Absence of Cyanotoxins in Llayta, Edible Nostocaceae Colonies from the Andes Highlands

 , , and
, , and
Abstract
:1. Introduction
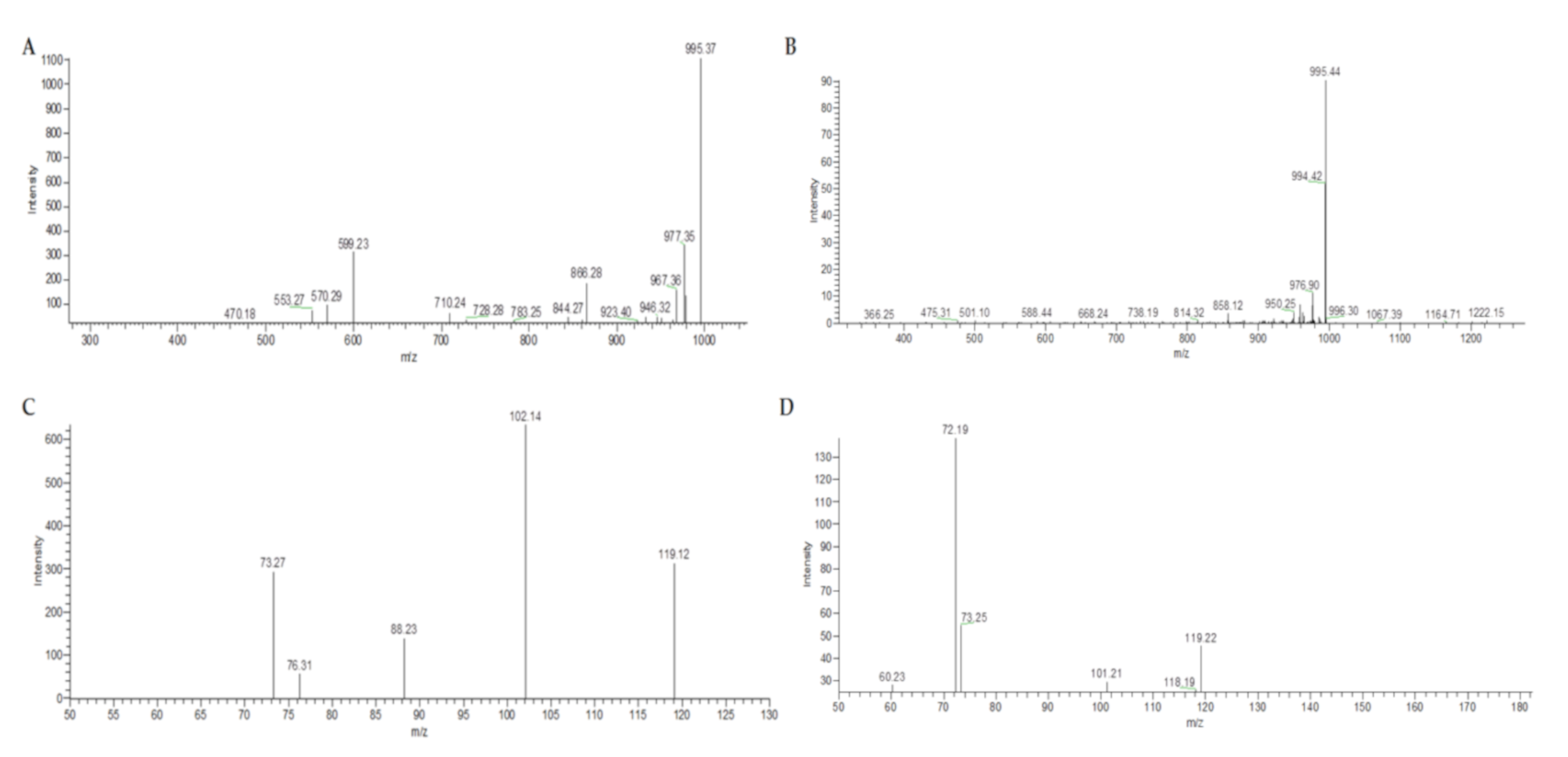
2. Results and Discussion
3. Conclusions
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Welker, M.; von Dören, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, H.; King, S.R.; Banack, S.A.; Webster, C.; Callanaupa, W.J.; Cox, P.A. Cyanobacteria (Nostoc commune) used as a dietary item in Peruvian highlands produce the neurotoxic amino acid BMAA. J. Ethnopharmacol. 2008, 118, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.S.; Cianca, R.C.C.; Almeida, C.M.R.; Vasconcelos, V.M. Determination of the non protein amino acid β-N-methylamino-l-alanine in estuarine cyanobacteria by capillary electrophoresis. Toxicon 2011, 58, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.; Costa, M.; Moreira, C.; Vasconcelos, V.M.; Baptista, M.S. Screening of BMAA-producing cyanobacteria in cultured isolates and in in situ blooms. J. Appl. Phycol. 2017, 29, 879–888. [Google Scholar] [CrossRef]
- Cirés, S.; Caser, M.C.; Quesada, A. Toxicity at the edge of life: A review on cyanobacterial toxins from extreme environments. Mar. Drugs 2017, 15, 233. [Google Scholar] [CrossRef] [PubMed]
- Buono, S.; Langellitti, A.L.; Martello, A.; Rinna, F.; Fogliano, V. Functional ingredients from microalgae. Food Funct. 2014, 5, 1669–1685. [Google Scholar] [CrossRef] [PubMed]
- Bertonio, L. Vocabulario de la Lengua Aymara. Colección Biblioteca Nacional de Chile: Impreso en la Compañía de Jesús, Perú. 1612. Available online: http://www.memoriachilena.cl/602/w3-article-8656.html (accessed on 26 July 2019).
- Aldave-Pajares, A. Algas andino peruanas como recurso hidrobiológico alimentario. Bol. Lima 1985, 7, 66–72. [Google Scholar]
- Gómez-Silva, B.; Mendizabal, C.; Tapia, I.; Olivares, H. Microalgas del norte de Chile. IV. Composición química de Nostoc commune Llaita. Rev. Invest. Cient. Tecnol. Cienc. Mar. 1994, 3, 19–25. [Google Scholar]
- Galetović, A.; Araya, J.; Gómez-Silva, B. Composición bioquímica y toxicidad de colonias comestibles de la cianobacteria andina Nostoc sp. Llayta. Rev. Chil. Nutr. 2017, 44, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Rivera, M.; Galetović, A.; Licuime, R.; Benito Gómez-Silva, B. A microethnographic and ethnobotanical approach to Llayta consumption among the Andes feeding practices. Foods 2018, 7, 202. [Google Scholar] [CrossRef] [Green Version]
- Jasser, I.; Bukowska, A.; Humbert, J.F.; Haukka, K.; Fewer, D.P. Analysis of Toxigenic Cyanobacterial Communities through Denaturing Gradient Gel Electrophoresis. In Molecular Tools for the Detection and Quantification of Toxigenic Cyanobacteria; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 263–275. [Google Scholar]
- Sivonen, K.; Carmichael, W.; Namikoshi, K.; Rinehart, I.; Dahlem, A.M.; Niemela, S.I. Isolation and Characterization of Hepatotoxic Microcystin Homologs from the Filamentous Freshwater Cyanobacterium Nostoc sp. Strain 152. Appl. Environ. Microbiol. 1990, 56, 2650–2657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, I.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Rikkinen, J.; Sivonen, K. Discovery of Rare and Highly Toxic Microcystins from Lichen-Associated Cyanobacterium Nostoc sp. Strain IO-102-I. Appl. Environ. Microbiol. 2004, 70, 5756–5763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy-Lachapelle, A.; Solliec, M.; Sauvé, S.; Gagnon, C. A Data-Independent Methodology for the Structural Characterization of Microcystins and Anabaenopeptins Leading to the Identification of Four New Congeners. Toxins 2019, 11, 619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural Diversity, Characterization and Toxicology of Microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikalsen, B.; Boison, G.; Skulberg, O.M.; Fastner, J.; William Davies, W.; Gabrielsen, T.M.; Rudi, K.; Jakobsen, K.S. Natural Variation in the Microcystin Synthetase Operon mcyABC and Impact on Microcystin Production in Microcystis Strains. J. Bacteriol. 2003, 185, 2774–2785. [Google Scholar] [CrossRef] [Green Version]
- Via-Ordorika, L.; Fastner, J.; Kurmayer, R.; Hisbergues, M.; Dittmann, E.; Komarek, J.; Erhard, M.; Chorus, I. Distribution of Microcystin-Producing and Non-Microcystin-Producing Microcystis sp. in European Freshwater Bodies: Detection of Microcystins and Microcystin Genes in Individual Colonies. Syst. Appl. Microbiol. 2004, 27, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Kurmayer, R.; Christiansen, G. The genetic basis of toxin production in Cyanobacteria. Freshw. Rev. 2009, 2, 31–50. [Google Scholar] [CrossRef]
- Rantala, A.; Fewer, D.P.; Hisbergues, M.; Rouhiainen, L.; Vaitomaa, J.; Börner, T.; Sivonen, K. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Fan, H.; Ma, F.; McCarron, P.; Thomas, K.; Tanga, X.; Quilliam, M.A. Elucidation of matrix effects and performance of solid-phase extraction for LC-MS/MS analysis of b-N-methylamino-L-alanine (BMAA) and 2,4-diaminobutyric acid (DAB) neurotoxins in cyanobacteria. Analyst 2012, 137, 1210–1219. [Google Scholar] [CrossRef]
- McCarron, P.; Logan, A.C.; Giddings, S.D.; Quilliam, M.A. Analysis of β-N-methylamino-L-alanine (BMAA) in Spirulina-containing supplements by liquid chromatography-tandem mass spectrometry. Aquat. Biosyst. 2014, 10, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Beach, D.G.; Kerrin, E.S.; Giddings, S.D.; Quilliam, M.A.; McCarron, P. Differential Mobility-Mass Spectrometry Double Spike Isotope Dilution Study of Release of β-Methylaminoalanine and Proteinogenic Amino Acids during Biological Sample Hydrolysis. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saker, M.L.; Jungblut, A.-D.; Neilan, B.; Rawn, T.; Vasconcelos, V.M. Detection of microcystin synthethase genes in health food supplements containing the freshwater cyanobacterium Aphanizomenon flos-aquae. Toxicon 2005, 46, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Rellan, S.; Osswald, J.; Saker, M.; Gago, A.; Vasconcelos, V.M. First detection of anatoxin-a in human and animal dietary supplements containing cyanobacteria. Food Chem. Toxicol. 2009, 47, 2189–2195. [Google Scholar] [CrossRef]
- Hisbergues, M.; Christiansen, G.; Rouhiainen, L.; Sivonen, K.; Borner, T. PCR-based identification of microcystin-producing genotypes of different cyanobacterial genera. Arch. Microbiol. 2003, 180, 402–410. [Google Scholar] [CrossRef]
- Jungblut, A.D.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin, synthetase genes in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casero, M.C.; Ballot, A.; Agha, R.; Quesada, A.; Cirés, S. Characterization of saxitoxin production and release and phylogeny of sxt genes in paralytic shellfish poisoning toxin-producing Aphanizomenon gracile. Harmful Algae 2014, 37, 28–37. [Google Scholar] [CrossRef]


{kind=link}
| Target Gene | Primer Pair | Target Group Producers | Size (bps) | References |
|---|---|---|---|---|
| mcyA | mcyA-Cd1F/mcyA-Cd1R | Microcystin | 297 | [26] |
| mcyE | HEPF/HEPR | Microcystin and nodularin | 472 | [27] |
| cyrJ | cynsulF; cylnamR | Cylindrospermopsin | 586 | [28] |
| anaC | anaC-genF/anaC-genR | Anatoxin | 366 | [28] |
| sxtA | sxtAF/sxtAR | Saxitoxin | 683 | [29] |
| sxtG | sxtGF/sxtGR | Saxitoxin | 893 | [29] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galetović, A.; Azevedo, J.; Castelo-Branco, R.; Oliveira, F.; Gómez-Silva, B.; Vasconcelos, V. Absence of Cyanotoxins in Llayta, Edible Nostocaceae Colonies from the Andes Highlands. Toxins 2020, 12, 382. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060382
Galetović A, Azevedo J, Castelo-Branco R, Oliveira F, Gómez-Silva B, Vasconcelos V. Absence of Cyanotoxins in Llayta, Edible Nostocaceae Colonies from the Andes Highlands. Toxins. 2020; 12(6):382. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060382
Chicago/Turabian StyleGaletović, Alexandra, Joana Azevedo, Raquel Castelo-Branco, Flavio Oliveira, Benito Gómez-Silva, and Vitor Vasconcelos. 2020. "Absence of Cyanotoxins in Llayta, Edible Nostocaceae Colonies from the Andes Highlands" Toxins 12, no. 6: 382. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060382