Oligomer Formation and Insecticidal Activity of Bacillus thuringiensis Vip3Aa Toxin

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Insecticidal Activity of Residue Substituted Vip3Aa Mutants Against Neonates of S. litura
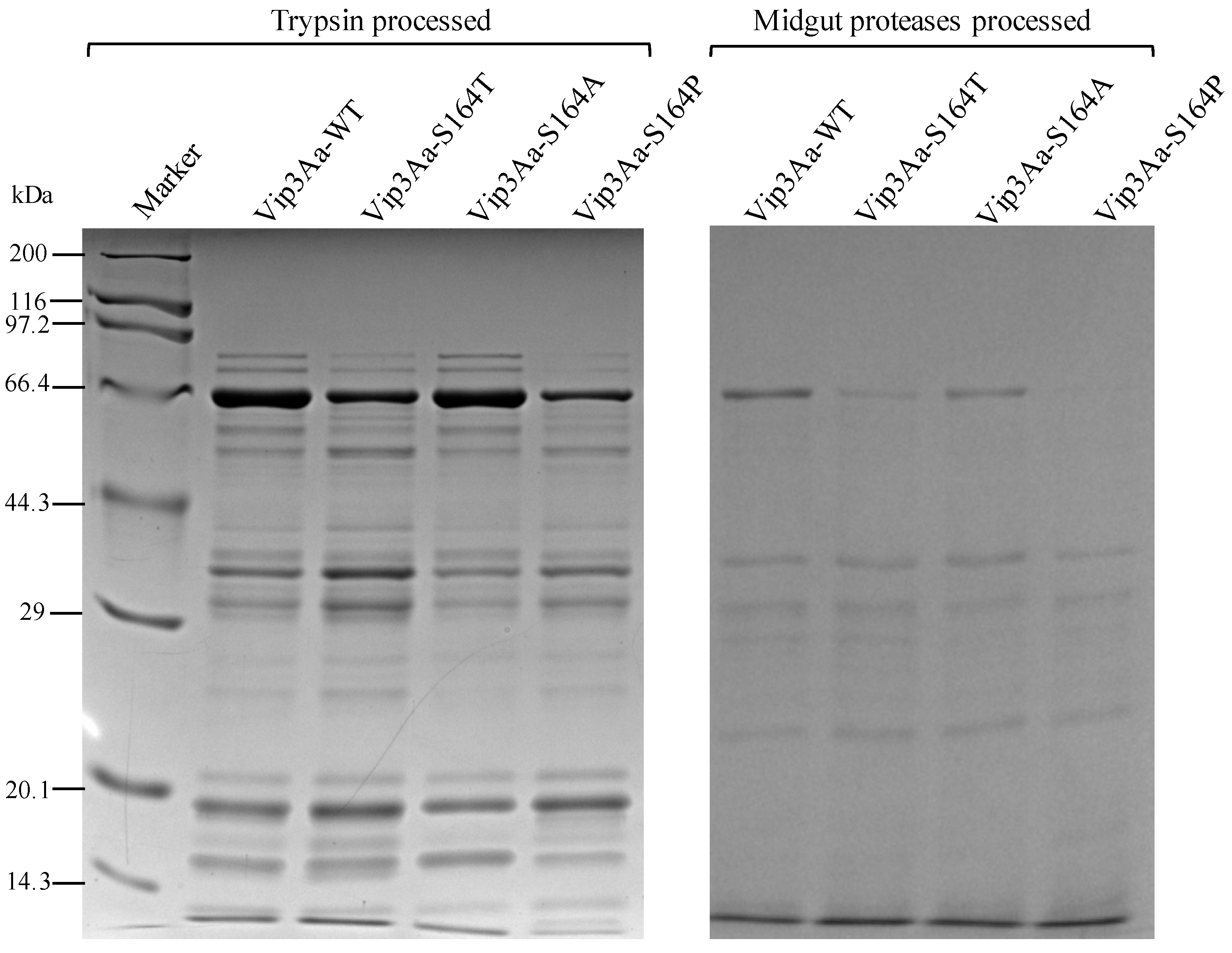
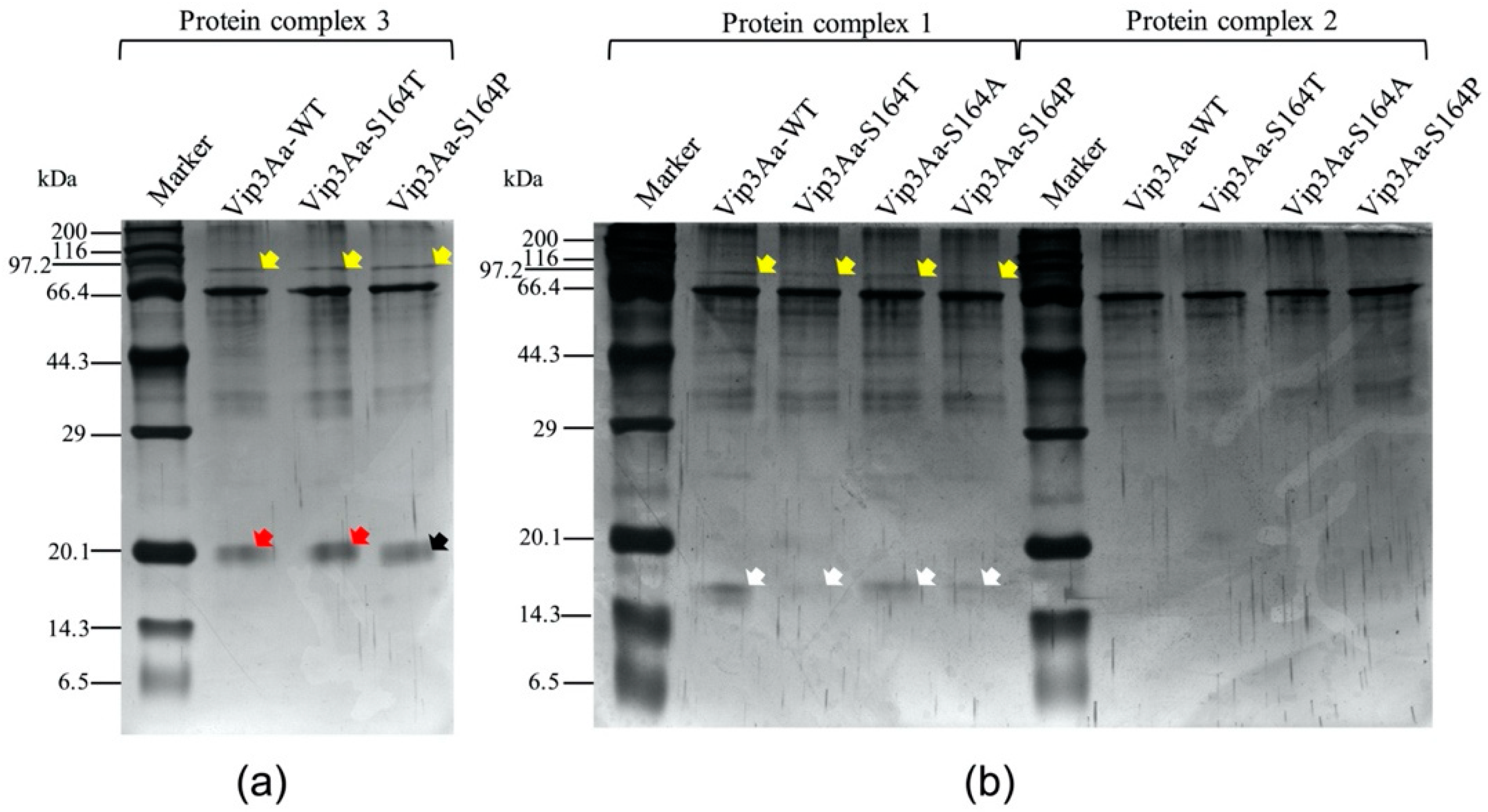
2.2. Analysis Vip3Aa Fragments After Proteolytic Processing
2.3. Analysis of Vip3Aa Protein Complexes by Native PAGE After Proteolytic Processing
2.4. Composition of the Three Protein Complexes Formed from Vip3Aa Toxins after Tryptic Processing
2.5. Identification of Tryptic Fragments from the 15, 17 and 19 kDa Protein Fragments by Peptide Fingerprinting
2.6. Correlation of Toxicity of Vip3Aa Protein with the Formation of the Protein Complex 3 Composed of 19 kDa and 65 kDa Peptides
3. Discussion
4. Materials and Methods
4.1. Site Directed Mutation on the vip3Aa Gene
4.2. Expression and Purification of Vip3Aa Proteins
4.3. Insects Rearing and Bioassays
4.4. In Vitro Proteolytic Processing of Vip3Aa Proteins
4.5. Analysis of Vip3Aa Proteins by the Native Gel and SDS-PAGE Gel
4.6. Identification of Trypsin-Processed Fragments
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Christou, P.; Capell, T.; Kohli, A.; Gatehouse, J.A.; Gatehouse, A.M. Recent developments and future prospects in insect pest control in transgenic crops. Trends Plant Sci. 2006, 11, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Chakraborty, A.; Sikdar, N.; Chakraborty, S.; Bhattacharyya, J.; Mitra, J.; Manna, A.; Dutta Gupta, S.; Sen, S.K. Marker-free transgenic rice expressing the vegetative insecticidal protein (Vip) of Bacillus thuringiensis shows broad insecticidal properties. Planta 2016, 244, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Chakroun, M.; Banyuls, N.; Bel, Y.; Escriche, B.; Ferré, J. Bacterial vegetative insecticidal proteins (Vip) from entomopathogenic bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 329–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, J.; Jiang, J.; Shu, C.; Wang, Z.; Song, F.; Geng, L.; Duan, J.; Zhang, J. Bacillus thuringiensis Vip1 Functions as a Receptor of Vip2 Toxin for Binary Insecticidal Activity against Holotrichia parallela. Toxins 2019, 11, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, Y.; Zhang, Y.; Shu, C.; Crickmore, N.; Wang, Q.; Du, L.; Song, F.; Zhang, J. Genomic sequencing identifies novel Bacillus thuringiensis Vip1/Vip2 binary and Cry8 toxins that have high toxicity to Scarabaeoidea larvae. Appl. Microbiol. Biotechnol. 2015, 99, 753–760. [Google Scholar] [CrossRef]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [Green Version]
- Palma, L.; de Escudero, I.R.; Maeztu, M.; Caballero, P.; Muñoz, D. Screening of vip genes from a Spanish Bacillus thuringiensis collection and characterization of two Vip3 proteins highly toxic to five lepidopteran crop pests. Biol. Control 2013, 66, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Sena, J.A.; Hernández-Rodríguez, C.S.; Ferré, J. Interaction of Bacillus thuringiensis Cry1 and Vip3A proteins with Spodoptera frugiperda midgut binding sites. Appl. Environ. Microbiol. 2009, 75, 2236–2237. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Miles, P.; Chen, J.-S. Brush border membrane binding properties of Bacillus thuringiensis Vip3A toxin to Heliothis virescens and Helicoverpa zea midguts. Biochem. Biophys. Res. Commun. 2006, 339, 1043–1047. [Google Scholar] [CrossRef]
- Gouffon, C.V.; Van Vliet, A.; Van Rie, J.; Jansens, S.; Jurat-Fuentes, J. Binding sites for Bacillus thuringiensis Cry2Ae toxin on heliothine brush border membrane vesicles are not shared with Cry1A, Cry1F, or Vip3A toxin. Appl. Environ. Microbiol. 2011, 77, 3182–3188. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-G.; Yang, A.-Z.; Shen, X.-H.; Hua, B.-G.; Shi, G.-L. Specific binding of activated Vip3Aa10 to Helicoverpa armigera brush border membrane vesicles results in pore formation. J. Invertebr. Pathol. 2011, 108, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, R.W. A review of Vip3A mode of action and effects on Bt Cry protein-resistant colonies of lepidopteran larvae. Southwest. Entomol. 2010, 35, 391–394. [Google Scholar] [CrossRef]
- Lee, M.K.; Walters, F.S.; Hart, H.; Palekar, N.; Chen, J.-S. The Mode of Action of the Bacillus thuringiensis Vegetative Insecticidal Protein Vip3A Differs from That of Cry1Ab δ-Endotoxin. Appl. Environ. Microbiol. 2003, 69, 4648–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banyuls, N.; Hernández-Rodríguez, C.; Van Rie, J.; Ferré, J. Critical amino acids for the insecticidal activity of Vip3Af from Bacillus thuringiensis: Inference on structural aspects. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellami, S.; Jemli, S.; Abdelmalek, N.; Cherif, M.; Abdelkefi-Mesrati, L.; Tounsi, S.; Jamoussi, K. A novel Vip3Aa16-Cry1Ac chimera toxin: Enhancement of toxicity against Ephestia kuehniella, structural study and molecular docking. Int. J. Biol. Macromol. 2018, 117, 752–761. [Google Scholar] [CrossRef]
- Zheng, M.; Evdokimov, A.G.; Moshiri, F.; Lowder, C.; Haas, J. Crystal structure of a Vip3B family insecticidal protein reveals a new fold and a unique tetrameric assembly. Protein Sci. 2019, 29, 824–829. [Google Scholar] [CrossRef]
- Yu, C.G.; Mullins, M.A.; Warren, G.W.; Koziel, M.G.; Estruch, J.J. The Bacillus thuringiensis vegetative insecticidal protein Vip3A lyses midgut epithelium cells of susceptible insects. Appl. Environ. Microbiol. 1997, 63, 532–536. [Google Scholar] [CrossRef] [Green Version]
- Banyuls, N.; Hernández-Martínez, P.; Quan, Y.; Ferré, J. Artefactual band patterns by SDS-PAGE of the Vip3Af protein in the presence of proteases mask the extremely high stability of this protein. Int. J. Biol. Macromol. 2018, 120, 59–65. [Google Scholar] [CrossRef]
- Hamadou-Charfi, D.B.; Boukedi, H.; Abdelkefi-Mesrati, L.; Tounsi, S.; Jaoua, S. Agrotis segetum midgut putative receptor of Bacillus thuringiensis vegetative insecticidal protein Vip3Aa16 differs from that of Cry1Ac toxin. J. Invertebr. Pathol. 2013, 114, 139–143. [Google Scholar] [CrossRef]
- Bel, Y.; Banyuls, N.; Chakroun, M.; Escriche, B.; Ferré, J. Insights into the structure of the Vip3Aa insecticidal protein by protease digestion analysis. Toxins 2017, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Quan, Y.; Ferré, J. Structural Domains of the Bacillus thuringiensis Vip3Af Protein Unraveled by Tryptic Digestion of Alanine Mutants. Toxins 2019, 11, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Martínez, P.; Hernández-Rodríguez, C.S.; Van Rie, J.; Escriche, B.; Ferré, J. Insecticidal activity of Vip3Aa, Vip3Ad, Vip3Ae, and Vip3Af from Bacillus thuringiensis against lepidopteran corn pests. J. Invertebr. Pathol. 2013, 113, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Chakroun, M.; Ferré, J. In vivo and in vitro binding of Vip3Aa to Spodoptera frugiperda midgut and characterization of binding sites by 125I radiolabeling. Appl. Environ. Microbiol. 2014, 80, 6258–6265. [Google Scholar] [CrossRef] [Green Version]
- Gayen, S.; Hossain, M.A.; Sen, S.K. Identification of the bioactive core component of the insecticidal Vip3A toxin peptide of Bacillus thuringiensis. J. Plant Biochem. Biotechnol. 2012, 21, 128–135. [Google Scholar] [CrossRef]
- Qin, H.; Ye, Z.; Huang, S.; Ding, J.; Luo, R. The correlation of the different host plants with preference level, life duration and survival rate of Spodoptera litura Fabricius. Chin. J. Eco-Agric. 2004, 12, 40–42. [Google Scholar]
- Qiong, L.; Cao, G.; Zhang, L.; Liang, G.; Gao, X.; Zhang, Y.; Guo, Y. The binding characterization of Cry insecticidal proteins to the brush border membrane vesicles of Helicoverpa armigera, Spodoptera exigua, Spodoptera litura and Agrotis ipsilon. J. Integr. Agric. 2013, 12, 1598–1605. [Google Scholar]
- Song, F.; Lin, Y.; Chen, C.; Shao, E.; Guan, X.; Huang, Z. Insecticidal activity and histopathological effects of Vip3Aa protein from Bacillus thuringiensis on Spodoptera litura. J. Microbiol. Biotechnol. 2016, 26, 1774–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Xu, N.; Huang, X.; Wang, W.; Cheng, J.; Wu, K.; Shen, Z. Bacillus thuringiensis Vip3 mutant proteins: Insecticidal activity and trypsin sensitivity. Biocontrol Sci. Technol. 2007, 17, 699–708. [Google Scholar] [CrossRef]
- Kunthic, T.; Surya, W.; Promdonkoy, B.; Torres, J.; Boonserm, P. Conditions for homogeneous preparation of stable monomeric and oligomeric forms of activated Vip3A toxin from Bacillus thuringiensis. Eur. Biophys. J. 2017, 46, 257–264. [Google Scholar] [CrossRef]
- Palma, L.; Scott, D.J.; Harris, G.; Din, S.-U.; Williams, T.L.; Roberts, O.J.; Young, M.T.; Caballero, P.; Berry, C. The Vip3Ag4 insecticidal protoxin from Bacillus thuringiensis adopts a tetrameric configuration that is maintained on proteolysis. Toxins 2017, 9, 165. [Google Scholar] [CrossRef]
- Zack, M.D.; Sopko, M.S.; Frey, M.L.; Wang, X.; Tan, S.Y.; Arruda, J.M.; Letherer, T.T.; Narva, K.E. Functional characterization of Vip3Ab1 and Vip3Bc1: Two novel insecticidal proteins with differential activity against lepidopteran pests. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, W.; Sha, L.; Zhou, J.; Huang, Z.; Guan, X. Functional analysis of active site residues of Bacillus thuringiensis WB7 chitinase by site-directed mutagenesis. World J. Microbiol. Biotechnol. 2009, 25, 2147–2155. [Google Scholar] [CrossRef]
- Guan, K.L.; Dixon, J.E. Eukaryotic proteins expressed in Escherichia coli: An improved thrombin cleavage and purification procedure of fusion proteins with glutathione S-transferase. Anal. Biochem. 1991, 192, 262. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kain, W.C.; Zhao, J.-Z.; Janmaat, A.F.; Myers, J.; Shelton, A.M.; Wang, P. Inheritance of resistance to Bacillus thuringiensis Cry1Ac toxin in a greenhouse-derived strain of cabbage looper (Lepidoptera: Noctuidae). J. Econ. Entomol. 2004, 97, 2073–2078. [Google Scholar] [CrossRef]
- Software, L. POLO-PC: A User’s Guide to Probit or Logit Analysis; LeOra Software: Berkeley, CA, USA, 1987. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LC50 (95% CI, μg/mL) | Slope | Χ2 (df, p) | Toxicity-Ratio | |
|---|---|---|---|---|
| Vip3Aa-WT | 1.69 (1.36–2.04) | 3.50 ± 0.47 | 0.34 (4, 0.08) | 1.0 |
| GST-Vip3Aa-S164T | 2.62 (2.24–3.17) | 4.19 ± 0.47 | 8.88 (8, 1.11) | 0.65 |
| GST-Vip3Aa-S164A | >480 | - | - | <0.0035 |
| GST-Vip3Aa-S164P | >480 | - | - | <0.0035 |
| GST-Vip3Aa-K152A | 2.23 (1.67–2.83) | 3.16 ± 0.63 | 0.005 (3, 0.002) | 0.76 |
| GST-Vip3Aa-D154A | 1.45 (1.36–1.55) | 8.80 ± 0.99 | 2.74 (4, 0.69) | 1.17 |
| Primers | Sequence 5′–3′ |
|---|---|
| P-3Aa-F | CATGCCATGGACATGAACAAGAATAATACTAAAT |
| P-3Aa-R | CGAGCTCTTACTTAATAGAGACATCGT |
| P-164P-R | TTCAGTAAGTGTaggGTTAATAAGTACA |
| P-164P-F | ATGTACTTATTAACcctACACTTACTG |
| P-164T-R | TTCAGTAAGTGTggtGTTAATAAGTACA |
| P-164T-F | GTACTTATTAACaccACACTTACTGAAA |
| P-164A-F | ACTTATTAACgcgACACTTACTG |
| P-164A-R | AGTAAGTGTcgcGTTAATAAGTA |
| P-152A-F | GATTTCTGATgcgTTGGATATTA |
| P-152A-R | ATAATATCCAAcgcATCAGAAAT |
| P-154A-F | TGATAAGTTGgcgATTATTAATG |
| P-154A-R | ATTAATAATcgcCAACTTATCAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, E.; Zhang, A.; Yan, Y.; Wang, Y.; Jia, X.; Sha, L.; Guan, X.; Wang, P.; Huang, Z. Oligomer Formation and Insecticidal Activity of Bacillus thuringiensis Vip3Aa Toxin. Toxins 2020, 12, 274. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040274
Shao E, Zhang A, Yan Y, Wang Y, Jia X, Sha L, Guan X, Wang P, Huang Z. Oligomer Formation and Insecticidal Activity of Bacillus thuringiensis Vip3Aa Toxin. Toxins. 2020; 12(4):274. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040274
Chicago/Turabian StyleShao, Ensi, Aishan Zhang, Yaqi Yan, Yaomin Wang, Xinyi Jia, Li Sha, Xiong Guan, Ping Wang, and Zhipeng Huang. 2020. "Oligomer Formation and Insecticidal Activity of Bacillus thuringiensis Vip3Aa Toxin" Toxins 12, no. 4: 274. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12040274