Morphology and Phylogenetics of Benthic Prorocentrum Species (Dinophyceae) from Tropical Northwestern Australia

 ,
,
Abstract
:1. Introduction
2. Results
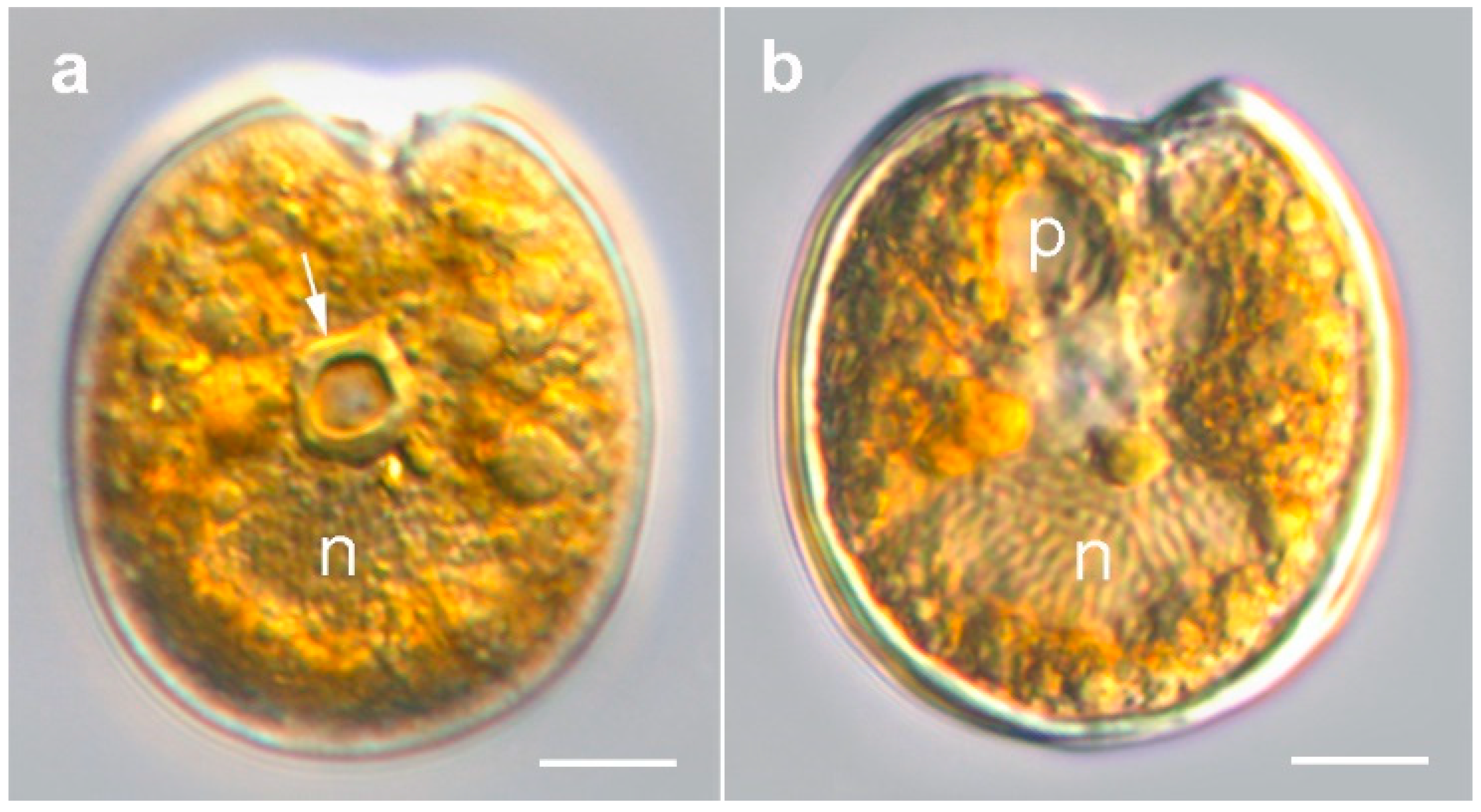
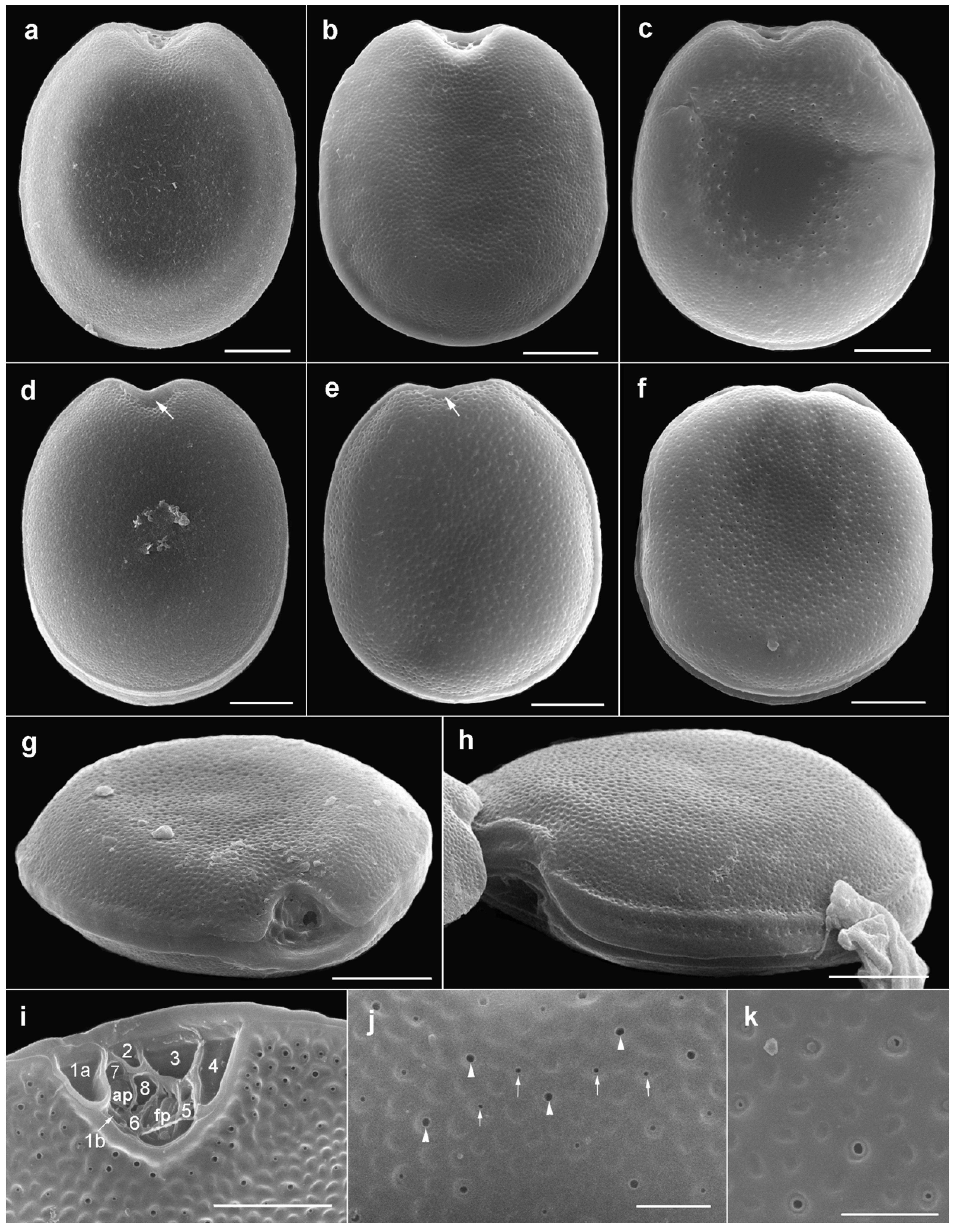
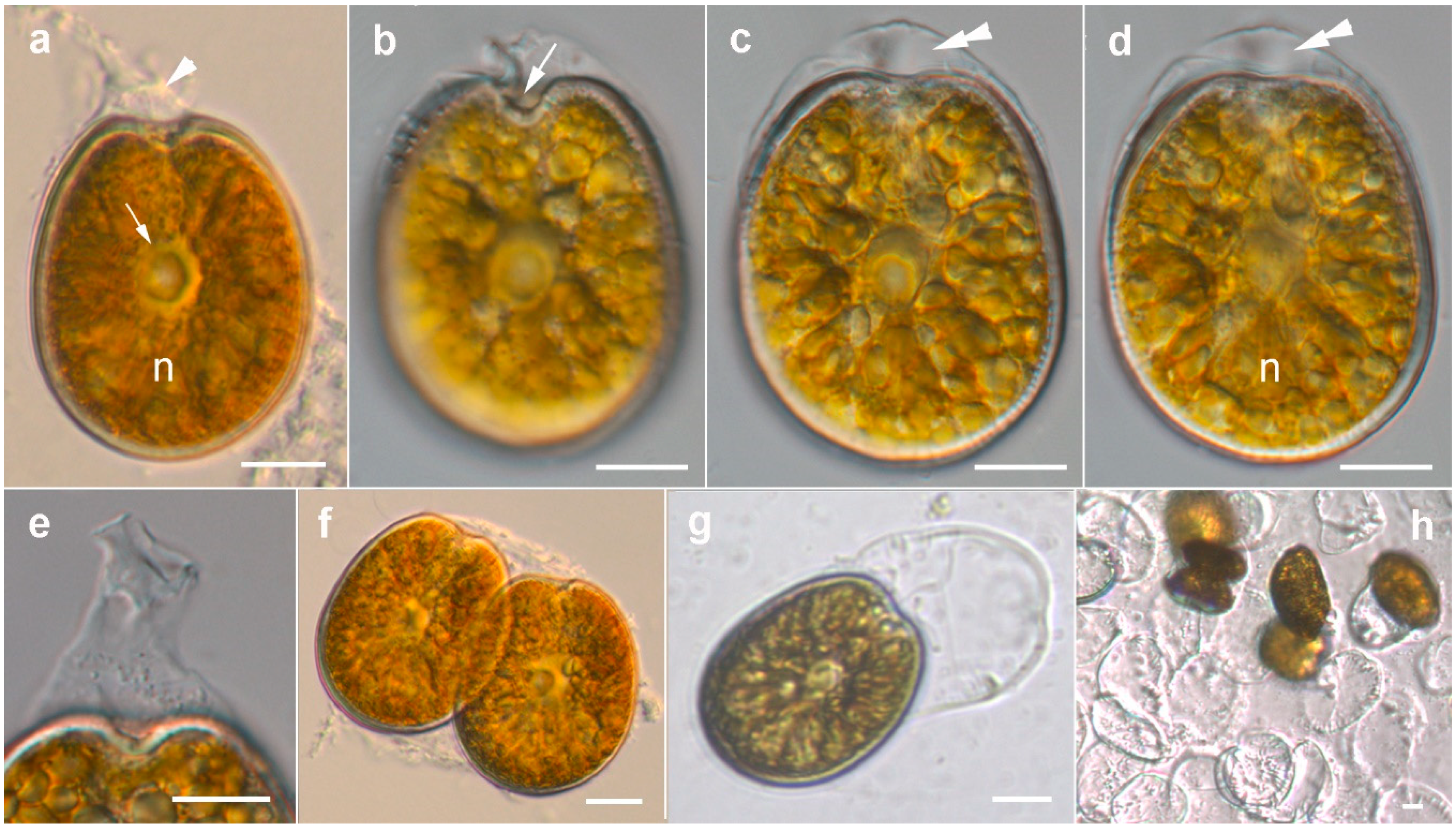
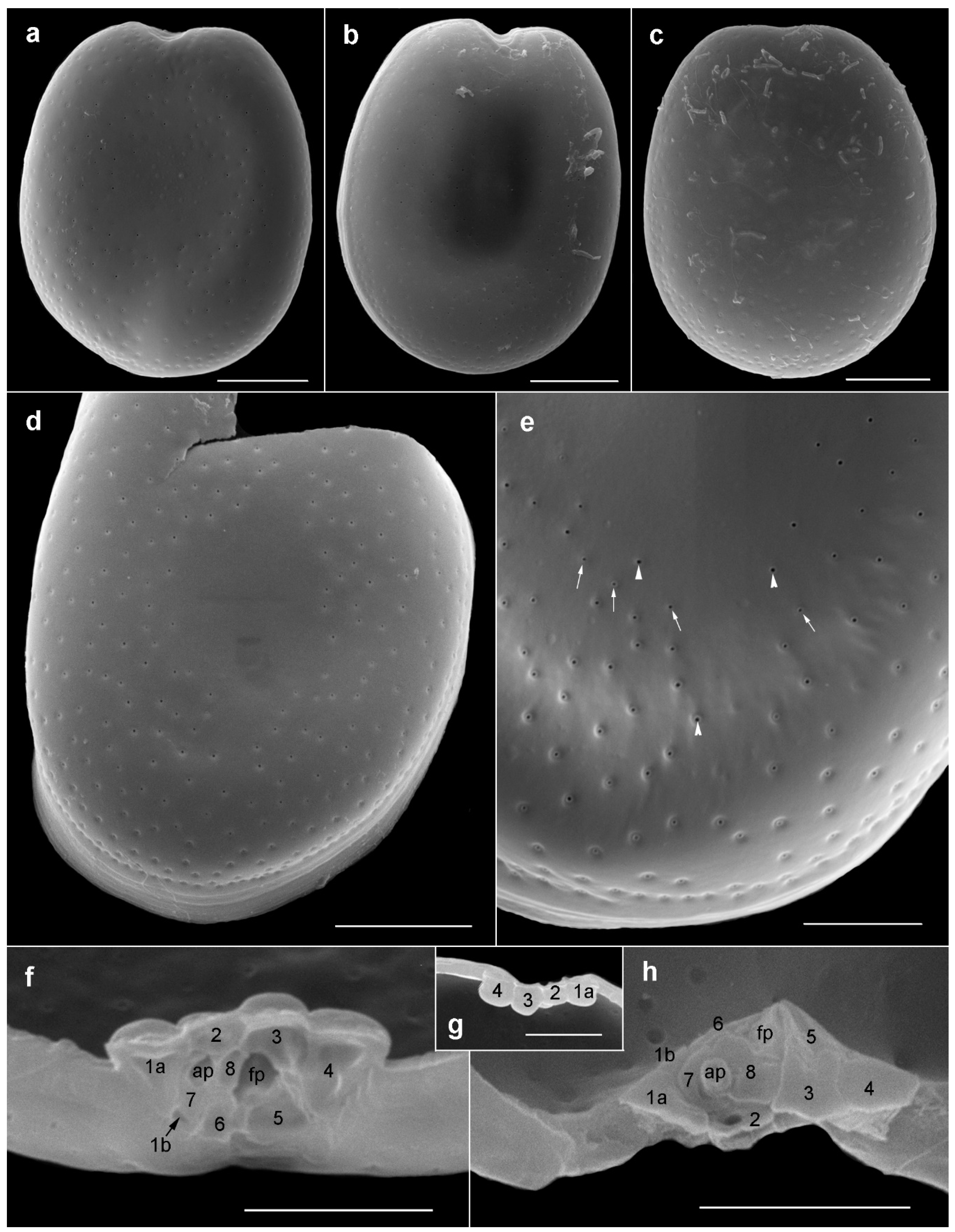
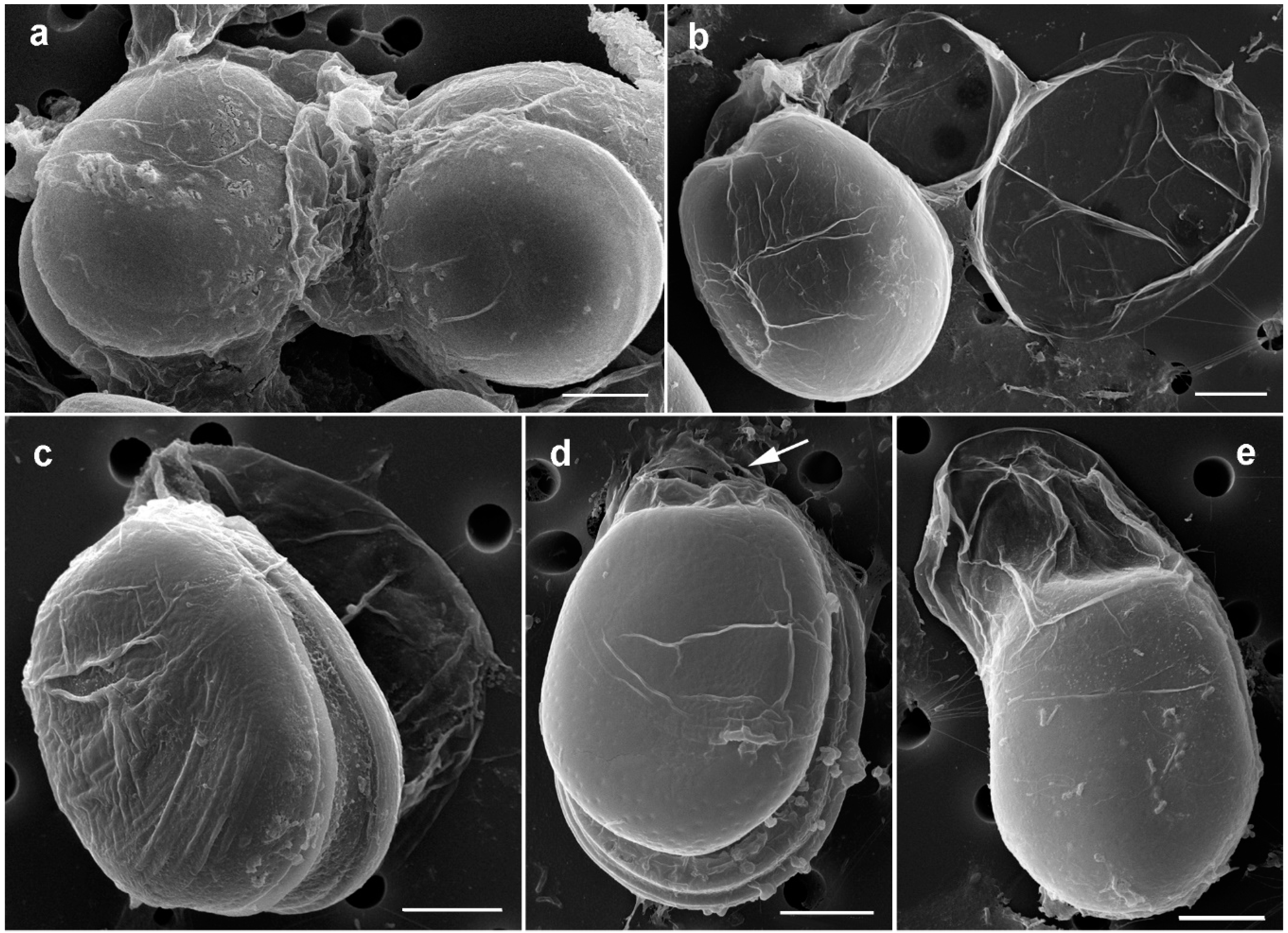
2.1. Morphology of Prorocentrum Species
2.1.1. Prorocentrum Concavum
2.1.2. Prorocentrum Malayense
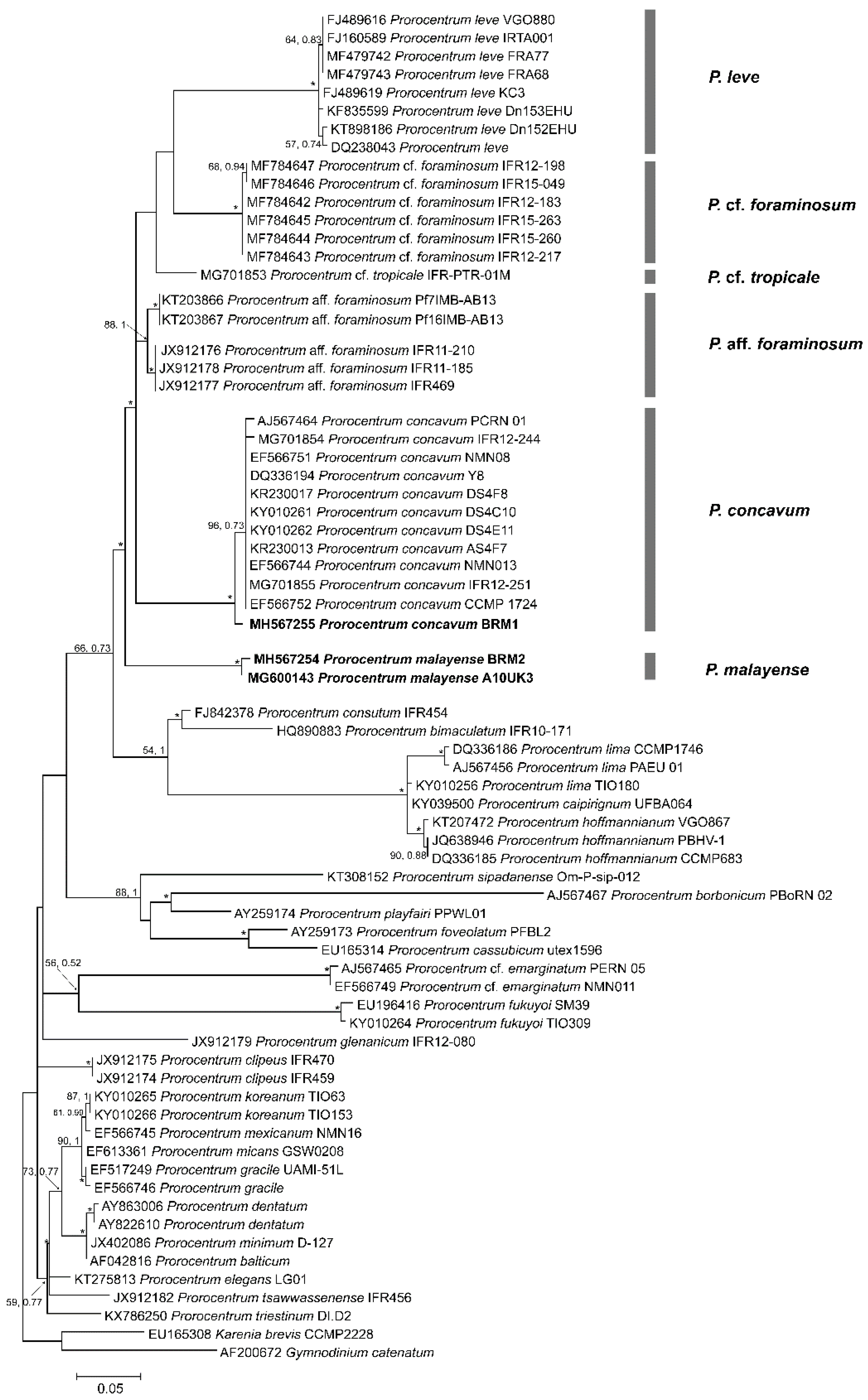
2.2. Phylogenetics
2.3. Toxin Analyses
3. Discussion
3.1. Morphological Study
3.2. Phylogenetic Analysis
3.3. Toxicity
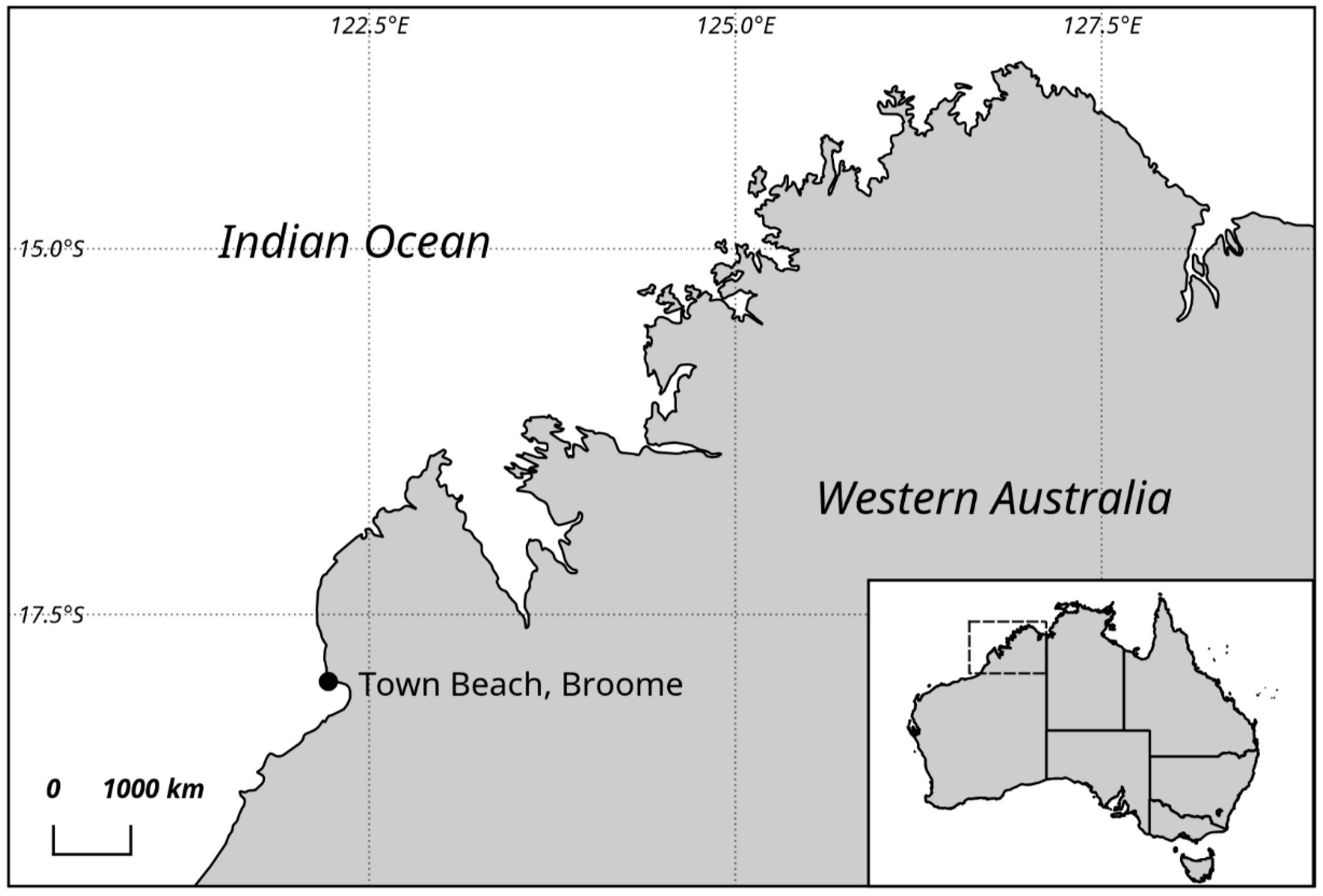
4. Materials and Methods
4.1. Sampling and Culture Establishment
4.2. Microscopy
4.3. DNA Extraction and PCR Amplification
4.4. Phylogenetic Analysis
4.5. Determination of Okadaic Acid (OA) and Dinophysistoxins (DTXs)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahé, F.; Logares, R.; Lara, E.; Berney, C.; Le Bescot, N.; Probert, I. Eukaryotic plankton diversity in the sunlit ocean. Science 2015, 348. [Google Scholar] [CrossRef]
- Massana, R.; Gobet, A.; Audic, S.; Bass, D.; Bittner, L.; Boutte, C.; Chambouvet, A.; Christen, R.; Claverie, J.M.; Decelle, J. Marine protist diversity in European coastal waters and sediments as revealed by high throughput sequencing. Environ. Microbiol. 2015, 17, 4035–4049. [Google Scholar] [CrossRef]
- Le Bescot, N.; Mahé, F.; Audic, S.; Dimier, C.; Garet, M.J.; Poulain, J.; Wincker, P.; de Vargas, C.; Siano, R. Global patterns of pelagic dinoflagellate diversity across protist size classes unveiled by metabarcoding. Environ. Microbiol. 2016, 18, 609–626. [Google Scholar] [CrossRef]
- Murray, S. Diversity and Phylogenetics of Sand-Dwelling Dino-Flagellates; VDM Verlag Dr. Müller Saarbrücken: Riga, Latvia, 2009. [Google Scholar]
- Hoppenrath, M.; Murray, S.A.; Chomérat, N.; Horiguchi, T. Marine Benthic Dinoflagellates-Unveiling Their Worldwide Biodiversity; Schweizerbart/Borntraeger: Stuttgart, Germany, 2014. [Google Scholar]
- Kohli, G.S.; Neilan, B.A.; Brown, M.V.; Hoppenrath, M.; Murray, S.A. Cob gene pyrosequencing enables characterization of benthic dinoflagellate diversity and biogeography. Environ. Microbiol. 2014, 16, 467–485. [Google Scholar] [CrossRef]
- Smith, K.F.; Kohli, G.S.; Murray, S.A.; Rhodes, L.L. Assessment of the metabarcoding approach for community analysis of benthic-epiphytic dinoflagellates using mock communities. N. Z. J. Mar. Freshw. Res. 2017, 51, 555–576. [Google Scholar] [CrossRef]
- Yasumoto, T.; Seino, N.; Murakami, Y.; Murata, M. Toxins produced by benthic dinoflagellates. Biol. Bull. 1987, 172, 128–131. [Google Scholar] [CrossRef]
- Yasumoto, T.; Murata, M. Marine toxins. Chem. Rev. 1993, 93, 1897–1909. [Google Scholar] [CrossRef]
- Kantiani, L.; Llorca, M.; Sanchís, J.; Farré, M.; Barceló, D. Emerging food contaminants: A review. Anal. Bioanal. Chem. 2010, 398, 2413–2427. [Google Scholar] [CrossRef]
- Wang, D.-Z. Neurotoxins from marine dinoflagellates: A brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Chomérat, N.; Horiguchi, T.; Schweikert, M.; Nagahama, Y.; Murray, S. Taxonomy and phylogeny of the benthic Prorocentrum species (Dinophyceae)—A proposal and review. Harmful Algae 2013, 27, 1–28. [Google Scholar] [CrossRef]
- Chomérat, N.; Bilien, G.; Zentz, F. A taxonomical study of benthic Prorocentrum species (Prorocentrales, Dinophyceae) from Anse Dufour (Martinique Island, eastern Caribbean Sea). Mar. Biodivers. 2019, 49, 1299–1319. [Google Scholar] [CrossRef]
- Lim, Z.F.; Luo, Z.; Lee, L.K.; Hii, K.S.; Teng, S.T.; Chan, L.L.; Chomérat, N.; Krock, B.; Gu, H.; Lim, P.T. Taxonomy and toxicity of Prorocentrum from Perhentian Islands (Malaysia), with a description of a non-toxigenic species Prorocentrum malayense sp. nov. (Dinophyceae). Harmful Algae 2019, 83, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.; Nagahama, Y.; Fukuyo, Y. Phylogenetic study of benthic, spine-bearing prorocentroids, including Prorocentrum fukuyoi sp. nov. Phycol. Res. 2007, 55, 91–102. [Google Scholar] [CrossRef]
- Herrera-Sepúlveda, A.; Medlin, L.K.; Murugan, G.; Sierra-Beltrán, A.P.; Cruz-Villacorta, A.A.; Hernández-Saavedra, N.Y. Are Prorocentrum hoffmannianum and Prorocentrum belizeanum (Dinophyceae, Prorocentrales), the same species? An integration of morphological and molecular data. J. Phycol. 2015, 51, 173–188. [Google Scholar] [CrossRef]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal revision of the Alexandrium tamarense species complex (Dinophyceae) taxonomy: The introduction of five species with emphasis on molecular-based (rDNA) classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Hoppenrath, M.; Dorantes-Aranda, J.J.; Harwood, D.T.; Murray, S.A. Molecular and phylogenetic characterization of Ostreopsis (Dinophyceae) and the description of a new species, Ostreopsis rhodesae sp. nov., from a subtropical Australian lagoon. Harmful Algae 2016, 60, 116–130. [Google Scholar] [CrossRef]
- Mohammad-Noor, N.; Moestrup, Ø.; Daugbjerg, N. Light, electron microscopy and DNA sequences of the dinoflagellate Prorocentrum concavum (syn. P. arabianum) with special emphasis on the periflagellar area. Phycologia 2007, 46, 549–564. [Google Scholar] [CrossRef]
- Leaw, C.P.; Tan, T.H.; Lim, H.C.; Teng, S.T.; Yong, H.L.; Smith, K.F.; Rhodes, L.; Wolf, M.; Holland, W.C.; Vandersea, M.W.; et al. New scenario for speciation in the benthic dinoflagellate genus Coolia (Dinophyceae). Harmful Algae 2016, 55, 137–149. [Google Scholar] [CrossRef]
- Gómez, F.; Qiu, D.; Lin, S. The synonymy of the toxic dinoflagellates Prorocentrum mexicanum and P. rhathymum and the description of P. steidingerae sp. nov.(Prorocentrales, Dinophyceae). J. Eukaryot. Microbiol. 2017, 64, 668–677. [Google Scholar] [CrossRef]
- Nagahama, Y.; Fukuyo, Y. Redescription of Cryptomonas lima, collected from Sorrento, Italy, the basionym of Prorocentrum lima. Plankton Biol. Ecol. 2005, 52, 100–106. [Google Scholar]
- Murray, S.; Jørgensen, M.F.; Ho, S.Y.; Patterson, D.J.; Jermiin, L.S. Improving the analysis of dinoflagellate phylogeny based on rDNA. Protist 2005, 156, 269–286. [Google Scholar] [CrossRef]
- Murakami, Y.; Oshima, Y.; Yasumoto, T. Identification of okadaic acid as a toxic component of a marine dinoflagellate Prorocentrum lima. Bull. Jpn. Soc. Sci. Fish. 1982, 48, 69–72. [Google Scholar] [CrossRef]
- Dickey, R.W.; Bobzin, S.C.; Faulkner, D.J.; Bencsath, F.A.; Andrzejewski, D. Identification of okadaic acid from a Caribbean dinoflagellate, Prorocentrum concavum. Toxicon 1990, 28, 371–377. [Google Scholar] [CrossRef]
- Ten-Hage, L.; Turquet, J.; Quod, J.-P.; Puiseux-Dao, S.; Coute, A. Prorocentrum borbonicum sp. nov.(Dinophyceae), a new toxic benthic dinoflagellate from the southwestern Indian Ocean. Phycologia 2000, 39, 296–301. [Google Scholar] [CrossRef]
- Nakajima, I.; Oshima, Y.; Yasumoto, T. Toxicity of benthic dinoflagellates in Okinawa. Bull. Jpn. Soc. Sci. Fish. 1981, 47, 1029–1033. [Google Scholar] [CrossRef]
- Zhou, J.; Fritz, L. Okadaic acid antibody localizes to chloroplasts in the DSP-toxin-producing dinoflagellates Prorocentrum lima and Prorocentrum maculosum. Phycologia 1994, 33, 455–461. [Google Scholar] [CrossRef]
- Grzebyk, D.; Denardou, A.; Berland, B.; Pouchus, Y. Evidence of a new toxin in the red-tide dinoflagellate Prorocentrum minimum. J. Plankton Res. 1997, 19, 1111–1124. [Google Scholar] [CrossRef]
- Bauder, A.G.; Cembella, A.D.; Bricelj, V.M.; Quilliam, M.A. Uptake and fate of diarrhetic shellfish poisoning toxins from the dinoflagellate Prorocentrum lima in the bay scallop Argopecten irradians. Mar. Ecol. Prog. Ser. 2001, 213, 39–52. [Google Scholar] [CrossRef]
- Gayoso, A.M.; Dover, S.; Morton, S.; Busman, M.; Moeller, P.; Fulco, V.K.; Maranda, L. Diarrhetic shellfish poisoning associated with Prorocentrum lima (Dinophyceae) in Patagonian gulfs (Argentina). J. Shellfish Res. 2002, 21, 461–463. [Google Scholar]
- Heredia-Tapia, A.; Arredondo-Vega, B.; Nuñez-Vázquez, E.; Yasumoto, T.; Yasuda, M.; Ochoa, J. Isolation of Prorocentrum lima (Syn. Exuviaella lima) and diarrhetic shellfish poisoning (DSP) risk assessment in the Gulf of California, Mexico. Toxicon 2002, 40, 1121–1127. [Google Scholar] [CrossRef]
- Cruz, P.G.; Daranas, A.H.; Fernández, J.J.; Souto, M.L.; Norte, M. DTX5c, a new OA sulphate ester derivative from cultures of Prorocentrum belizeanum. Toxicon 2006, 47, 920–924. [Google Scholar] [CrossRef] [PubMed]
- Faust, M.A.; Vandersea, M.W.; Kibler, S.R.; Tester, P.A.; Litaker, R.W. Prorocentrum levis, a new benthic species (dinophyceae) from a mangrove island, twin cays, Belize. J. Phycol. 2008, 44, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Flórez-Barrós, F.; Prado-Alvarez, M.; Méndez, J.; Fernández-Tajes, J. Evaluation of genotoxicity in gills and hemolymph of clam Ruditapes decussatus fed with the toxic dinoflagellate Prorocentrum lima. J. Toxicol. Environ. Health A 2011, 74, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, S.M.; Mendes, M.C.Q.; Menezes, M.; Rodríguez, F.; Alves-de-Souza, C.; Branco, S.; Riobó, P.; Franco, J.; Nunes, J.M.C.; Huk, M. Morphology and phylogeny of Prorocentrum caipirignum sp. nov. (Dinophyceae), a new tropical toxic benthic dinoflagellate. Harmful Algae 2017, 70, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Tubaro, A.; Florio, C.; Luxich, E.; Sosa, S.; Della Loggia, R.; Yasumoto, T. A protein phosphatase 2A inhibition assay for a fast and sensitive assessment of okadaic acid contamination in mussels. Toxicon 1996, 34, 743–752. [Google Scholar] [CrossRef]
- Vale, P.; Sampayo, M.A.M. First confirmation of human diarrhoeic poisonings by okadaic acid esters after ingestion of razor clams (Solen marginatus) and green crabs (Carcinus maenas) in Aveiro lagoon, Portugal and detection of okadaic acid esters in phytoplankton. Toxicon 2002, 40, 989–996. [Google Scholar] [CrossRef]
- Fukuyo, Y. Taxonomical study on benthic dinoflagellates collected in coral reefs. Bull. Jpn. Soc. Sci. Fish. 1981, 47, 967–978. [Google Scholar] [CrossRef]
- Faust, M.A. Morphologilca details of six benthic species of Prorocentrum (Pyrrophyta) from a mangrove island, Twin Cays, Belize, including two new species. J. Phycol. 1990, 26, 548–558. [Google Scholar] [CrossRef]
- Morton, S. Morphology and toxicology of Prorocentrum faustiae sp. nov., a toxic species of non-planktonic dinoflagellate from Heron Island, Australia. Bot. Mar. 1998, 41, 565–570. [Google Scholar] [CrossRef]
- Grzebyk, D.; Sako, Y.; Berland, B. Phylogenetic analysis of nine species of Prorocentrum (Dinophyceae) inferred from 18S ribosomal DNA sequences, morphological comparisons, and description of Prorocentrum panamensis, sp. nov. J. Phycol. 1998, 34, 1055–1068. [Google Scholar] [CrossRef]
- Morton, S.L.; Faust, M.A.; Fairey, E.A.; Moeller, P.D. Morphology and toxicology of Prorocentrum arabianum sp. nov. (Dinophyceae) a toxic planktonic dinoflagellate from the Gulf of Oman, Arabian Sea. Harmful Algae 2002, 1, 393–400. [Google Scholar] [CrossRef]
- Mohammad-Noor, N.; Daugbjerg, N.; Moestrup, O.; Anton, A. Marine epibenthic dinoflagellates from Malaysia—A study of live cultures and preserved samples based on light and scanning electron microscopy. Nord. J. Bot. 2005, 24, 629–690. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, H.; Krock, B.; Lu, S.; Yang, W.; Gu, H. Morphology, molecular phylogeny and okadaic acid production of epibenthic Prorocentrum (Dinophyceae) species from the northern South China Sea. Algal Res. 2017, 22, 14–30. [Google Scholar] [CrossRef]
- Faust, M. Three new benthic species of Prorocentrum (Dinophyceae) from Twin Cays, Belize: P. maculosum sp. nov., P. foraminosum sp. nov. and P. formosum sp. nov. Phycologia 1993, 32, 410–418. [Google Scholar] [CrossRef]
- David, H.; Laza-Martínez, A.; García-Etxebarria, K.; Riobó, P.; Orive, E. Characterization of Prorocentrum elegans and Prorocentrum levis (D inophyceae) from the southeastern Bay of Biscay by morphology and molecular phylogeny. J. Phycol. 2014, 50, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Nishimura, T.; Uehara, K.; Sakanari, H.; Tawong, W.; Hariganeya, N.; Smith, K.; Rhodes, L.; Yasumoto, T.; Taira, Y. Phylogeography of Ostreopsis along west Pacific coast, with special reference to a novel clade from Japan. PLoS ONE 2011, 6, e27983. [Google Scholar] [CrossRef] [PubMed]
- Selina, M.S.; Efimova, K.V.; Hoppenrath, M. Redefinition of the genus Thecadinium (Dinophyceae) using morphological and molecular data, and description of Thecadinium pseudokofoidii sp. nov. Phycologia 2019, 58, 36–50. [Google Scholar] [CrossRef]
- Richlen, M.L.; Morton, S.L.; Barber, P.H.; Lobel, P.S. Phylogeography, morphological variation and taxonomy of the toxic dinoflagellate Gambierdiscus toxicus (Dinophyceae). Harmful Algae 2008, 7, 614–629. [Google Scholar] [CrossRef]
- Penna, A.; Fraga, S.; Battocchi, C.; Casabianca, S.; Giacobbe, M.G.; Riobó, P.; Vernesi, C. A phylogeographical study of the toxic benthic dinoflagellate genus Ostreopsis Schmidt. J. Biogeogr. 2010, 37, 830–841. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Cen, J.; Wang, H.; Cui, L.; Dong, Y.; Lu, S. Morphotypes of Prorocentrum lima (Dinophyceae) from Hainan island, South China Sea: Morphological and molecular characterization. Phycologia 2015, 54, 503–516. [Google Scholar] [CrossRef]
- Nagahama, Y.; Murray, S.; Tomaru, A.; Fukuyo, Y. Species boundaries in the toxic dinoflagellate Prorocentrum lima (Dinophyceae, Prorocentrales), based on morphological and phylogenetic characerters. J. Phycol. 2011, 47, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Marr, J.; de Freitas, A.S.; Quilliam, M.A.; Walter, J.A.; Wright, J.L.; Pleasance, S. New diol esters isolated from cultures of the dinoflagellates Prorocentrum lima and Prorocentrum concavum. J. Nat. Prod. 1992, 55, 1631–1637. [Google Scholar] [CrossRef]
- Kameneva, P.; Efimova, K.; Rybin, V.; Orlova, T. Detection of dinophysistoxin-1 in clonal culture of marine dinoflagellate Prorocentrum foraminosum (Faust MA, 1993) from the Sea of Japan. Toxins 2015, 7, 3947–3959. [Google Scholar] [CrossRef] [PubMed]
- Selina, M. The morphology and seasonal dynamics of the potentially toxic microalga Prorocentrum foraminosum Faust 1993 (Dinophyta) in Peter the Great Bay, the Sea of Japan. Russ. J. Mar. Biol. 2017, 43, 196–201. [Google Scholar] [CrossRef]
- Ajani, P.; Brett, S.; Krogh, M.; Scanes, P.; Webster, G.; Armand, L. The risk of harmful algal blooms (HABs) in the oyster-growing estuaries of New South Wales, Australia. Environ. Monit. Assess. 2013, 185, 5295–5316. [Google Scholar] [CrossRef]
- Hoppenrath, M. Dinoflagellate taxonomy—A review and proposal of a revised classification. Mar. Biodivers. 2017, 47, 381–403. [Google Scholar] [CrossRef]
- Murray, S.; Patterson, D.J. Cabra matta, gen. nov., sp. nov., a new benthic, heterotrophic dinoflagellate. Eur. J. Phycol. 2004, 39, 229–234. [Google Scholar] [CrossRef]
- Uhlig, G. Eine einfache Methode zur Extraktion der vagilen, mesopsammalen Mikrofauna. Helgoland Wiss. Meer 1964, 11, 178. [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms: Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Bachvaroff, T.R.; Handy, S.M.; Delwiche, C.F.; Leander, B.S. Molecular phylogeny of ocelloid-bearing dinoflagellates (Warnowiaceae) as inferred from SSU and LSU rDNA sequences. BMC Evol. Biol. 2009, 9, 116. [Google Scholar] [CrossRef]
- Scholin, C.A.; Herzog, M.; Sogin, M.; Anderson, D.M. Identification of group-and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- Nunn, G.; Theisen, B.; Christensen, B.; Arctander, P. Simplicity-correlated size growth of the nuclear 28S ribosomal RNA D3 expansion segment in the crustacean order Isopoda. J. Mol. Evol. 1996, 42, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- McNabb, P.; Selwood, A.I.; Holland, P.T. Multiresidue method for determination of algal toxins in shellfish: Single-laboratory validation and interlaboratory study. J. AOAC Int. 2005, 88, 761–772. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | P. concavum | P. cf. foraminosum | P. aff. foraminosum | P. leve | P. cf. tropicale | P. malayense |
|---|---|---|---|---|---|---|
| P. concavum (n = 12) | 0.005 (0.002) | |||||
| P. cf. foraminosum (n = 6) | 0.158 (0.017) | 0.001 (0.001) | ||||
| P. aff. foraminosum (n = 5) | 0.119 (0.015) | 0.093 (0.013) | 0.013 (0.004) | |||
| P. leve (n = 8) | 0.181 (0.019) | 0.153 (0.018) | 0.138 (0.016) | 0.005 (0.002) | ||
| P. cf. tropicale (n = 1) | 0.132 (0.017) | 0.103 (0.015) | 0.084 (0.013) | 0.151 (0.017) | NA | |
| P. malayense (n = 2) | 0.163 (0.018) | 0.164 (0.018) | 0.129 (0.016) | 0.167 (0.018) | 0.159 (0.018) | 0.015 (0.004) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verma, A.; Kazandjian, A.; Sarowar, C.; Harwood, D.T.; Murray, J.S.; Pargmann, I.; Hoppenrath, M.; Murray, S.A. Morphology and Phylogenetics of Benthic Prorocentrum Species (Dinophyceae) from Tropical Northwestern Australia. Toxins 2019, 11, 571. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100571
Verma A, Kazandjian A, Sarowar C, Harwood DT, Murray JS, Pargmann I, Hoppenrath M, Murray SA. Morphology and Phylogenetics of Benthic Prorocentrum Species (Dinophyceae) from Tropical Northwestern Australia. Toxins. 2019; 11(10):571. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100571
Chicago/Turabian StyleVerma, Arjun, Aniuska Kazandjian, Chowdhury Sarowar, D. Tim Harwood, J. Sam Murray, Insa Pargmann, Mona Hoppenrath, and Shauna A. Murray. 2019. "Morphology and Phylogenetics of Benthic Prorocentrum Species (Dinophyceae) from Tropical Northwestern Australia" Toxins 11, no. 10: 571. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11100571