Red-Crowned Crane (Grus japonensis) Reproduction Was Improved by Inhibiting Mycotoxins with Montmorillonite in Feed


Abstract
:1. Introduction
2. Results
2.1. Dietary Mycotoxin Concentration
2.2. Reproductive Behavior
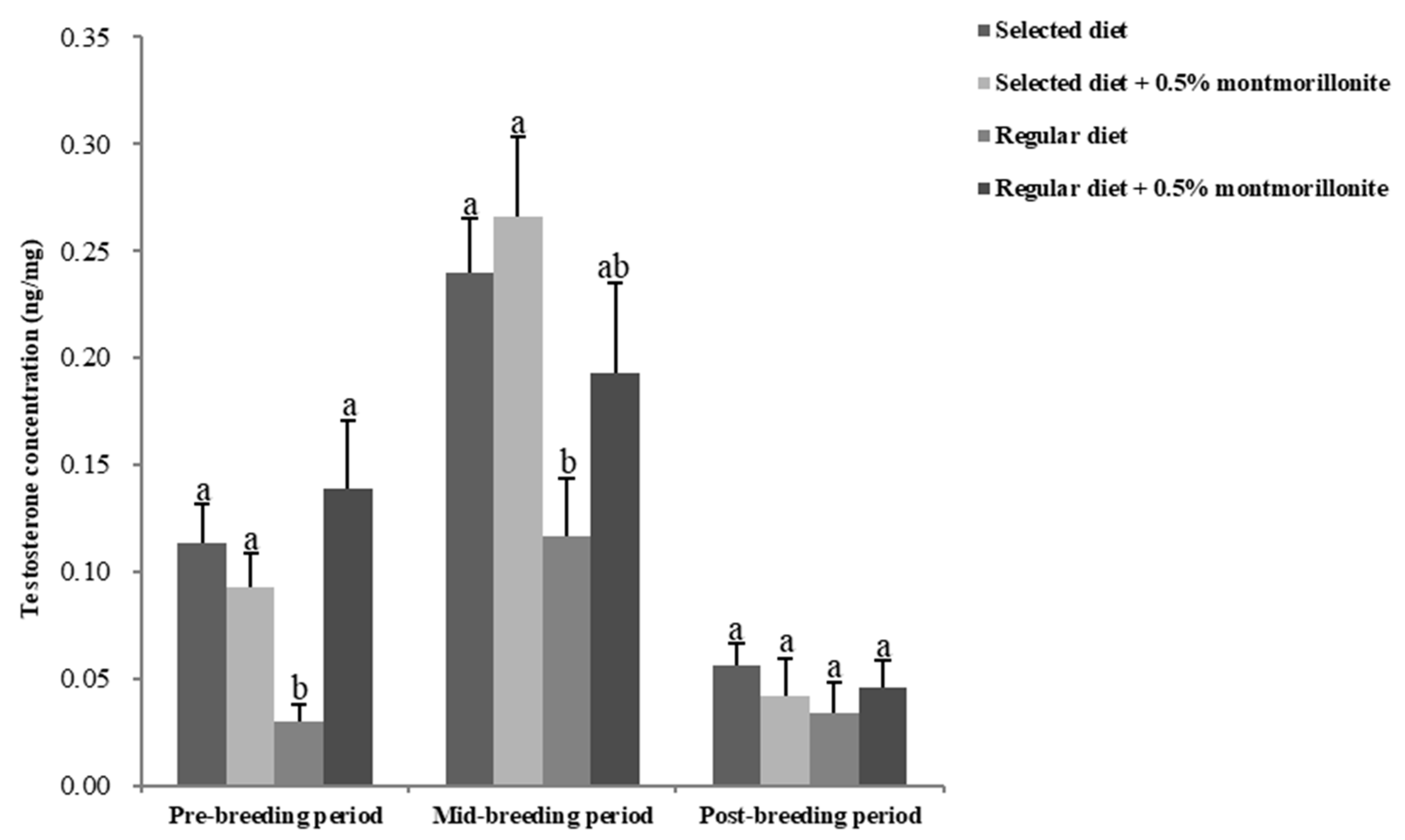
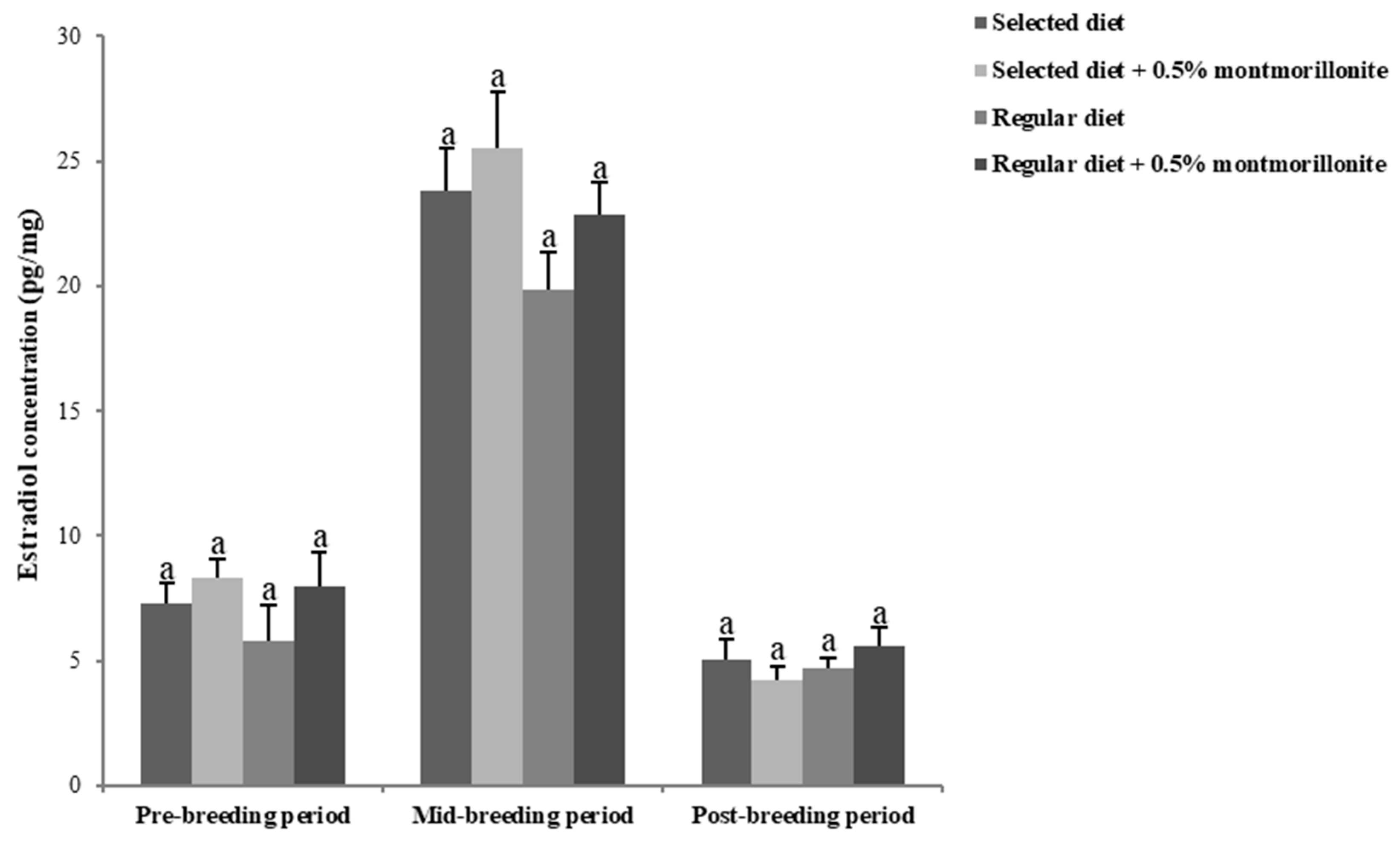
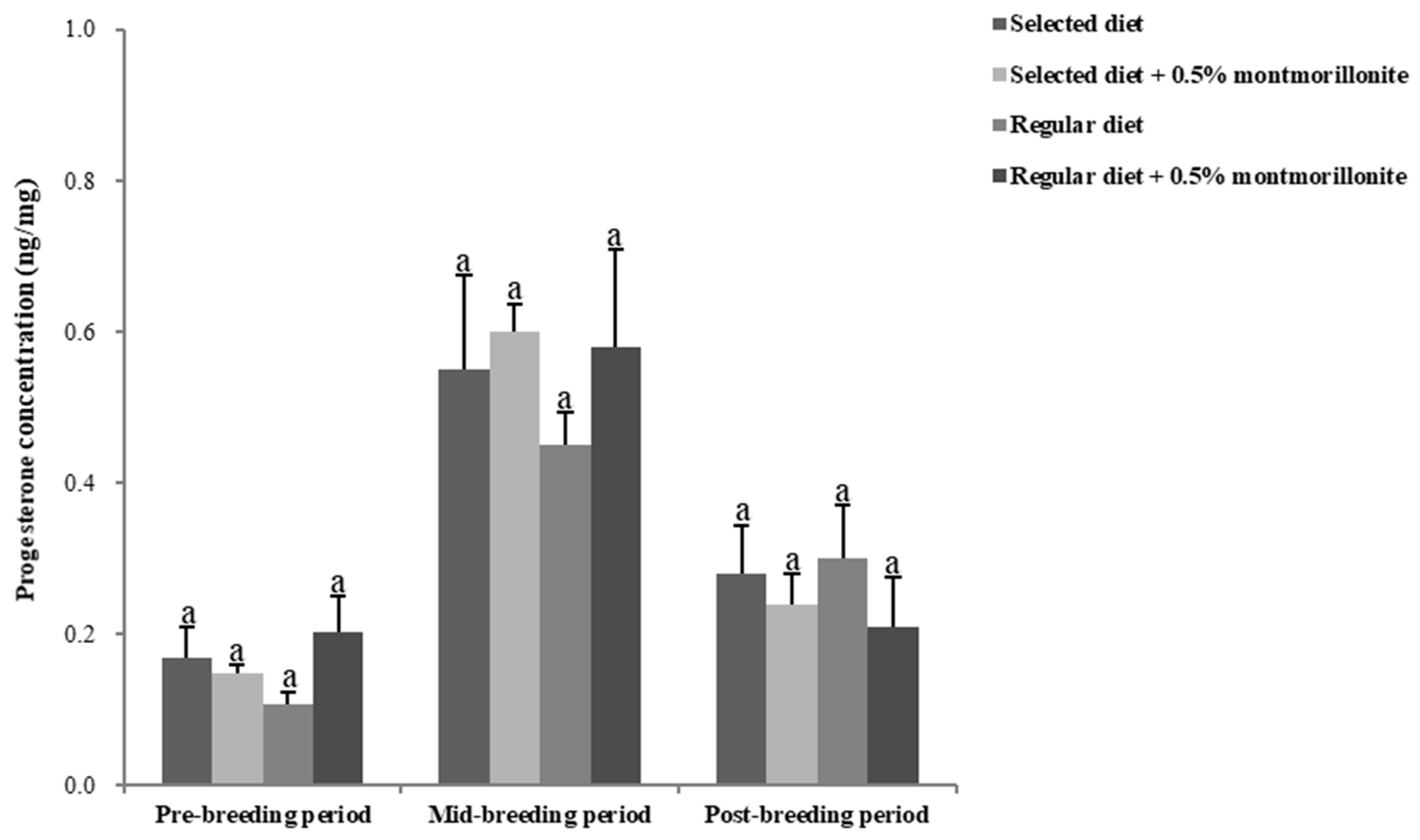
2.3. Sex Hormone Levels
2.4. Egg Quality
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Area
5.2. Chemicals, Animals and Diet
5.3. Determination of Mycotoxin Concentration in Diets
5.4. Behavioral Observations
5.5. Determination of Sex Hormones in the Feces
5.6. Measurement and Calculation of Egg Index
5.7. Incubation of Eggs
5.8. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Wood, G.E. Mycotoxins in foods and feeds in the United States. J. Anim. Sci. 1992, 70, 3941–3949. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, I.; Naehrer, K. Prevalence of mycotoxins in feedstuffs and feed surveyed worldwide in 2009 and 2010. Phytother. Med. 2012, 51, 175–192. [Google Scholar]
- Wang, Y.; Liu, S.L.; Zheng, H.; He, C.H.; Zhang, H.B. T-2 toxin, zearalenone and fumonisin B1 in feedstuffs from China. Food Addi. Contam. 2013, 6, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Jones, F.T.; Hagler, W.H.; Hamilton, P.B. Association of low levels of aflatoxins in feed with productivity losses in commercial broiler operations. Poult. Sci. 1982, 61, 861–868. [Google Scholar] [CrossRef]
- Diaz, D.E.; Smith, T.K. Mycotoxin Sequestering Agents: Practical Tools for Neutralization of Mycotoxins; Nottingham University Press: Nottingham, UK, 2005. [Google Scholar]
- Hamilton, P.B. Determining safe levels of mycotoxins. J. Food Prot. 1984, 7, 508–575. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Chowdhury, S.R.; Smith, T.K.; Boermans, H.J.; Sefton, A.E.; Downey, R.; Woodward, B. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on performance metabolism, hematology, and immunocompetence of duckings. Poult. Sci. 2005, 84, 1179–1185. [Google Scholar] [CrossRef]
- Yegani, M.; Smith, T.K.; Leeson, S.; Boermans, H.J. Effects of feeding grains naturally contaminated with Fusarium mycotoxins on performance and metabolism of broiler breeders. Poult. Sci. 2006, 85, 1541–1549. [Google Scholar] [CrossRef]
- Fischer, J.R.; Jain, A.V.; Shipes, D.A.; Osborne, J.S. Aflatoxin contamination of corn used as bait for deer in the southeastern United States. J. Wildl. Dis. 1995, 31, 570–572. [Google Scholar] [CrossRef] [Green Version]
- Quist, C.F.; Howerth, E.W.; Fisher, J.R.; Wyatt, R.D.; Miller, D.M.; Nettles, V.F. Evaluation of low-level aflatoxin in the diet of white-tailed deer. J. Wildl. Dis. 1997, 33, 112–121. [Google Scholar] [CrossRef] [Green Version]
- Quist, C.F.; Bounous, D.I.; Kilburn, J.V.; Nettles, V.F.; Wyatt, R.D. The effect of dietary aflatoxin on wild turkey poults. J. Wildl. Dis. 2000, 36, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyatt, R.D.; Doerr, J.A.; Hamilton, P.B.; Burmeister, H.R. Egg production, shell thickness, and other physiological parameters of laying hens affected by T-2 toxin. Appl. Microbiol. 1975, 29, 641–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafez, A.H.; Megalla, S.E.; Abdel-Fattah, H.M.; Kamel, Y.Y. Aflatoxin and alatoxicosis II. Effect of aflatoxin on ovaries and testicels in mayure fowls. Mycopathologia 1982, 77, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Price, W.D.; Lovel, R.A.; McChesney, D.G. Naturally occurring toxins in feedstuffs: Center for veterinary medicine perspective. J. Anim. Sci. 1993, 71, 2556–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahem, M.; Kersten, S.; Valenta, H.; Breves, G.; Beineke, A.; Hermeyer, K.; Dänicke, S. Effects of feeding deoxynivalenol (DON)-contaminated wheat to laying hens and roosters of different genetic background on the reproductive performance and health of the newly hatched chicks. Mycotoxin Res. 2014, 30, 131–140. [Google Scholar] [CrossRef]
- Phillips, T.D.; Afriyie-Gyawu, E.; Williams, J.; Huebner, H.; Ankrah, N.A.; Ofori-Adjei, D.; Jolly, P.; Johnson, N.; Taylor, J.; Marroquin-Cardona, A.; et al. Reducing human exposure to aflatoxin through the use of clay: A review. Food Addit. Contam. 2008, 25, 134–145. [Google Scholar] [CrossRef]
- Vila-Donat, P.; Marin, S.; Sanchis, V.; Ramos, A.J. A review of the mycotoxin adsorbing agents, with an emphasis on their multi-binding capacity, for animal feed decontamination. Food Chem. Toxicol. 2018, 114, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Kooli, F.; Liu, Y.; Al-Faze, R.; Al Suhaimi, A. Effect of acid activation of Saudi local clay mineral on removal properties of basic blue 41 from an aqueous solution. Appl. Clay Sci. 2015, 116–117, 23–30. [Google Scholar] [CrossRef]
- Zeng, L.; Wang, S.P. Adsorption of zearalenone by montmorillonite. Adv. Mater. Res. 2013, 683, 343–347. [Google Scholar] [CrossRef]
- Sposito, G.; Skipper, N.T.; Sutton, R.; Park, S.; Soper, A.K.; Greathouse, J.A. Surface geochemistry of the clay mineral. Proc. Natl. Acad. Sci. USA 1999, 96, 3358–3364. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhao, Z.; Deng, Y.; Zhou, Z.; Hou, J. Toxicity induced by F. poae-contaminated feed and the protective effect of Montmorillonite supplementation in broilers. Food Chem. Toxicol. 2014, 74, 120–130. [Google Scholar] [CrossRef]
- Wang, J.P.; Chi, F.; Kim, I.H. Effects of montmorillonite clay on growth performance, nutrient digestibility, vulva size, faecal microflora, and oxidative stress in weaning gilts challenged with zearalenone. Anim. Feed Sci. Technol. 2012, 178, 158–166. [Google Scholar] [CrossRef]
- Desheng, Q.; Fan, L.; Yanhu, Y.; Niya, Z. Adsorption of aflatoxin B1 on montmorillonite. Poult. Sci. 2005, 84, 959–961. [Google Scholar] [CrossRef] [PubMed]
- BirdLife International. BirdLife International: Species Factsheet: Grus Japonensis. Available online: http://www.birdlife.org (accessed on 20 March 2019).
- IUCN (International Union for Conservation of Nature). IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org (accessed on 20 March 2019).
- Ma, Z.J.; Li, W.J.; Wang, Z.J. Nature Conservation of Red-Crowned Cranes; Qinghua University Press: Beijing, China, 2000. [Google Scholar]
- Snyder, N.F.; Derrickson, S.R.; Beissinger, S.R.; Wiley, J.W.; Smith, T.B.; Toone, W.D.; Miller, B. Limitations of captive breeding in endangered species recovery. Conserv. Biol. 2002, 10, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.Q.; Wang, Z.; Gao, J.; Zhang, H.N.; Jiang, M.K. Population size and distribution patterns of captive red-crowned cranes (Grus japonensis) in zoos in China. J. Ecol. R. Environ. 2014, 30, 731–735. [Google Scholar]
- Li, Y.J.; Yang, D.H.; Zhao, W.L.; Wang, Z.Y.; Chen, H.; Wang, H.; Gao, Z.D. Study on artificial incubation about eggs of Red-crowned cranes. Chin. Poult. 2002, 24, 19–20. [Google Scholar]
- Jacquet, J.M. Photo-refractory period of the Muscovy duck (Cairina moschata): Endocrine and neuroendocrine response to day length after a full reproductive cycle. Br. Poult. Sci. 1997, 38, 209–216. [Google Scholar] [CrossRef]
- Sharp, P.J.; Dawson, A.; Lea, R.W. Control of luteinizing hormone and prolactin secretion in birds. Comp. Biochem. Physiol. 1998, 119, 275–282. [Google Scholar] [CrossRef]
- Bacon, W.L.; Kurginski-Noonan, B.A.; Yang, J. Effect of environmental lighting on early semen production and correlated hormonal responses in turkeys. Poult. Sci. 2000, 79, 1669–1678. [Google Scholar] [CrossRef] [Green Version]
- Hang, Y.M.; Shi, Z.D.; Li, X.W.; Liu, Z.; Liu, Y. Regulation of photoperiod on seasonal reproductive activities and endocrine in magang geese. Chin. J. Anim. Vet. Sci. 2008, 39, 29–36. [Google Scholar]
- Ellis, D.H.; Gee, G.F.; Mirande, C.M. Cranes: Their Biology, Husbandry, and Conservation; International Crane Foundation: Baraboo, WI, USA, 1996. [Google Scholar]
- Liu, D.W.; Zhou, Y.W.; Fei, Y.L.; Xie, C.P.; Liu, H.Y.; Lu, C.H.; Hou, S.L. A survey on the occurrence of mycotoxins in the feeds of captive red-crowned cranes (Grus japonensis) in some areas of China. J. Sichuan Agric. Univ. 2019, 37, 868–875. [Google Scholar]
- National Feed Safety Standards of China. Tolerance Limits for Ochratoxin a and Zearalenone in Feeds, Hygienical Standard for Feeds; GB13078.2–2006; China’s Ministry of Health. China’s General Administration of Quality Supervision, Inspection and Quarantine: Beijing, China, 2006.
- National Feed Safety Standards of China. Tolerance Limits for Deoxynivalenol in Formula Feed, Hygienicalstandard for Feeds; GB13078.3–2007; China’s General Administration of Quality Supervision, Inspection and Quarantine: Beijing, China, 2007.
- National Feed Safety Standards of China. Hygienical Standard for Feeds; GB13078–2001; China’s Ministry of Health. China’s General Administration of Quality Supervision, Inspection and Quarantine: Beijing, China, 2011.
- Bunmi, K.; Solomon, U.O.; Obinna, C.N.; Isiaka, A.L.; Sefater, G.; Patrick, B.N. Decontamination of T-2 Toxin in Maize by Modified Montmorillonite Clay. Toxins 2019, 11, 616. [Google Scholar]
- Ma, J.Q.; Lv, G.L.; Zhou, R.X. Effect of acid-treated and hexadecyltrimethylammonium bromide–modified montmorillonites on adsorption performance of mycotoxins. Environ. Sci. Pollut. Res. 2020, 27, 4284–4293. [Google Scholar] [CrossRef] [PubMed]
- Lasley, B.L.; Kirkpatrick, J.F. Monitoring ovarian function in captive and free-ranging wildlife by means of urinary and fecal steroids. J. Zoo Wildl. Med. 1990, 22, 23–31. [Google Scholar]
- Ványi, A.; Glávits, R.; Bata, A.; Kovács, F. Pathomorphological changes caused by T-2 trichothecene fusariotoxin in geese. Acta Vet. Hung. 1994, 42, 447–457. [Google Scholar]
- Jiang, S.Z.; Yang, Z.B.; Yang, W.R.; Yao, B.Q.; Zhao, H.; Liu, F.X. Effects of feeding purified zearalenone contaminated diets with or without clay enterosorbent on growth, nutrient availability, and genital organs in post-weaning female pigs. Asian Austral. J. Anim. 2010, 23, 74–81. [Google Scholar] [CrossRef]
- David, A.E.; Ellen, D.K.; Val, N.J.R. Testosterone and mate choice in the Dark-eyed Junco. Anim. Behav. 1997, 54, 1135–1146. [Google Scholar]
- Zhang, Y.Y.; Zheng, G.M.; Wang, H.; Ma, M.J. A study on the changes of the sex steroid extracted from the droppings of cabots tragopan (tragopan caboti) in capativity. J. Behav. Normal Univ. (Nat. Sci. Ed.) 2001, 37, 685–689. [Google Scholar]
- Wang, S.Y.; Tian, X.H.; Zhang, D.D.; Ma, X.F.; He, X.B. Relation between the fecal hormone concentration and reproductive behavior of captive red-crowned crane. Chin. J. Wildl. 2014, 35, 69–74. [Google Scholar]
- Kaya, A.; Ortatatli, M.; Ciftci, M.K.; Tuzcu, M. The effects of aflatoxin on the reproductive system of roosters. Res. Vet. Sci. 2002, 72, 29–36. [Google Scholar]
- Eraslan, G.; Akdoǧan, M.; Liman, B.C.; Kanbur, M.; Delibaş, N. Effects of dietary aflatoxin and hydrate sodium calcium aluminosillcate on triiodothyonine, thyroxine, thyrotrophin and testosterone levels in quails. Turk. J. Vet. Anim. Sci. 2006, 30, 41–45. [Google Scholar]
- Koga, T. Studies on the reproduction of cranes, especially on their aritificial incubation and breeding. J. Jpn. Assoc. of Zool. Gard. Aquar. 1961, 3, 51–58. [Google Scholar]
- King’ori, A.M. Review of the factors that influence egg fertility and hatchabilty in poultry. Int. J. Poult. Sci. 2011, 10, 483–492. [Google Scholar]
- Rizzi, L.; Simioli, M.; Roncada, P.; Zaghini, A. Aflatoxin B1 and clinoptilolite in feed for laying hens: Effects on egg quality, mycotoxin residues in livers, and hepatic mixed-function oxygenase activities. J. Food Prot. 2003, 66, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Jessen, K.A.; Beltran, R.; Starkl, V.; Schatzmayr, G.; Borutova, R.; Caldwell, D.J. Effects of mycotoxin-contaminated diets and deactivating compound in laying hens: 2 Effects on white shell egg quality and characteristics. Poult. Sci. 2012, 91, 2096–2104. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Ma, Q.G.; Fan, Y.; Ji, C.; Zhang, J.Y.; Liu, T.; Zhao, L.H. The toxic effects of combined aflatoxins an zearalenone in naturally contaminated diets on laying performance, egg quality and mycotoxins residues in eggs of layer and the protective effect of Bacillus subtilis biodegradation product. Food Chem. Toxicol. 2016, 90, 142–150. [Google Scholar] [CrossRef]
- Pasha, T.N.; Farooq, M.U.; Khattak, F.M.; Jabbar, M.A.; Khan, A.D. Effectiveness of sodium bentonite and two commercial products as aflatoxin absorbents in diets for broiler chickens. Anim. Feed Sci. Tech. 2007, 132, 103–110. [Google Scholar] [CrossRef]
- Osweiler, G.D.; Jagannatha, S.; Trampel, D.W.; Imerman, P.M.; Ensley, S.M.; Yoon, I.; Moore, D.T. Evaluation of XPC and prototypes on aflatoxin-challenged broilers. Poult. Sci. 2010, 89, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Kermanshahi, H.; Akbari, M.R.; Maleki, M.; Behgar, M. Effect of prolonged low level inclusion of aflatoxin B1 into diet on performance, nutrient digestibility, histopathology and blood enzymes of broiler chickens. J. Anim. Vet. Adv. 2007, 6, 686–692. [Google Scholar]
- Yildirim, E.; Yalcinkaya, I.; Kanbur, M.; Cinar, M.; Oruc, E. Effects of yeast glucomannan on performance, some biochemical parameters and pathological changes in experimental aflatoxicosis in broiler chickens. Rev. Med. Vet. 2011, 162, 413–420. [Google Scholar]
- Bergsjø, B.; Herstad, O.; Nafstad, I. Effects of feeding deoxynivalenol-contaminated oats on reproduction performance in white leghorn hens. Br. Poult. Sci. 1993, 34, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M.A.; Brake, J.; Hamilton, P.B.; Hagler, W.M., Jr.; Nesheim, S. Dietary exposure of broiler breeders to aflatoxin results in immune dysfunction in progeny chicks. Poult. Sci. 1998, 77, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Liu, D.W. Artificial incubation of Red Crowned Crane in winter habitat. Anim. Vet. Med. Husb. 2013, 45, 109–110. [Google Scholar]
- Liu, D.W.; Liu, H.Y.; Zhang, H.; Cao, M.C.; Sun, Y.; Wu, W.D.; Lu, C.H. Potential natural exposure of endangered red-crowned crane (Grus japonensis) to mycotoxins aflatoxin B1, deoxynivalenol, zearalenone, T-2 toxin, and ochratoxin A. J. Zhejiang Univ. Sci. B 2016, 17, 158–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Dietary Treatment | Mycotoxin (μg/kg) | ||||
|---|---|---|---|---|---|
| AFB1 | DON | ZEN | T-2 | OTA | |
| Selected diet | ND a | 16.4 ± 8.3 | 9.4 ± 3.8 | <LOQ b | <LOQ |
| Selected diet + 0.5% montmorillonite | ND | 18.1 ± 7.8 | 11.7 ± 5.2 | ND | <LOQ |
| Regular diet | 7.3 ± 2.5 | 776.7 ± 95.3 | 479.9 ± 38.7 | 51.6 ± 13.6 | <LOQ |
| Regular diet + 0.5% montmorillonite | 6.7 ± 3.1 | 875.2 ± 85.9 | 486.6 ± 43.4 | 62.7 ± 17.8 | <LOQ |
| Dietary Treatment | Courtship Times | Mating Times | Nesting Times |
|---|---|---|---|
| Selected diet | 60.3 ± 6.7 a | 24.0 ± 4.7 a | 45.0 ± 6.7 a |
| Selected diet + 0.5% montmorillonite | 72.0 ± 4.4 a | 23.3 ± 2.6 a | 40.3 ± 6.2 a |
| Regular diet | 36.7 ± 4.9 b | 11.7 ± 2.0 b | 25.7 ± 5.2 a |
| Regular diet + 0.5% montmorillonite | 58.3 ± 9.0 a | 26.5 ± 4.5 a | 38.0 ± 7.2 a |
| Dietary Treatment | Mid-Breeding Period | Post-Breeding Period | |||
|---|---|---|---|---|---|
| Courtship | Mating | Nesting | Laying and/or Hatching | Laying and/or Hatching | |
| Selected diet | 26.0 ± 2.7 a | 12.3 ± 2.5a | 4.3 ± 3.8 a | 822.3 ± 31.9 a | 57.0 ± 4.6 a |
| Selected diet + 0.5% montmorillonite | 23.3 ± 5.8 ab | 8.0 ± 2.3 ab | 5.7 ± 2.6 a | 851.7 ± 75.7 a | 72.0 ± 7.0 a |
| Regular diet | 11.3 ± 2.0 b | 3.7 ± 0.9 b | 6.3 ± 2.0 a | 789.0 ± 28.7 a | 65.3 ± 4.1a |
| Regular diet + 0.5% montmorillonite | 24.0 ± 4.7 ab | 10.7 ± 4.5 a | 2.3 ± 1.5 a | 774.7 ± 67.4 a | 60.7 ± 5.6 a |
| Dietary Treatment | Number of Eggs (n) | Egg Weight (g) | Shell Thickness (mm) | Egg Shape Index | Fertilization (%) | Hatching Rate (%) | Healthy Chick Rate (%) |
|---|---|---|---|---|---|---|---|
| Selected diet | 2.7 ± 0.7 a | 264.1 ± 3.1 a | 0.6 ± 0.01 a | 1.6 ± 0.0 a | 83.3 ± 16.7 a | 72.3 ± 14.7 a | 91.7 ± 8.3 a |
| Selected diet + 0.5% montmorillonite | 4.7 ± 0.9 a | 256.5 ± 2.0 a | 0.5 ± 0.01 ab | 1.5 ± 0.0 a | 91.7 ± 8.3 a | 100.0 ± 0.0 a | 89.0 ± 11.0 a |
| Regular diet | 3.7 ± 1.3 a | 234.7 ± 5.2 b | 0.5 ± 0.02 b | 1.6 ± 0.0 a | 75.0 ± 14.4 a | 100.0 ± 0.0 a | 72.3 ± 14.7 a |
| Regular diet + 0.5% montmorillonite | 4.3 ± 1.7 a | 258.3 ± 2.6 a | 0.6 ± 0.02 a | 1.6 ± 0.0 a | 91.7 ± 8.3 a | 83.3 ± 16.7 a | 91.7 ± 8.3 a |
| Ingredient | Percentage | Nutrition Index | Value |
|---|---|---|---|
| Corn | 41.2 | Metabolic (Mcal/kg) | 2.73 |
| Barely | 12.5 | Crude protein (%) | 21.2 |
| Soybean meal | 23 | Calcium (%) | 2.57 |
| Wheat bran | 10 | Phosphor (%) | 0.96 |
| Fish meal | 5 | ||
| Sodium chloride | 0.3 | ||
| Premix * | 1 | ||
| Shell power | 3 | ||
| Calcium phosphate | 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Gu, C.; Lu, C.; Wu, Q.; Kuca, K.; Wu, W. Red-Crowned Crane (Grus japonensis) Reproduction Was Improved by Inhibiting Mycotoxins with Montmorillonite in Feed. Toxins 2020, 12, 191. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030191
Liu D, Gu C, Lu C, Wu Q, Kuca K, Wu W. Red-Crowned Crane (Grus japonensis) Reproduction Was Improved by Inhibiting Mycotoxins with Montmorillonite in Feed. Toxins. 2020; 12(3):191. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030191
Chicago/Turabian StyleLiu, Dawei, Chao Gu, Changhu Lu, Qinghua Wu, Kamil Kuca, and Wenda Wu. 2020. "Red-Crowned Crane (Grus japonensis) Reproduction Was Improved by Inhibiting Mycotoxins with Montmorillonite in Feed" Toxins 12, no. 3: 191. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030191