridium botulinum spores found in honey from small apiaries in Poland. J. Apic. Sci. 2016, 60, 89–100. [Google Scholar] [CrossRef]
- Wojtacka, J.; Wysok, B.; Kabašinskienė, A.; Wiszniewska-Łaszczych, A.; Gomółka-Pawlicka, M.; Szteyn, J.; Malakauskas, M.; Migowska-Calik, A. Prevalence of Clostridium botulinum Type A, B, E and F Isolated from Directly Sold Honey in Lithuania. J. Agric. Sci. Technol. 2017, 19, 335–343. [Google Scholar]
- Lindström, M.; Korkeala, H. Laboratory diagnostics of botulism. Clin. Microbiol. Rev. 2006, 19, 298–314. [Google Scholar] [CrossRef]
- Różańska, H. Microbial quality of Polish honey. Bull. Vet. Inst. Pulawy 2011, 55, 443–445. [Google Scholar]
- Midura, T.F.; Snowden, S.; Wood, R.M.; Arnon, S.S. Isolation of Clostridium botulinum from honey. J. Clin. Microbiol. 1979, 9, 282–283. [Google Scholar]
- Nakano, H.; Okabe, T.; Hashimoto, M.; Sakaguchi, G. Incidence of Clostridium botulinum in honey of various origins. Jpn. J. Med Sci. Biol. 1990, 43, 183–195. [Google Scholar] [CrossRef]
- Huhtanen, C.N.; Knox, D.; Shimanuki, H. Incidence and origin of Clostridium botulinum spores in honey. J. Food Prot. 1981, 44, 812–814. [Google Scholar] [CrossRef]
- Schocken-Iturino, R.P.; Carneiro, M.C.; Kato, E.; Sorbara, J.O.B.; Rossi, O.D.; Gerbasi, L.E.R. Study of the presence of the spores of Clostridium botulinum in honey in Brazil. FEMS Immunol. Med. Microbiol. 1999, 24, 379–382. [Google Scholar] [CrossRef]
- Grenda, T.; Kukier, E.; Sieradzki, Z.; Goldsztejn, M.; Kwiatek, K. In-house validation of multiplex PCR method for detection of Clostridium botulinum in food and feed. Bull. Vet. Inst. Pulawy 2012, 56, 155–160. [Google Scholar] [CrossRef]
- Grenda, T.; Kwiatek, K. In-house validation of real-time PCR method for detection of Clostridium botulinum in food and feed matrixes. Bull. Vet. Inst. Pulawy 2010, 54, 557–562. [Google Scholar]
- Raphael, B.H.; Andreadis, J.D. Real-time PCR detection of the nontoxic nonhemagglutinin gene as a rapid screening method for bacterial isolates harboring the botulinum neurotoxin (A–G) gene complex. J. Microbiol. Methods 2007, 71, 343–346. [Google Scholar] [CrossRef]
- Carter, A.T.; Peck, M.W. Genomes, neurotoxins and biology of Clostridium botulinum Group I and Group II. Res. Microbiol. 2015, 166, 303–317. [Google Scholar] [CrossRef]
- Collins, M.D.; East, A.K. Phylogeny and taxonomy of the food-borne pathogen Clostridium botulinum and its neurotoxins. J. Appl. Microbiol. 1998, 84, 5–17. [Google Scholar] [CrossRef]
- Scalfaro, C.; Auricchio, B.; De Medici, D.; Anniballi, F. Foodborne botulism: An evolving public health challenge. Infect. Dis. (Lond.) 2019, 51, 97–101. [Google Scholar] [CrossRef]
- Skarin, H.; Segerman, B. Horizontal gene transfer of toxin genes in Clostridium botulinum. Mob. Genet. Elem. 2011, 1, 213–215. [Google Scholar] [CrossRef]
- Wentz, T.G.; Muruvanda, T.; Lomonaco, S.; Thirunavukkarasu, N.; Hoffmann, M.; Allard, M.W.; Hodge, D.R.; Pillai, S.P.; Hammack, T.S.; Brown, E.W.; et al. Closed genome sequence of Chryseobacterium piperi strain CTMT/ATCC BAA-1782, a Gram-negative bacterium with clostridial neurotoxin-like coding sequences. Genome Announc. 2017, 5, e01296-17. [Google Scholar] [CrossRef]
- Zhang, S.; Lebreton, F.; Mansfield, M.J.; Miyashita, S.I.; Zhang, J.; Schwartzman, J.A.; Tao, L.; Masuyer, G.; Martinez-Carranza, M.; Stenmark, P.; et al. Emergence of a botulinum neurotoxin-like toxin in a commensal strain of Enterococcus faecium. Cell Host Microbe 2018, 23, 169–176. [Google Scholar] [CrossRef]
- Popoff, M.R. Botulinum neurotoxins: Still a privilege of clostridia? Cell Host Microbe 2018, 23, 145–146. [Google Scholar] [CrossRef]
- Poulain, B.; Popoff, M.R. Why are botulinum neurotoxin-producing bacteria so diverse and botulinum neurotoxins so toxic? Toxins 2019, 11, 34. [Google Scholar] [CrossRef]
- Li, L.; Chen, X.; van der Tol, C.; Luo, G.; Su, Z. Growing season net ecosystem CO2 exchange of two desert ecosystems with alkaline soils in Kazakhstan. Ecol. Evol. 2014, 4, 14–26. [Google Scholar] [CrossRef]
- Kazakhstan in 2009. Statistical Yearbook of Kazakhstan. Statistic Agency of Republic of Kazakhstan, Astana, 2010. Available online: http://www.stat.kz/publishing/DocLib/2010/Ewegodnik_2010.pdf (accessed on 20 June 2019).
- Grenda, T.; Grabczak, M.; Goldsztejn, M.; Kozieł, N.; Kwiatek, K.; Pohorecka, K.; Skubida, M.; Bober, A. Clostridium perfringens spores in Polish honey samples. J. Vet. Res. 2018, 62, 281–284. [Google Scholar] [CrossRef]
- Kiu, R.; Caim, S.; Alexander, S.; Pachori, P.; Hall, L.J. Probing genomic aspects of the multi-host pathogen Clostridium perfringens reveals significant pangenome diversity, and a diverse array of virulence factors. Front. Microbiol. 2017, 8, 2485. [Google Scholar] [CrossRef]
- Tlenchieva, T.M.; Sultankulova, K.T.; Shoraeva, K.A.; Strochkov, V.M.; Orynbaev, M.B.; Zaitsev, V.L.; Rystaeva, R.A.; Sandybaev, N.T.; Sansyzbai, A.R. Detection and characteristics of alphatoxin strain Clostridium perfringens genetic variability, isolated from Kazakhstan Saiga in 2010–2013. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1449–1458. [Google Scholar]
- Li, M.; Zhang, X.; Zhu, L.; Wang, H.; Zhao, N.; Luo, J.; Wang, C.; Wang, Y.; Liu, Y.; Zhou, W.; et al. Identification, isolation, and phylogenetic analysis of Clostridium perfringens type A and type C from Wild Boar (Sus scrofa) in the People’s Republic of China. J. Wildl. Dis. 2017, 53, 642–648. [Google Scholar] [CrossRef]
- Matches, J.R.; Liston, J.; Curran, D. Clostridium perfringens in the environment. Appl. Microbiol. 1974, 28, 655–660. [Google Scholar]
- Fletcher, G.C.; Youssef, J.F.; Lu, G. Selecting Methods for Determining the Presence of BoNT Genes in New Zealand Marine Sediments; Crop & Food Research Confidential Report; New Zealand Food Safety Authority (NZFSA): Wellington, New Zealand, 2008.
- De Medici, D.; Anniballi, F.; Wyatt, G.M.; Lindström, M.; Messelhäußer, U.; Aldus, C.F.; Delibato, E.; Korkeala, H.; Peck, M.W.; Fenicia, L. Multiplex PCR for detection of botulinum neurotoxin-producing Clostridia in clinical, food, and environmental samples. Appl. Environ. Microbiol. 2009, 75, 6457–6461. [Google Scholar] [CrossRef]
- Baums, C.G.; Schotte, U.; Amtsberg, G.; Goethe, R. Diagnostic multiplex PCR for toxin genotyping of Clostridium perfringens isolates. Vet. Microbiol. 2004, 100, 11–16. [Google Scholar] [CrossRef]
- Kukier, E.; Kwiatek, K. Occurrence of Clostridium perfringens in food chain. Bull. Vet. Inst. Pulawy 2010, 54, 571–576. [Google Scholar]
- Vaneechoutte, M.; Cartwright, C.P.; Williams, E.C.; Jäger, B.; Tichy, H.V.; de Baere, T.; de Rouck, A.; Verschraegen, G. Evaluation of 16S rRNA gene restriction analysis for the identification of cultured organisms of clinically important Clostridium species. Anaerobe 1996, 4, 249–256. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
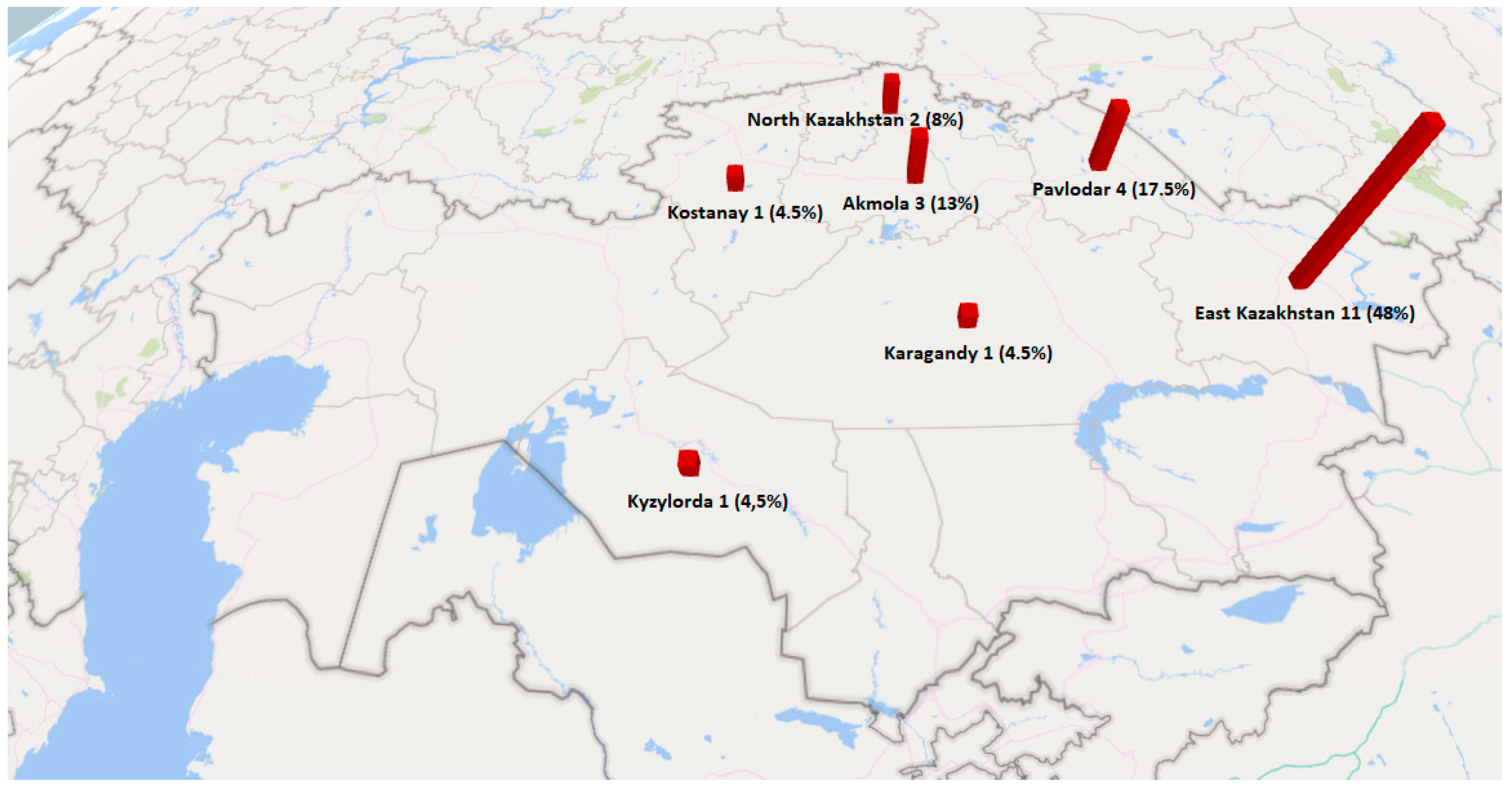
| Province | Number of C. botulinum Isolates (Honey Type) | Number of Isolates Related to C. botulinum Species (Honey Type) | Number of C. perfringens Isolates (Honey Type) | Total in Provinces (%) |
|---|---|---|---|---|
| East Kazakhstan | 1 (Buckwheat) | 1 (Herbal) | 4 (Buckwheat), 2 (Polyfloral), 3 (Herbal) | 11 (48%) |
| Almaty | - | - | - | - |
| Pavlodar | - | - | 2 (Buckwheat), 1 (Clover), 1 (Polyfloral) | 4 (17.5%) |
| Akmola | - | 2 (Buckwheat) | 1 (Herbal) | 3 (13%) |
| Kostanay | - | 1 (Buckwheat) | - | 1 (4.5%) |
| Karagandy | - | - | 1 (Herbal) | 1 (4.5%) |
| North Kazakhstan | - | - | 1 (Polyfloral), 1 (Herbal) | 2 (8.5%) |
| Kyzylorda | - | - | 1 (Herbal) | 1 (4.5%) |
| South Kazakhstan | - | - | - | - |
| Zhambyl | - | - | - | - |
| West Kazakhstan | - | - | - | - |
| Total | 1 (4.5%) | 4 (17.5%) | 18 (78%) | 23 (100%) |
| Province | Number of Suspected Isolates | Number of Isolates Carrying Particular Genes | ||||
|---|---|---|---|---|---|---|
| ntnh | bont/A | bont/B | bont/E | bont/F | ||
| East Kazakhstan | 2 | 1 | 1 | - | - | - |
| Akmola | 2 | - | - | - | - | - |
| Kostanay | 1 | - | - | - | - | - |
| Total | 5 | 1 | 1 | - | - | - |
| Province | Number of Suspected Isolates | Number of Isolates Carrying Particular Genes | |||||
|---|---|---|---|---|---|---|---|
| cpa | cpb | etx | iap | cpe | cpb2 | ||
| East Kazakhstan | 9 | 9 | - | - | - | - | - |
| Pavlodar | 4 | 4 | - | - | - | - | - |
| Akmola | 1 | 1 | - | - | - | - | - |
| Karagandy | 1 | 1 | - | - | - | - | - |
| North Kazakhstan | 2 | 2 | - | - | - | - | - |
| Kyzylorda | 1 | 1 | - | - | - | - | - |
| Total | 18 | 18 | - | - | - | - | - |
| Isolate | Similar Sequence from GenBank (NCBI) | Sequence ID | % Similarity | Origin |
|---|---|---|---|---|
| K2 | Clostridium sporogenes strain A4 16S rRNA | KY910419.1 | 98% | Akmola province/buckwheat honey |
| K3 | Clostridium sporogenes strain 1101M 16S rRNA | KU950270.1 | 99% | Akmola province/buckwheat honey |
| K4 | Clostridium beijerinckii strain JCM 7837 16S rRNA | LC258130.1 | 95% | East Kazakhstan province/herbal honey |
| K5 | Clostridium beijerinckii strain JCM 7837 16S rRNA | LC258130.1 | 94% | Kostanay province/buckwheat honey |
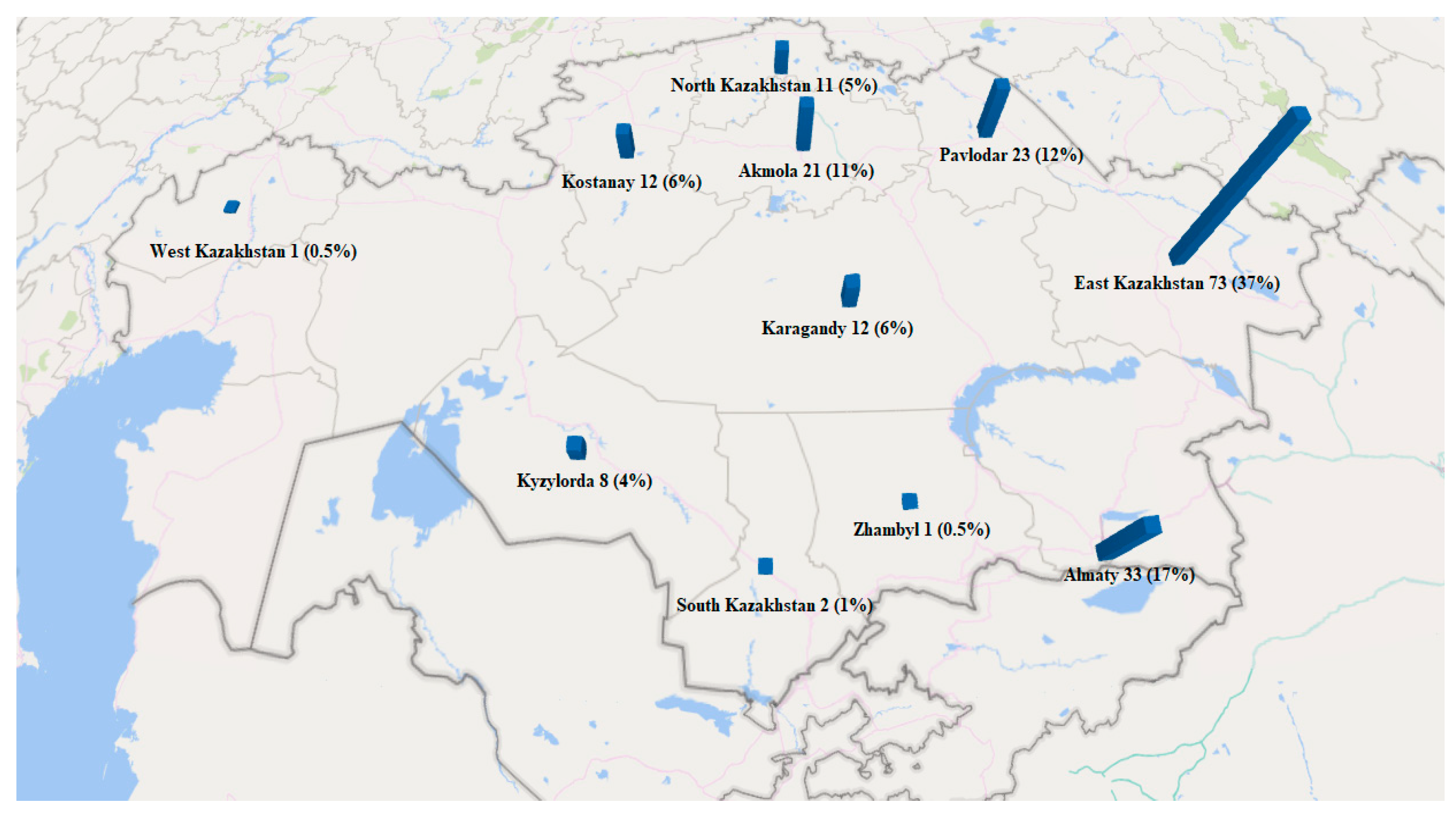
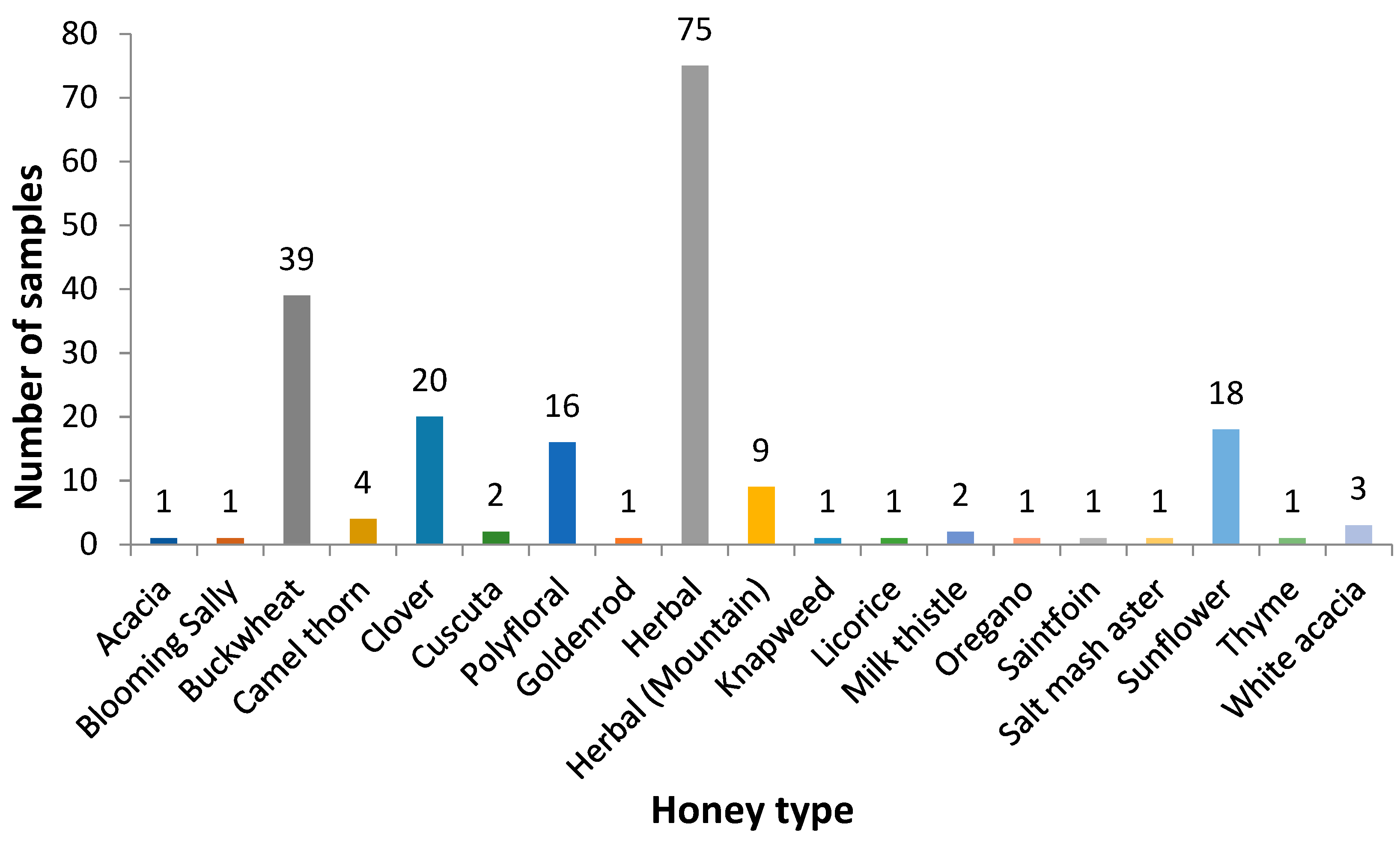
| Province | Number of Samples | Honey Type |
|---|---|---|
| East Kazakhstan | 73 (37%) | Acacia (1), Buckwheat (19), Clover (5), Polyfloral (4), Herbal (31), Herbal (Mountain) (2), Milk thistle (1), Oregano (1), Sunflower (7), White acacia (2) |
| Almaty | 33 (17%) | Buckwheat (2), Camel thorn (2), Clover (3), Polyfloral (1), Herbal (14), Herbal (Mountain) (7), Salt marsh aster (1), Sunflower (3) |
| Pavlodar | 23 (12%) | Buckwheat (7), Clover (5), Polyfloral (2), Herbal (5), Sunflower (4) |
| Akmola | 21 (11%) | Buckwheat (4), Clover (3), Polyfloral (2), Herbal (7), Sainfoin (1), Sunflower (2), White acacia (1), Camel thorn (1) |
| Kostanay | 12 (6%) | Buckwheat (2), Clover (1), Polyfloral (2), Herbal (6), Sunflower (1) |
| Karagandy | 12 (6%) | Buckwheat (3), Clover (1), Polyfloral (2), Goldenrod (1), Herbal (3), Milk thistle (1), Thyme (1) |
| North Kazakhstan | 11 (5%) | Blooming Sally (1), Buckwheat (2), Clover (1), Polyfloral (1), Herbal (5), Sunflower (1) |
| Kyzylorda | 8 (4%) | Camel thorn (1), Cuscuta (2), Polyfloral (1), Herbal (3), Licorice (1) |
| South Kazakhstan | 2 (1%) | Clover (1), Polyfloral (1) |
| Zhambyl | 1 (0.5%) | Knapweed (1) |
| West Kazakhstan | 1 (0.5%) | Herbal (1) |
| Total | 197 (100%) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maikanov, B.; Mustafina, R.; Auteleyeva, L.; Wiśniewski, J.; Anusz, K.; Grenda, T.; Kwiatek, K.; Goldsztejn, M.; Grabczak, M. Clostridium botulinum and Clostridium perfringens Occurrence in Kazakh Honey Samples. Toxins 2019, 11, 472. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11080472
Maikanov B, Mustafina R, Auteleyeva L, Wiśniewski J, Anusz K, Grenda T, Kwiatek K, Goldsztejn M, Grabczak M. Clostridium botulinum and Clostridium perfringens Occurrence in Kazakh Honey Samples. Toxins. 2019; 11(8):472. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11080472
Chicago/Turabian StyleMaikanov, Balgabay, Raikhan Mustafina, Laura Auteleyeva, Jan Wiśniewski, Krzysztof Anusz, Tomasz Grenda, Krzysztof Kwiatek, Magdalena Goldsztejn, and Magdalena Grabczak. 2019. "Clostridium botulinum and Clostridium perfringens Occurrence in Kazakh Honey Samples" Toxins 11, no. 8: 472. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11080472