Single-Dose Toxicity of Individual and Combined Sterigmatocystin and 5-Methoxysterigmatocistin in Rat Lungs

 ,
,  , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. LDH Activity
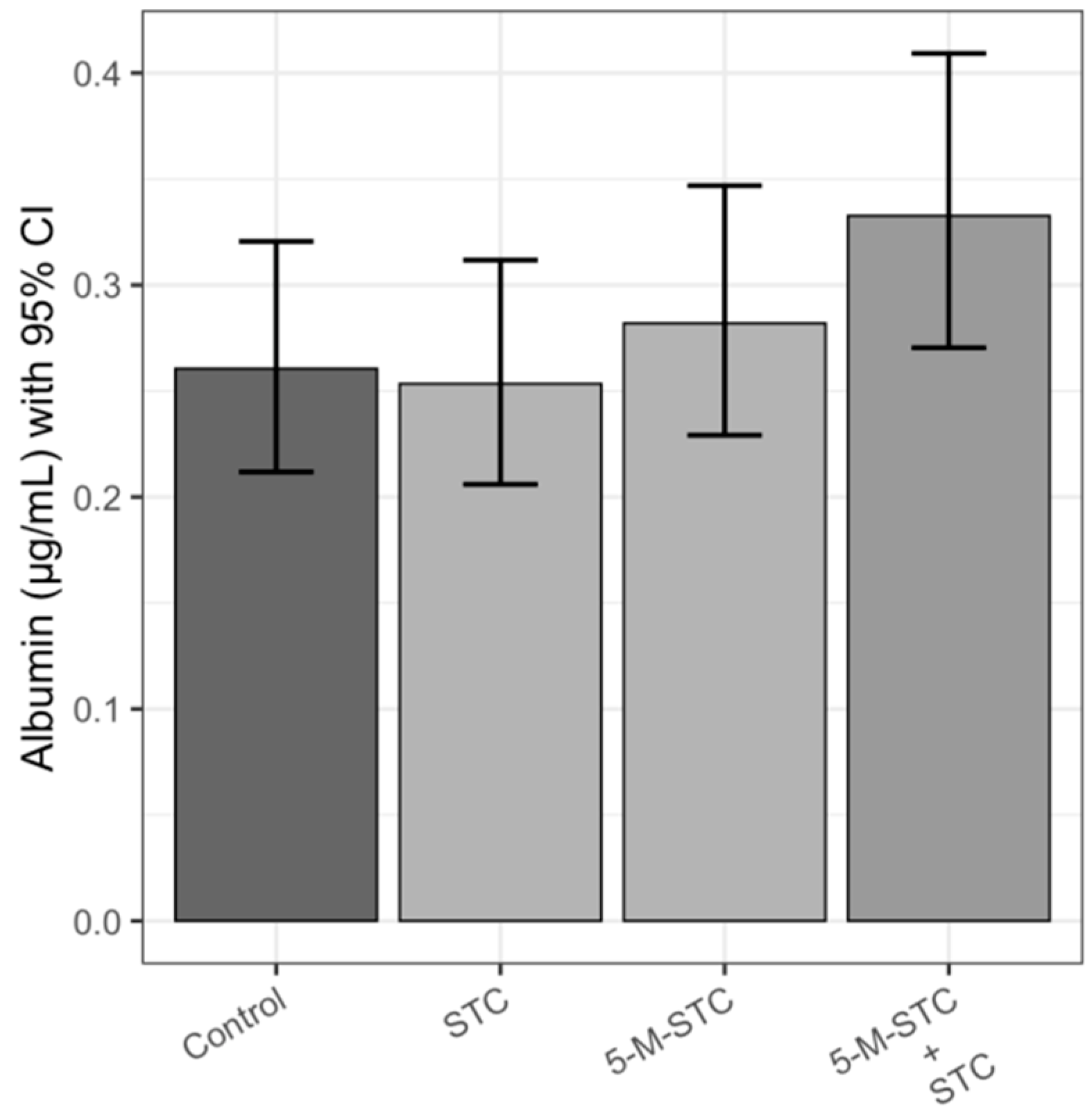
2.2. Albumin
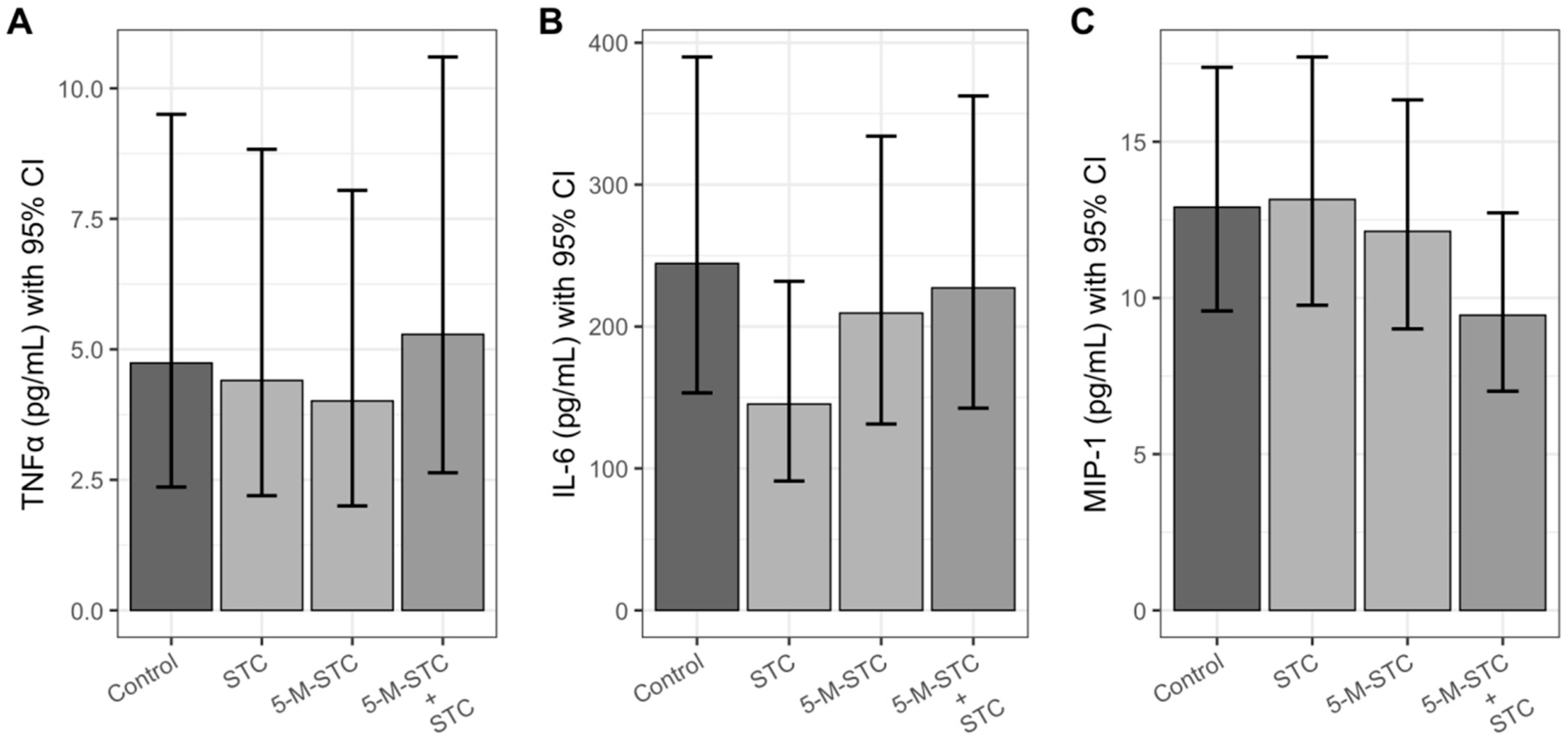
2.3. Pro-Inflammatory Cytokines and Chemokine
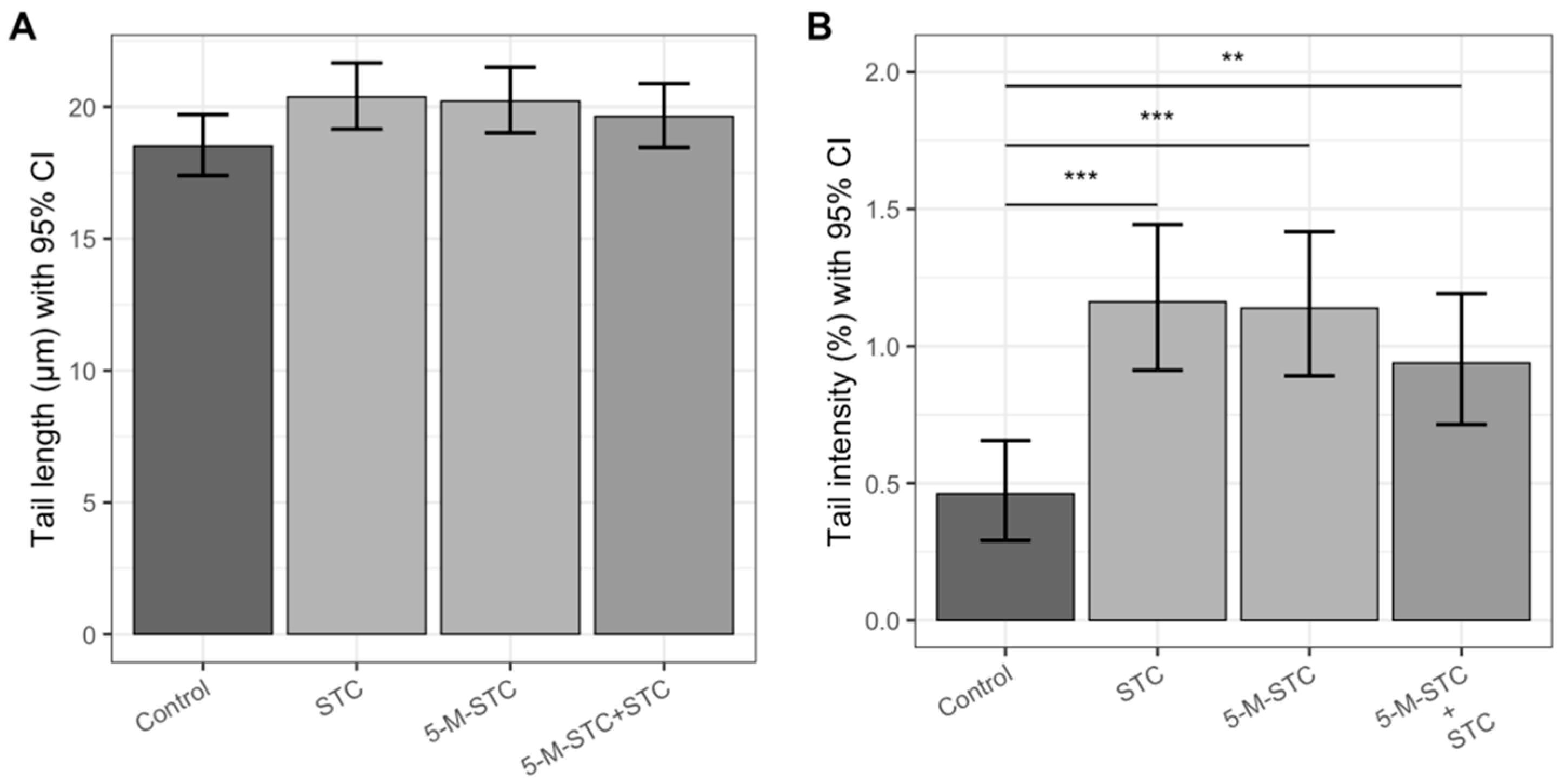
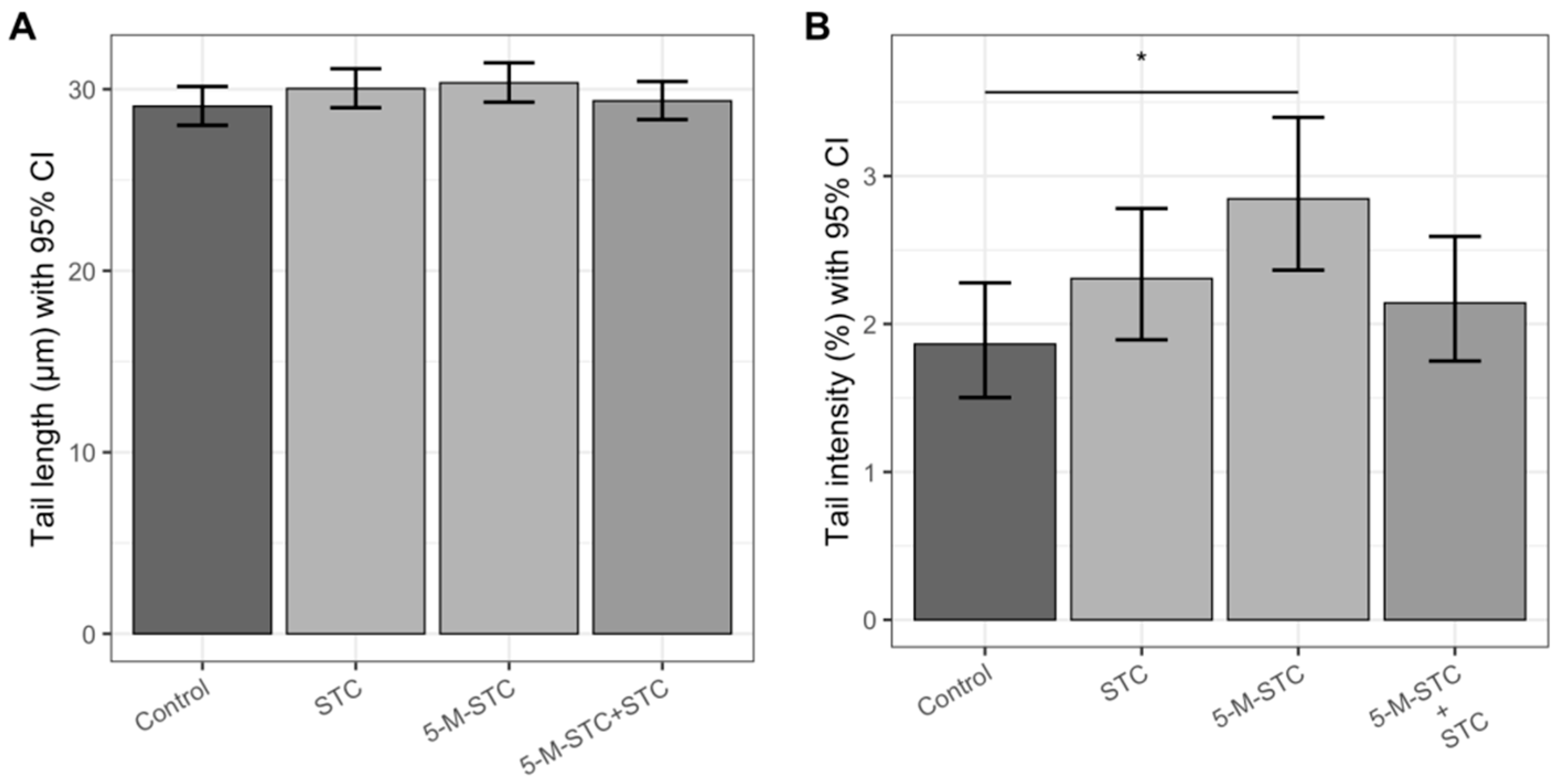
2.4. DNA Damage Measured by Alkaline and Neutral Comet Assays
3. Discussion
4. Materials and Methods
4.1. Experimental Design and BALF Sampling
4.2. LDH Activity Analysis
4.3. Albumin Analysis
4.4. Cytokines and Chemokine Analysis
4.5. Alkaline and Neutral Comet Assay
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bloom, E.; Bal, K.; Nyman, E.; Must, A.; Larsson, L. Mass spectrometry-based strategy for direct detection and quantification of some mycotoxins produced by Stachybotrys and Aspergillus spp. in indoor environments. Appl. Env. Microbiol. 2007, 73, 4211–4217. [Google Scholar] [CrossRef] [Green Version]
- Engelhart, S.; Loock, A.; Skutlarek, D.; Sagunski, H.; Lommel, A.; Färber, H.; Exner, M. Occurrence of toxigenic Aspergillus versicolor isolates and sterigmatocystin in carpet dust from damp indoor environments. Appl. Env. Microbiol. 2002, 68, 3886–3890. [Google Scholar] [CrossRef] [Green Version]
- Täubel, M.; Hyvärinen, A. Occurrence of mycotoxins in indoor environments. In Environmental Mycology in Public Health: Fungi and Mycotoxins Risk Assessment and Management; Viegas, C., Pinheiro, A.C., Sabino, R., Viegas, S., Brandão, J., Veríssimo, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 299–323. ISBN 9780124114715. [Google Scholar]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef] [PubMed]
- Jakšić Despot, D.; Kocsubé, S.; Bencsik, O.; Kecskeméti, A.; Szekeres, A.; Vágvölgyi, C.; Varga, J.; Šegvić Klarić, M. New sterigmatocystin-producing species of Aspergillus section Versicolores from indoor air in Croatia. Mycol. Prog. 2016, 16, 63–72. [Google Scholar] [CrossRef]
- Šegvić Klarić, M.; Jakšić, D.; Kocsubé, S.; Kifer, D.; Sulyok, M.; Jelić, D.; Šarkanj, B. Post-flood indoor occurrence of toxigenic Aspergilli from the Versicolores clade: Is it dangerous? In Proceedings of the Romanian Journal of Laboratory Medicine, Timisoara, Romania, 13–15 September 2018; Volume 26, pp. S12–S13. [Google Scholar]
- Jakšić Despot, D.; Kocsubé, S.; Bencsik, O.; Kecskeméti, A.; Szekeres, A.; Vágvölgyi, C.; Varga, J.; Šegvić Klarić, M. Species diversity and cytotoxic potency of airborne sterigmatocystin-producing Aspergilli from the section Versicolores. Sci. Total Environ. 2016, 562, 296–304. [Google Scholar] [CrossRef]
- Cabaret, O.; Puel, O.; Botterel, F.; Delaforge, M.; Bretagne, S. Metabolic detoxification pathways for 5-methoxy-sterigmatocystin in primary tracheal epithelial cells. Xenobiotica 2013, 44, 1–9. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the risk for public and animal health related to the presence of sterigmatocystin in food and feed. EFSA J. 2013, 11, 1–81. [Google Scholar]
- IARC. Sterigmatocystin. Iarc Monogr. Eval. Carcinog. Risk Chem. Man 1976, 10, 245–251. [Google Scholar]
- Cui, J.; Wang, J.; Huang, S.; Jiang, X.; Li, Y.; Wu, W.; Zhang, X. Sterigmatocystin induced apoptosis in human pulmonary cells in vitro. Exp. Toxicol. Pathol. 2017, 69, 695–699. [Google Scholar] [CrossRef]
- Nieto, C.H.D.; Granero, A.M.; Zon, M.A.; Fernández, H. Sterigmatocystin: A mycotoxin to be seriously considered. Food Chem. Toxicol. 2018, 118, 460–470. [Google Scholar] [CrossRef]
- Wang, J.; Huang, S.; Xing, L.; Shen, H.; Yan, X.; Wang, J.; Zhang, X. Role of hMLH1 in sterigmatocystin-induced G2 phase arrest in human esophageal epithelial Het-1A cells in vitro. Toxicol. Lett. 2013, 217, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Sun, M.; Gilyan, A.; Roy, J.; Rand, T. Inflammation-associated gene transcription and expression in mouse lungs induced by low molecular weight compounds from fungi from the built environment. Chem. Interact. 2010, 183, 113–124. [Google Scholar] [CrossRef]
- Mori, H.; Sugie, S.; Yoshimi, N.; Kitamura, J.; Niwa, M.; Hamasaki, T.; Kawai, K. Genotoxic effects of a variety of sterigmatocystin-related compounds in the hepatocyte/DNA-repair test and the Salmonella microsome assay. Mutat. Res. Lett. 1986, 173, 217–222. [Google Scholar] [CrossRef]
- Jakšić, D.; Puel, O.; Canlet, C.; Kopjar, N.; Kosalec, I.; Šegvić Klarić, M. Cytotoxicity and genotoxicity of versicolorins and 5-methoxysterigmatocystin in A549 cells. Arch. Toxicol. 2012, 86, 1583–1591. [Google Scholar] [CrossRef]
- Kifer, D.; Jakšić, D.; Šegvić Klarić, M. Assessing the effect of mycotoxin combinations: Which mathematical model is (the most) appropriate? Toxins 2020, 12, 153. [Google Scholar] [CrossRef] [Green Version]
- Cabaret, O.; Puel, O.; Botterel, F.; Pean, M.; Khoufache, K.; Costa, J.-M.; Delaforge, M.; Bretagne, S. Metabolic detoxication pathways for sterigmatocystin in primary tracheal epithelial cells. Chem. Res. Toxicol. 2010, 23, 1673–1681. [Google Scholar] [CrossRef]
- Micheluz, A.; Sulyok, M.; Manente, S.; Krska, R.; Varese, G.; Ravagnan, G. Fungal secondary metabolite analysis applied to Cultural Heritage: The case of a contaminated library in Venice. World Mycotox. J. 2016, 9, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, A.; Henry, F.S.; Butler, J.P. Particle transport and deposition: Basic physics of particle kinetics. Compreh. Physiol. 2013, 3, 1437–1471. [Google Scholar] [CrossRef] [Green Version]
- Morawska, L.; Afshari, A.; Bae, G.N.; Buonanno, G.; Chao, C.Y.H.; Hänninen, O.; Hofmann, W.; Isaxon, C.; Jayaratne, E.R.; Pasanen, P.; et al. Indoor aerosols: From personal exposure to risk assessment. Indoor Air 2013, 23, 462–487. [Google Scholar] [CrossRef] [Green Version]
- Gravesen, S.; Nielsen, P.A.; Iversen, R.; Nielsen, K.F. Microfungal contamination of damp buildings--examples of risk constructions and risk materials. Environ. Health Perspect. 1999, 107, 505–508. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Gravesen, S.; Nielsen, P.A.; Andersen, B.; Thrane, U.; Frisvad, J.C. Production of mycotoxins on artificially and naturally infested building materials. Mycopathologia 1999, 145, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rašić, D.; Jakšić, D.; Hulina Tomašković, A.; Kifer, D.; Kopjar, N.; Rumora, L.; Želježić, D.; Peraica, M.; Šegvić Klarić, M. Sterigmatocystin moderately induces oxidative stress in male Wistar rats after short-term oral treatment. Mycotox. Res. 2019, 36, 181–191. [Google Scholar] [CrossRef]
- Liu, L.; Liu, R.; Basnet, B.B.; Bao, L.; Han, J.; Wang, L.; Liu, H. New phenolic bisabolane sesquiterpenoid derivatives with cytotoxicity from Aspergillus tennesseensis. J. Antibiot. 2018, 71, 538–542. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 2563–2582. [Google Scholar]
- Suganuma, M.; Okabe, S.; Marino, M.W.; Sakai, A.; Sueoka, E.; Fujiki, H. Essential role of tumor necrosis factor α (TNF-α) in tumor promotion as revealed by TNF-α-deficient mice. Cancer Res. 1999, 59, 4516–4518. [Google Scholar]
- Duque, G.A.; Descoteaux, A. Macrophage Cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [Green Version]
- Preedy, V.R.; Patel, V.B. (Eds.) General Methods in Biomarker Research and Their Applications; Springer: Berlin, Germany, 2015. [Google Scholar]
- Zhang, X.H.; Yao, Z.G.; Wang, J.; Xing, L.X.; Xia, Y. Effects of sterigmatocystin on TNF-? IL-6 and IL-12 expression in murine peripheral blood mononuclear cells and peritoneal macrophages in vivo. Mol. Med. Rep. 2012, 5, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Korkalainen, M.; Täubel, M.; Naarala, J.; Kirjavainen, P.; Koistinen, A.; Hyvärinen, A.; Komulainen, H.; Viluksela, M. Synergistic proinflammatory interactions of microbial toxins and structural components characteristic to moisture-damaged buildings. Indoor Air 2016, 27, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.R.; Oscoz, A.A.; Brunborg, G.; Gaivão, I.; Giovannelli, L.; Kruszewski, M.; Smith, C.C.; Štětina, R. The comet assay: Topical issues. Mutagenesis 2008, 23, 143–151. [Google Scholar] [CrossRef]
- Collins, A.R. The Comet Assay for DNA Damage and Repair: Principles, Applications, and Limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Jakšić, D.; Šegvić Klarić, M.; Crnolatac, I.; Vujičić, N.; Šijaković Smrečki, V.; Górecki, M.; Pescitelli, G.; Piantanida, I. Unique aggregation of sterigmatocystin in water yields strong and specific circular dichroism response allowing highly sensitive and selective monitoring of bio-relevant interactions. Mar. Drugs 2019, 17, 629. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Wang, J.; Xing, L.; Shen, H.; Yan, X.; Wang, J.; Zhang, X. Impairment of cell cycle progression by sterigmatocystin in human pulmonary cells in vitro. Food Chem. Toxicol. 2014, 66, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Mihaljević, Z.; Ternjej, I.; Stanković, I.; Ivković, M.; Zelježić, D.; Mladinić, M.; Kopjar, N. Assessment of genotoxic potency of sulfate-rich surface waters on medicinal leech and human leukocytes using different versions of the Comet assay. Ecotoxicol. Env. Saf. 2011, 74, 1416–1426. [Google Scholar] [CrossRef]
- Wojewódzka, M.; Gradzka, I.; Buraczewska, I. Modified neutral comet assay for human lymphocytes. Nukleonika 2002, 47, 1–5. [Google Scholar]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means. 2019. Available online: https://github.com/rvlenth/emmeans (accessed on 22 November 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakšić, D.; Ćurtović, I.; Kifer, D.; Rašić, D.; Kopjar, N.; Micek, V.; Peraica, M.; Klarić, M.Š. Single-Dose Toxicity of Individual and Combined Sterigmatocystin and 5-Methoxysterigmatocistin in Rat Lungs. Toxins 2020, 12, 734. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110734
Jakšić D, Ćurtović I, Kifer D, Rašić D, Kopjar N, Micek V, Peraica M, Klarić MŠ. Single-Dose Toxicity of Individual and Combined Sterigmatocystin and 5-Methoxysterigmatocistin in Rat Lungs. Toxins. 2020; 12(11):734. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110734
Chicago/Turabian StyleJakšić, Daniela, Ida Ćurtović, Domagoj Kifer, Dubravka Rašić, Nevenka Kopjar, Vedran Micek, Maja Peraica, and Maja Šegvić Klarić. 2020. "Single-Dose Toxicity of Individual and Combined Sterigmatocystin and 5-Methoxysterigmatocistin in Rat Lungs" Toxins 12, no. 11: 734. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12110734