Effects of Microcystin-LR on the Microstructure and Inflammation-Related Factors of Jejunum in Mice


Abstract
:1. Introduction
2. Results
2.1. Appearance and Length of Small Intestine of Mice
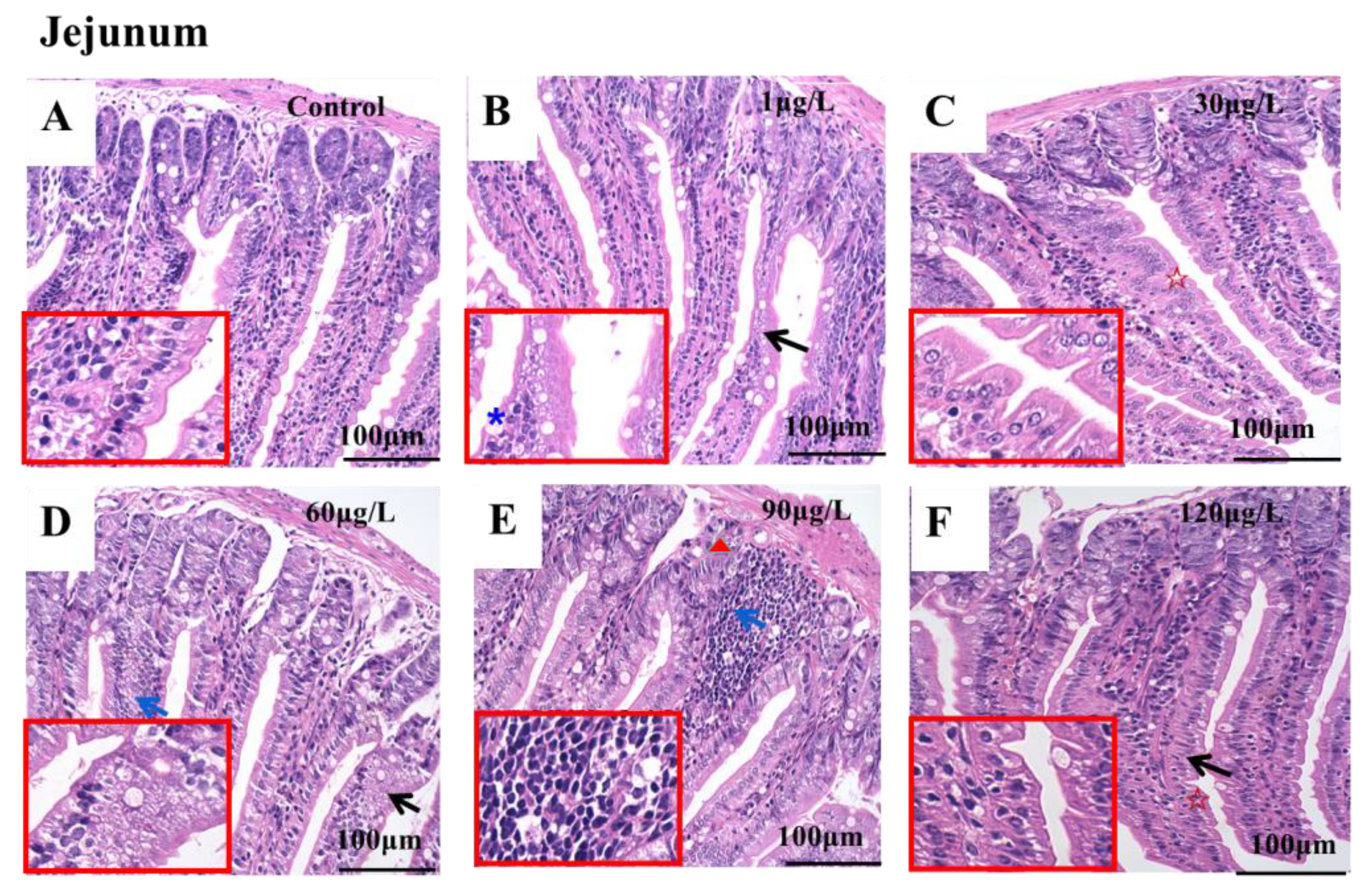
2.2. Microstructure of Jejunum
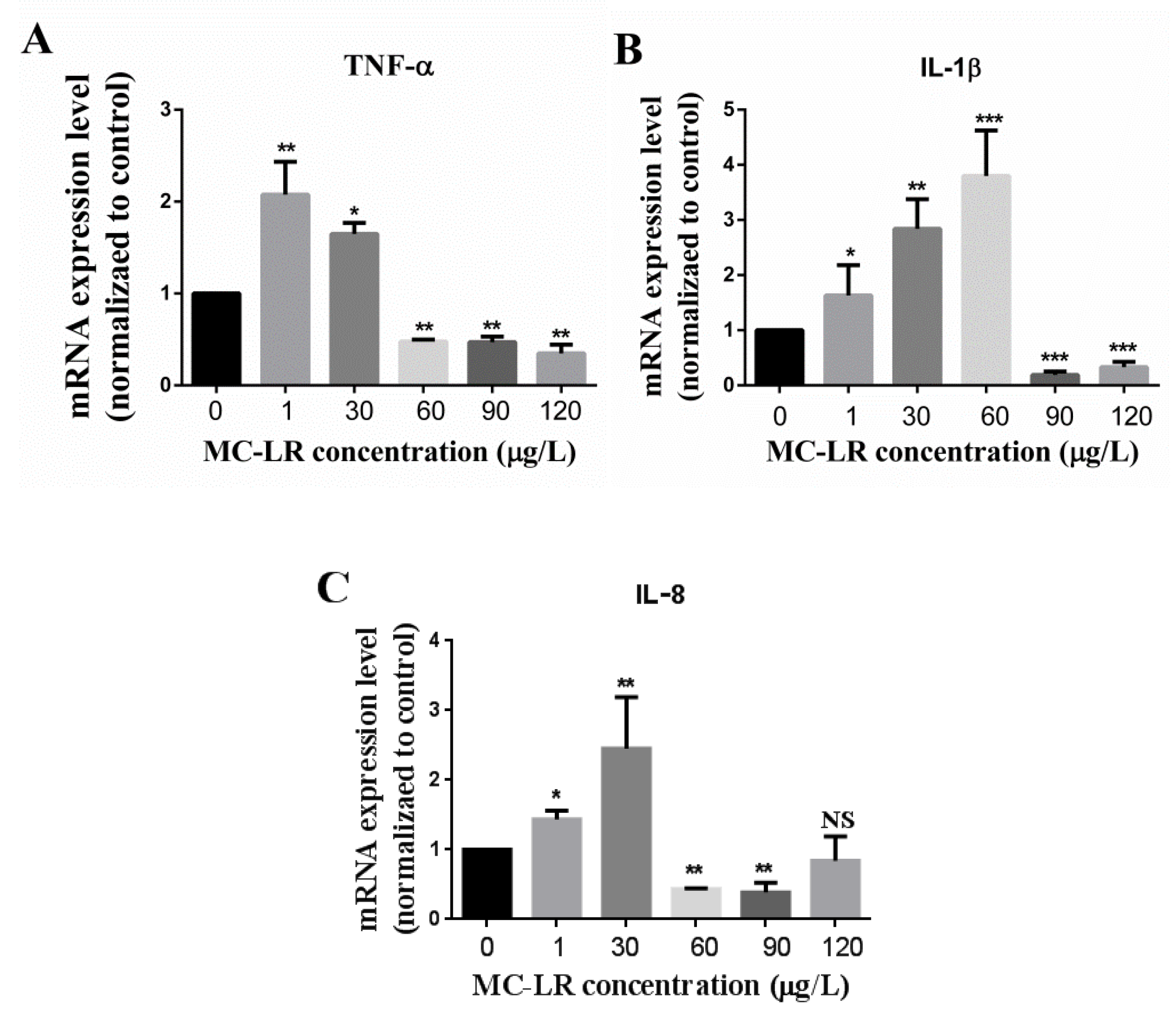
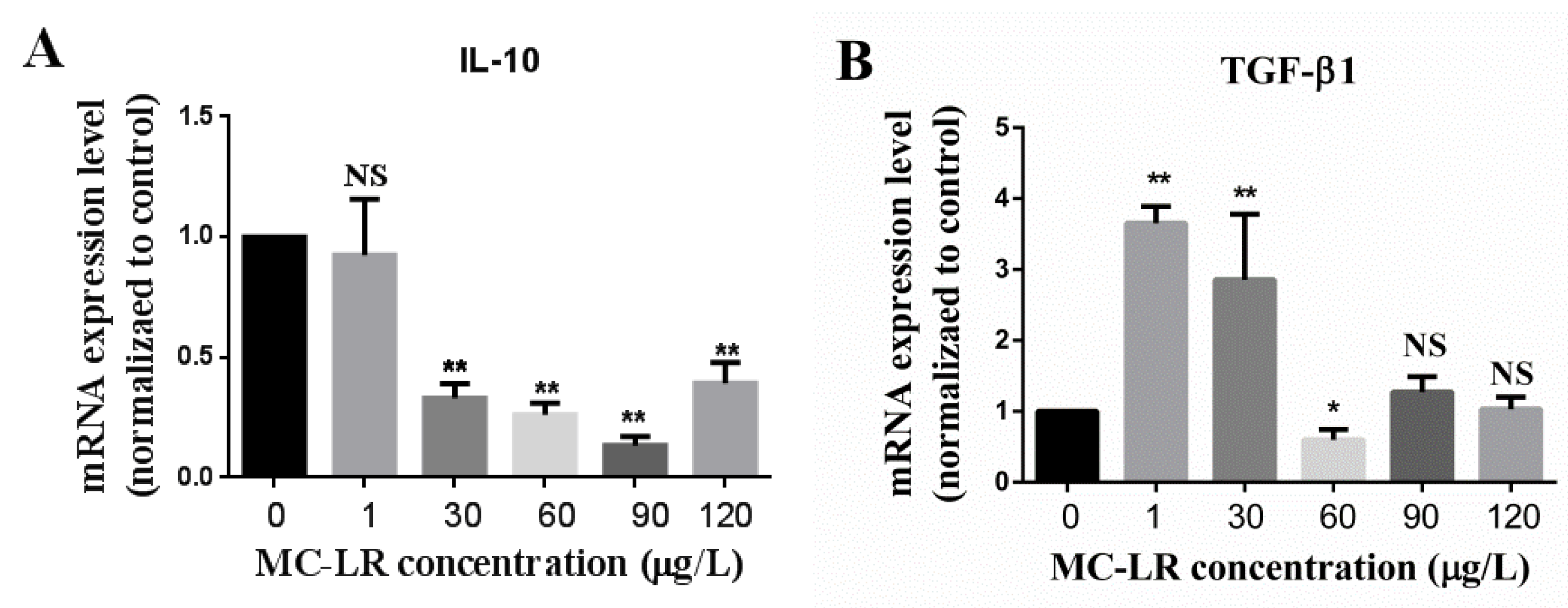
2.3. Effect of MC-LR on the Expression of Jejunum Inflammation-Related Factors in Mice
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Mice and Reagents
5.2. Tissue Anatomy
5.3. Hematoxylin and Eosin Staining
5.4. Quantitative RT-PCR
5.5. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Carmichael, W.W.; Azevedo, S.M.F.O.; An, J.S.; Molica, R.J.R.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human Fatalities from Cyanobacteria: Chemical and Biological Evidence for Cyanotoxins. Environ. Health. Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Chia, M.A.; Kramer, B.J.; Jankowiak, J.G.; Bittencourt-Oliveira, M.D.C.; Gobler, C.J. The Individual and Combined Effects of the Cyanotoxins, Anatoxin-a and Microcystin-LR, on the Growth, Toxin Production, and Nitrogen Fixation of Prokaryotic and Eukaryotic Algae. Toxins 2019, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wen, C.; Zheng, S.; Yang, S.; Chen, J.; Feng, X. Involvement of MAPK/ERK1/2 pathway in microcystin-induced microfilament reorganization in HL7702 hepatocytes. J. Toxicol. Environ. Health Part A 2018, 81, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Guo, J.; Huang, F.Y.; Massey, I.Y.; Huang, R.X.; Li, Y.H.; Wen, C.; Ding, P.; Zeng, W.M.; Liang, G.Y. Removal of Microcystin-LR by a Novel Native Effective Bacterial Community Designated as YFMCD4 Isolated from Lake Taihu. Toxin 2018, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Guo, H.; Wang, L.; Zhang, D.; Wu, X.; Li, L.; Li, D.; Tang, R. Nitrite Enhances MC-LR-Induced Changes on Splenic Oxidation Resistance and Innate Immunity in Male Zebrafish. Toxins 2018, 10, 512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhuang, H.; Yang, H.; Xue, W.; Wang, L.; Wei, W. Microcystin-LR disturbs testicular development of giant freshwater prawn Macrobrachium rosenbergii. Chemosphere 2019, 222, 584–592. [Google Scholar] [CrossRef]
- Spoof, L.; Catherine, A. Appendix 3. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 526–537. [Google Scholar]
- Chen, J.; Xie, P. Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and -RR in two freshwater shrimps, Palaemon modestus and Macrobrachium nipponensis, from a large shallow, eutrophic lake of the subtropical China. Toxicon 2005, 45, 615–625. [Google Scholar] [CrossRef]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Tezi, C.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef]
- Papadimitriou, T.; Kagalou, I.; Stalikas, C.; Pilidis, G.; Leonardos, I.D. Assessment of microcystin distribution and biomagnification in tissues of aquatic food web compartments from a shallow lake and evaluation of potential risks to public health. Ecotoxicology 2012, 21, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-X.; Huang, H.; Yang, L.; Zhang, X.-F.; Zhang, S.-S.; Liu, H.-H.; Wang, Y.-Q.; Yuan, L.; Cheng, X.-M.; Zhuang, D.-G.; et al. Gastrointestinal toxicity induced by microcystins. World J. Clin. Cases 2018, 6, 344–354. [Google Scholar] [CrossRef]
- Chen, L.; Yang, S.; Wen, C.; Zheng, S.; Yang, Y.; Feng, X.; Chen, J.; Luo, D.; Liu, R.; Yang, F. Regulation of Microcystin-LR-Induced DNA Damage by miR-451a in HL7702 Cells. Toxins 2019, 11, 164. [Google Scholar] [CrossRef] [PubMed]
- Mrdjen, I.; Morse, M.A.; Ruch, R.J.; Knobloch, T.J.; Choudhary, S.; Weghorst, C.M.; Lee, J. Impact of Microcystin-LR on Liver Function Varies by Dose and Sex in Mice. Toxins 2018, 10, 435. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, S.; Liu, C.; Wu, J.; Wang, Y.; Yuan, L.; Du, X.; Wang, R.; Marwa, P.W.; Zhuang, D.; et al. Resveratrol Ameliorates Microcystin-LR-Induced Testis Germ Cell Apoptosis in Rats via SIRT1 Signaling Pathway Activation. Toxins 2018, 10, 235. [Google Scholar] [CrossRef]
- Vidal, F.; Sedan, D.; D’Agostino, D.; Cavalieri, M.L.; Mullen, E.; Varela, M.M.P.; Flores, C.; Caixach, J.; Andrinolo, D. Recreational Exposure during Algal Bloom in Carrasco Beach, Uruguay: A Liver Failure Case Report. Toxins 2017, 9, 267. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, S.M.; Carmichael, W.W.; Jochimsen, E.M.; Rinehart, K.L.; Lau, S.; Shaw, G.R.; Eaglesham, G.K. Human intoxication by microcystins during renal dialysis treatment in Caruaru—Brazil. Toxicology 2002, 181, 441–446. [Google Scholar] [CrossRef]
- WHO. Cyanobacterial Toxins: Microcystin-LR. Guidelines for Drinking Water Quality; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Thomson, A.B.; Drozdowski, L.; Iordache, C.; Thomson, B.K.; Vermeire, S.; Clandinin, M.T.; Wild, G. Small bowel review: Normal physiology, part 1. Dig. Dis. Sci. 2003, 48, 1546–1564. [Google Scholar] [CrossRef]
- Middendorp, S.; Schneeberger, K.; Wiegerinck, C.L.; Mokrý, M.; Akkerman, R.D.L.; Van Wijngaarden, S.; Clevers, H.; Nieuwenhuis, E.E.S. Adult Stem Cells in the Small Intestine Are Intrinsically Programmed with Their Location-Specific Function. Stem Cells 2014, 32, 1083–1091. [Google Scholar] [CrossRef]
- McKie, A.T.; Barrow, D.; Latunde-Dada, G.O.; Rolfs, A.; Sager, G.; Mudaly, E.; Mudaly, M.; Richardson, C.; Barlow, D.; Bomford, A.; et al. An Iron-Regulated Ferric Reductase Associated with the Absorption of Dietary Iron. Science 2001, 291, 1755–1759. [Google Scholar] [CrossRef] [PubMed]
- Shneider, B.L. Intestinal Bile Acid Transport: Biology, Physiology, and Pathophysiology. J. Pediatr. Gastroenterol. Nutr. 2001, 32, 407–417. [Google Scholar] [CrossRef]
- Kogut, M.H.; Genovese, K.J.; Swaggerty, C.L.; He, H.; Broom, L. Inflammatory phenotypes in the intestine of poultry: Not all inflammation is created equal. Poult. Sci. 2018, 97, 2339–2346. [Google Scholar] [CrossRef]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef]
- Rigby, R.J.; Hunt, M.R.; Scull, B.P.; Simmons, J.G.; Speck, K.E.; Helmrath, M.A.; Lund, P.K. A new animal model of post-surgical bowel inflammation and fibrosis: The effect of commensal microflora. Gut 2009, 58, 1104–1112. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C.; Wang, T.; Hong, S.; Peddie, S.; Crampe, M.; Laing, K.; Cunningham, C.; Zou, J.; Laing, K. Cytokines and innate immunity of fish. Dev. Comp. Immunol. 2001, 25, 713–723. [Google Scholar] [CrossRef]
- Tran, H.B.; Chen, S.-C.; Chaung, H.-C.; Cheng, T.-C. Molecular cloning of IL-6, IL-10, IL-11, IFN-ɤ and modulation of pro- and anti-inflammatory cytokines in cobia (Rachycentron canadum) after Photobacterium damselae subsp. piscicida infection. Comp. Biochem. Physiol. Part B Biochem. Mol. Boil. 2019, 230, 10–18. [Google Scholar] [CrossRef]
- Weel, J.F.; Hopman, C.T.; Van Putten, J.P. Stable expression of lipooligosaccharide antigens during attachment, internalization, and intracellular processing of Neisseria gonorrhoeae in infected epithelial cells. Infect. Immun. 1989, 57, 3395–3402. [Google Scholar] [PubMed]
- Brundin, L.; Bryleva, E.Y.; Thirtamara Rajamani, K. Role of Inflammation in Suicide: From Mechanisms to Treatment. Neuropsychopharmacology 2017, 42, 271–283. [Google Scholar] [CrossRef]
- Yoshie, O.; Imai, T.; Nomiyama, H. Chemokines in immunity. Adv. Immunol. 2001, 78, 57–110. [Google Scholar] [PubMed]
- Zuena, A.R.; Casolini, P.; Lattanzi, R.; Maftei, D. Chemokines in Alzheimer’s Disease: New Insights into Prokineticins, Chemokine-Like Proteins. Front. Pharmacol. 2019, 10, 622. [Google Scholar] [CrossRef]
- Liu, N.; Chen, R.; Du, H.; Wang, J.; Zhang, Y.; Wen, J. Expression of IL-10 and TNF-alpha in rats with cerebral infarction after transplantation with mesenchymal stem cells. Cell. Mol. Immunol. 2009, 6, 207–213. [Google Scholar] [CrossRef]
- Kim, S.K.; Barron, L.; Hinck, C.S.; Petrunak, E.M.; Cano, K.E.; Thangirala, A.; Iskra, B.; Brothers, M.; Vonberg, M.; Leal, B.; et al. An engineered transforming growth factor beta (TGF-beta) monomer that functions as a dominant negative to block TGF-beta signaling. J. Biol. Chem. 2017, 292, 7173–7188. [Google Scholar] [CrossRef]
- Alijotas-Reig, J.; Esteve-Valverde, E.; Ferrer-Oliveras, R.; Llurba, E.; Gris, J.M. Tumor Necrosis Factor-Alpha and Pregnancy: Focus on Biologics. An Updated and Comprehensive Review. Clin. Rev. Allergy Immunol. 2017, 53, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Sobczak, M.; Jozkowicz, A.; Dulak, J. TGF-β1/Smads and miR-21 in Renal Fibrosis and Inflammation. Mediat. Inflamm. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Fuss, I.J.; Blumberg, R.S. The immunology of mucosal models of inflammation. Annu. Rev. Immunol. 2002, 20, 495–549. [Google Scholar] [CrossRef]
- Mehaffey, E.; Majid, D.S.A. Tumor necrosis factor-α, kidney function, and hypertension. Am. J. Physiol. Physiol. 2017, 313, F1005–F1008. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P.; Li, L.; Xu, J. First Identification of the Hepatotoxic Microcystins in the Serum of a Chronically Exposed Human Population Together with Indication of Hepatocellular Damage. Toxicol. Sci. 2009, 108, 81–89. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.A.; Zhao, Q.; Pu, C.; Qiu, Z.; Zhang, R.; Shu, W. А сrоѕѕ-ѕесtіоnаl іnvеѕtіgаtіоn оf сhrоnіс ехр оѕurе tо mісrосуѕtіn іn relationship to childhood liver damage in the Three Gorges R eservoir Region, China. Environ. Health Perspect. 2011, 119, 1483–1488. [Google Scholar] [CrossRef]
- Sedan, D.; Laguens, M.; Copparoni, G.; Aranda, J.O.; Giannuzzi, L.; Marra, C.A.; Andrinolo, D. Hepatic and intestine alterations in mice after prolonged exposure to low oral doses of Microcystin-LR. Toxicon 2015, 104, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Su, R.C.; Blomquist, T.M.; Kleinhenz, A.L.; Khalaf, F.K.; Dube, P.; Lad, A.; Breidenbach, J.D.; Mohammed, C.J.; Zhang, S.G.; Baum, C.E.; et al. Exposure to the Harmful Algal Bloom (HAB) Toxin Microcystin-LR (MC-LR) Prolongs and Increases Severity of Dextran Sulfate Sodium (DSS)-Induced Colitis. Toxins 2019, 11, 371. [Google Scholar] [CrossRef]
- Pan, C.; Chen, Y.; Xu, T.; Wang, J.; Li, D.; Han, X. Chronic exposure to microcystin-leucine-arginine promoted proliferation of prostate epithelial cells resulting in benign prostatic hyperplasia. Environ. Pollut. 2018, 242, 1535–1545. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Dignass, A.U. Intestinal barrier function. Curr. Opin. Clin. Nutr. 2002, 5, 685–694. [Google Scholar] [CrossRef]
- Ren, Z.; Guo, C.; Yu, S.; Zhu, L.; Wang, Y.; Hu, H.; Deng, J. Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function. Int. J. Mol. Sci. 2019, 20, 2777. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Ito, E.; Kondo, F.; Terao, K.; Harada, K.-I. Neoplastic nodular formation in mouse liver induced by repeated intraperitoneal injections of microcystin-LR. Toxicon 1997, 35, 1453–1457. [Google Scholar] [CrossRef]
- Ito, E.; Kondo, F.; Harada, K.-I. First report on the distribution of orally administered microcystin-LR in mouse tissue using an immunostaining method. Toxicon 2000, 38, 37–48. [Google Scholar] [CrossRef]
- Chen, C.; Liu, W.; Wang, L.; Li, J.; Chen, Y.; Jin, J.; Kawan, A.; Zhang, X. Pathological damage and immunomodulatory effects of zebrafish exposed to microcystin-LR. Toxicon 2016, 118, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, B.; Wu, H.; Nie, P. Effects of pure microcystin-LR on the transcription of immune related genes and heat shock proteins in larval stage of zebrafish (Danio rerio). Aquaculture 2009, 289, 154–160. [Google Scholar] [CrossRef]
- Rymuszka, A.; Sierosławska, A.; Bownik, A.; Skowroński, T. Microcystin-LR modulates selected immune parameters and induces necrosis/apoptosis of carp leucocytes. Environ. Toxicol. Chem. 2010, 29, 569–574. [Google Scholar] [CrossRef]
- Xia, H.; Song, T.; Wang, L.; Jiang, L.; Zhou, Q.; Wang, W.; Liu, L.; Yang, P.; Zhang, X. Effects of dietary toxic cyanobacteria and ammonia exposure on immune function of blunt snout bream (Megalabrama amblycephala). Fish Shellfish. Immunol. 2018, 78, 383–391. [Google Scholar] [CrossRef]
- Fenton, M.J.; Vermeulen, M.W.; Clark, B.D.; Webb, A.C.; Auron, P.E. Human pro-IL-1 beta gene expression in monocytic cells is regulated by two distinct pathways. J. Immunol. 1988, 140, 2267–2273. [Google Scholar]
- Jarrous, N.; Kaempfer, R. Induction of human interleukin-1 gene expression by retinoic acid and its regulation at processing of precursor transcripts. J. Boil. Chem. 1994, 269, 23141–23149. [Google Scholar]
- Zheng, J.; Wang, J.; Pouliot, M.; Authier, S.; Zhou, D.; Loose, D.S.; Hauer-Jensen, M. Gene expression profiling in non-human primate jejunum, ileum and colon after total-body irradiation: A comparative study of segment-specific molecular and cellular responses. BMC Genom. 2015, 16, 23. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H.W.; Seidler, S.; Gassler, N.; Nattermann, J.; Luedde, T.; Trautwein, C.; Tacke, F. Interleukin-8 Is Activated in Patients with Chronic Liver Diseases and Associated with Hepatic Macrophage Accumulation in Human Liver Fibrosis. PLoS ONE 2011, 6, e21381. [Google Scholar] [CrossRef]
- Freire-De-Lima, C.G.; Xiao, Y.Q.; Gardai, S.J.; Bratton, D.L.; Schiemann, W.P.; Henson, P.M. Apoptotic Cells, through Transforming Growth Factor-β, Coordinately Induce Anti-inflammatory and Suppress Pro-inflammatory Eicosanoid and NO Synthesis in Murine Macrophages. J. Boil. Chem. 2006, 281, 38376–38384. [Google Scholar] [CrossRef] [PubMed]
- Savill, J.; Fadok, V. Corpse clearance defines the meaning of cell death. Nature 2000, 407, 784–788. [Google Scholar] [CrossRef]
- Voll, R.E.; Herrmann, M.; Roth, E.A.; Stach, C.; Kalden, J.R.; Girkontaite, I. Immunosuppressive effects of apoptotic cells. Nature 1997, 390, 350–351. [Google Scholar] [CrossRef] [PubMed]
- Savill, J.; Dransfield, I.; Gregory, C.; Haslett, C. A blast from the past: Clearance of apoptotic cells regulates immune responses. Nat. Rev. Immunol. 2002, 2, 965–975. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, C.; Chen, Y.; Chen, Y. TGF-β in Inflammatory Bowel Diseases: A Tale of the Janus-Like Cytokine. Crit. Rev. Eukaryot. Gene Expr. 2015, 25, 335–347. [Google Scholar] [CrossRef]
- Ruemmele, F.M.; Garnier-Lengliné, H. Transforming Growth Factor and Intestinal Inflammation: The Role of Nutrition. Nestle Nutr. Inst. Workshop Ser. 2013, 77, 91–98. [Google Scholar] [PubMed]
- Huang, X.R.; Chung, A.C.; Zhou, K.L.; Wang, X.J.; Lan, H.Y. Latent TGF- 𝛽1 protects against crescentic glomerulonephritis. J. Am. Soc. Nephrol. 2008, 19, 233–242. [Google Scholar] [CrossRef]
- Dai, C.; Wen, X.; He, W.; Liu, Y. Inhibition of proinflammatory RANTES expression by TGF-𝛽1 is mediated by glycogen synthase kinase-3𝛽-dependent 𝛽-catenin signaling. J. Biol. Chem. 2011, 286, 7052–7059. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, L.; Ma, R.; Mu, Q.; Yu, N.; Zhang, Y.; Tang, Y.; Li, Y.; Jiang, G.; Zhao, D.; et al. JiangTang XiaoKe granule attenuates cathepsin K expression and improves IGF-1 expression in the bone of high fat diet induced KK-Ay diabetic mice. Life Sci. 2016, 148, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Chang, H.; Ji, W.; Peng, H.; Shi, F.; Bai, Y.B.; Yin, J.S. Comparative study on different methods of detecting eosinophils in nasal polyps. Lin Chung Er Bi Yan Hou Tou Jing Wai Ke Za Zhi 2018, 32, 745–748. (In Chinese) [Google Scholar] [PubMed]
- Wen, C.; Yang, S.; Zheng, S.; Feng, X.; Chen, J.; Yang, F. Analysis of long non-coding RNA profiled following MC-LR-induced hepatotoxicity using high-throughput sequencing. J. Toxicol. Env. Heal. A. 2018, 22, 1165–1172. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Groups | Weight Gains (g) | Intestinal Length / mice heights (cm) | Water Consumption (mL) |
|---|---|---|---|
| Control (0 μg/L) | 7.24 ± 0.89 | 4.34 ± 0.22 | 4074.5 ± 126.50 |
| 1 μg/L | 7.66 ± 0.51 | 4.23 ± 0.16 | 3651.5 ± 202.07 |
| 30 μg/L | 6.32 ± 0.48 | 4.05 ± 0.20 | 4178 ± 99.70 |
| 60 μg/L | 6.65 ± 0.43 | 4.30 ± 0.22 | 3732 ± 166.45 |
| 90 μg/L | 5.91 ± 0.70 | 4.20 ± 0.15 | 4110 ± 107.38 |
| 120 μg/L | 6.03 ± 0.69 | 4.33 ± 0.17 | 4008 ± 77.92 |
| Primer | Sequence (5’→3’) | Annealing temperature (°C) | Product length (bp) |
|---|---|---|---|
| TNF-α | Forward: GTGCCTATGTCTCAGCCTCT Reverse: AGGCTTGTCACTCGAATTTTGA | 60 | 187 |
| IL-8 | Forward: TGGGTGAAGGCTACTGTTGG Reverse: AGGCTTTTCATGCTCAACACT | 60 | 114 |
| IL-10 | Forward: ATAACTGCACCCACTTCCCA Reverse: GGGCATCACTTCTACCAGGT | 60 | 206 |
| TGF-β1 | Forward: AGGGCTACCATGCCAACTTC Reverse: CCACGTAGTAGACGATGGC | 60 | 168 |
| IL-1β | Forward: AAGGGCTGCTTCCAAACCTTTGAC Reverse: ATACTGCCTGCCTGAAGCTCTTGT | 60 | 100 |
| β-actin | Forward: CTAAGGCCAACCGTGAAAAG Reverse: ACCAGAGGCATACAGGGACA | 60 | 104 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, L.; Huang, F.; Massey, I.Y.; Wen, C.; Zheng, S.; Xu, S.; Yang, F. Effects of Microcystin-LR on the Microstructure and Inflammation-Related Factors of Jejunum in Mice. Toxins 2019, 11, 482. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090482
Cao L, Huang F, Massey IY, Wen C, Zheng S, Xu S, Yang F. Effects of Microcystin-LR on the Microstructure and Inflammation-Related Factors of Jejunum in Mice. Toxins. 2019; 11(9):482. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090482
Chicago/Turabian StyleCao, Linghui, Feiyu Huang, Isaac Yaw Massey, Cong Wen, Shuilin Zheng, Shuaishuai Xu, and Fei Yang. 2019. "Effects of Microcystin-LR on the Microstructure and Inflammation-Related Factors of Jejunum in Mice" Toxins 11, no. 9: 482. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090482