Potential for Bacillus thuringiensis and Other Bacterial Toxins as Biological Control Agents to Combat Dipteran Pests of Medical and Agronomic Importance



Abstract
:1. Introduction
2. The Entomopathogenic Bacterium Bacillus thuringiensis
3. Importance of Dipteran Control
4. Dipteran-Active Insecticidal Bacterial Toxins
4.1. Bt Toxins Active against Diptera

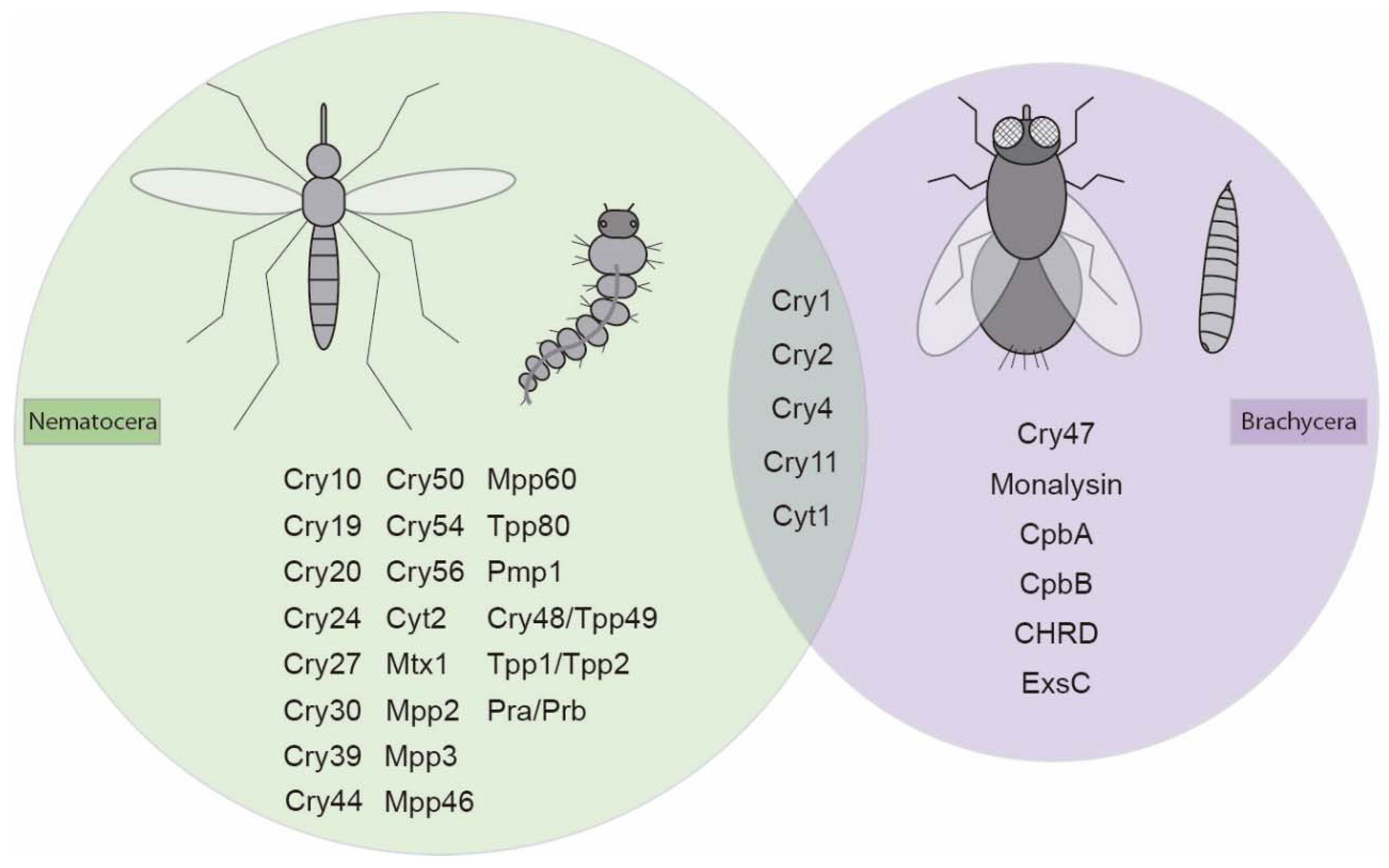{kind=link}
{kind=link}
{kind=link}
| Family | Name (Former Name) | Insect Target | Activity Range LC50 (µg/mL) | References | |
|---|---|---|---|---|---|
| Family | Species | ||||
| Cry1 | Cry1Ab7 | Culicidae | Aedes aegypti | ND a | [34] |
| Cry1Ac8 | Glossinidae | Glossina morsitans | 0.42–0.74 a | [35] | |
| Cry1Ba1 | Muscidae | Musca domestica | 20 a | [36] | |
| Calliphoridae | Lucilia cuprina | ND a,b | [37,38] | ||
| Cry1Bc1 | Muscidae | Musca domestica | 79.4 a | [37] | |
| Calliphoridae | Lucilia cuprina | 308 a | [37] | ||
| Chrysomya albiceps | 807 a | [37] | |||
| Cry1Ca1 | Culicidae | Aedes aegypti | 39.3–141 a | [39,40] | |
| Anopheles gambiae | 143–283 a | [39] | |||
| Culex quinquefasciatus | 126 a | [39] | |||
| Cry2 | Cry2Aa1 | Culicidae | Aedes aegypti | 37.06–79.46 a,b | [41,42,43,44] |
| Anopheles quadrimaculatus | 0.37 a | [44] | |||
| Aedes triseriatus (Ochlerotatus triseriatus) | 2.84 a | [44] | |||
| Culex quinquefasciatus | 0.53 a | [45] | |||
| Cry2Aa2 | Culicidae | Culex quinquefasciatus | 1.63 a | [46] | |
| Cry2Aa4 | Culicidae | Aedes aegypti | ND a | [47] | |
| Anopheles stephensi | ND a | [47] | |||
| Culex quinquefasciatus (Culex fatigans) | ND a | [47] | |||
| Cry2Aa14 | Culicidae | Culex quinquefasciatus | 0.894 a | [48] | |
| Cry2Aa | Culicidae | Anopheles gambiae | 0.11 a | [49] | |
| Cry2Ab1 | Culicidae | Aedes aegypti | 23.42–35.80 a,b | [6,43,50,51,52] | |
| Cry2Ab2 | Culicidae | Anopheles gambiae | 0.54 a | [49] | |
| Cry2Ab25 | Tephritidae | Rhagoletis cerasi | ND b | [53] | |
| Cry2Ac11 | Culicidae | Aedes aegypti | [6] | ||
| Cry2Ag | Culicidae | Aedes aegypti | 2.54 b | [51] | |
| Cry2Am1 | Culicidae | Aedes aegypti | [6] | ||
| Cry4 | Cry4Aa1 | Culicidae | Aedes aegypti | 0.03–13 a,b | [54,55,56,57,58,59,60,61,62,63] |
| Anopheles gambiae | 1.07–1.17 a | [55] | |||
| Anopheles stephensi | 0.52–7.4 a,b | [59,60,61,64] | |||
| Culex pipiens | 0.25–0.97 a | [54,59,60,61,65] | |||
| Culex quinquefasciatus | 0.05–5.04 a,b | [33,54,55,66] | |||
| Simuliidae | Simulium spp. | ND b | [67] | ||
| Cry4Ba1 | Chironomidae | Chironomus tepperi | 0.94 b | [68] | |
| Culicidae | Aedes aegypti | 0.12–0.94 a,b | [54,55,57,58,59,61,64,69] | ||
| Anopheles albimanus | 1.3 b | [70] | |||
| Anopheles gambiae | 0.79 a | [55] | |||
| Anopheles quadrimaculatus | 0.25 a | [54] | |||
| Anopheles stephensi | 0.55–17 a,b | [59,61,64] | |||
| Culex quinquefasciatus | 24.5 a | [55] | |||
| Culicidae | Culex pipiens | ND b | [64] | ||
| Tipulidae | Tipula oleracea | ND a | [71] | ||
| Simuliidae | Simulium spp. | ND b | [67] | ||
| Cry4Ba2 | Culicidae | Aedes aegypti | ND a | [72] | |
| Cry4Cb1 | Culicidae | Aedes aegypti | 0.083 b | [73] | |
| Cry10 | Cry10Aa | Culicidae | Aedes aegypti | 0.3–20.61 a,b | [62,74] |
| Cry11 | Cry11Aa1 | Chironomidae | Chironomus tepperi | 0.56 b | [68] |
| Culicidae | Aedes aegypti | 0.01-1.35 a,b | [41,42,57,58,61,75,76,77,78,79] | ||
| Anopheles albimanus | 0.9 b | [70] | |||
| Anopheles stephensi | 0.13-0.45 a,b | [60,75,78,80,81] | |||
| Anopheles albimanus | 6.759 a | [77] | |||
| Culex pipiens | 0.009-0.27 a | [75,78] | |||
| Culex quinquefasciatus | 0.01-0.13 a,b | [45,66,76,77,82,83] | |||
| Tipulidae | Tipula oleracea | ND b | [81] | ||
| Simuliidae | Simulium spp. | ND b | [67] | ||
| Cry11Ba1 | Culicidae | Aedes aegypti | 0.02–0.03 a,b | [75,77,84] | |
| Anopheles albimanus | 0.10 a | [77] | |||
| Anopheles stephensi | 0.04 a | [75] | |||
| Culex pipiens | 0.01–0.11 a | [61,65] | |||
| Culex quinquefasciatus | 0.006–0.02 a,b | [77,83,84] | |||
| Cry11Bb1 | Culicidae | Aedes aegypti | 0.02–0.85 a,b | [76,77,85,86] | |
| Anopheles albimanus | 0.17 a | [77] | |||
| Anopheles stephensi | 0.07 a | [85] | |||
| Culex pipiens | 0.04 a | [85] | |||
| Culex quinquefasciatus | 0.01–0.13 a,b | [76,77,86] | |||
| Cry16-Cry17-Cbm17.1-Cbm17.2 | Cry16-Cry17-Cbm17.1-Cbm17.2 | Culicidae | Aedes aegypti | ND b | [87] |
| Cry19 | Cry19Aa | Culicidae | Anopheles stephensi | 1.04 a | [88] |
| Culex pipiens | 0.19 a | [88] | |||
| Cry19B | Culicidae | Culex pipiens molestus | 5.93 a | [89] | |
| Cry20 | Cry20Aa1 | Culicidae | Aedes aegypti | 648 b | [90] |
| Culex quinquefasciatus | 700 b | [90] | |||
| Cry24 | Cry24Ca1 | Culicidae | Aedes aegypti | 0.48 b | [91] |
| Cry27 | Cry27Aa1 | Culicidae | Anopheles stephensi | 94.3 a | [92] |
| Cry30 | Cry30Fa1 | Culicidae | Aedes aegypti | 15.4 a | [93] |
| Cry30Ga1 | Culicidae | Aedes aegypti | 7.10 b | [73] | |
| Cry39 | Cry39Aa1 | Culicidae | Anopheles stephensi | 0.75 b | [94] |
| Culex pipiens | 41.94 b | [95] | |||
| Cry44 | Cry44Aa1 | Culicidae | Aedes aegypti | 0.01 a | [60] |
| Anopheles stephensi | 1.26 a | [60] | |||
| Culex pipiens | 0.006 a | [60] | |||
| Mpp46 | Mpp46Ab (Cry46Ab) | Culicidae | Culex pipiens | 1.02 a | [96] |
| Cry47 | Cry47Aa1 | Calliphoridae | Lucilia cuprina | ND a | [97,98] |
| Cry50 | Cry50Ba | Culicidae | Culex quinquefasciatus | 0.07 a | [99] |
| Cry54 | Cry54Aa1 | Culicidae | Aedes aegypti | 9.02 a | [100] |
| Cry56 | Cry56Aa1 | Culicidae | Aedes aegypti | 0.15 a | [101] |
| Mpp60 | Mpp60Aa (Cry60Aa) | Culicidae | Culex quinquefasciatus | 7.9 b | [102] |
| Mpp60Ba (Cry60Ba) | Culicidae | Culex quinquefasciatus | 5.5 b | [102] | |
| Mpp60Aa+Mpp60Ba (Cry60Aa+Cry60Ba) | Culicidae | Culex quinquefasciatus | 2.9 b | [102] | |
| Tpp80 | Tpp80Aa1 (Cry80Aa1) | Culicidae | Culex pipiens pallens | 71.9 a | [103] |
| Cyt1 | Cyt1Aa1 | Calliphoridae | Calliphora stygia | 305 a | [104] |
| Lucilia cuprina | 296 a | [104] | |||
| Lucilia sericata | 236 a | [104] | |||
| Chironomidae | Chironomus tepperi | 31 b | [68] | ||
| Culicidae | Aedes aegypti | 0.15–1.86 a,b | [74,105,106,107,108,109] | ||
| Anopheles stephensi | 2.7–6.3 a | [105,108] | |||
| Culex pipiens | 0.6–1.2 a | [105,108] | |||
| Culex quinquefasciatus | 0.4 a | [108] | |||
| Tephritidae | Ceratitis capitata | ND a | [26] | ||
| Tipulidae | Tipula paludosa | ND a | [110] | ||
| Cyt1Aa2 | Culicidae | Aedes aegypti | 0.12–1.21 a,b | [58,63,111] | |
| Anopheles gambiae | 1–2 a,b | [111] | |||
| Culex pipiens | 0.5–2 a,b | [111] | |||
| Cyt1Aa4 | Culicidae | Aedes aegypti | 0.06 a | [79] | |
| Culex quinquefasciatus | >10 a | [112] | |||
| Cyt1Ab1 | Culicidae | Aedes aegypti | 32.6–59 a,b | [105,113] | |
| Anopheles stephensi | 20 a | [105] | |||
| Culex pipiens | 5.7 a | [105] | |||
| Culex quinquefasciatus | 32.9–114.5 b | [113] | |||
| Cyt1Ba1 | Agromyzidae | Liriomyza trifolii | ND a | [114] | |
| Cyt2 | Cyt2Aa1 | Culicidae | Aedes aegypti | 1–4 a,b | [111] |
| Anopheles gambiae | 1–2 a,b | [111] | |||
| Culex pipiens | 0.5–4 a,b | [111] | |||
| Cyt2Aa2 | Culicidae | Aedes aegypti | 0.35–0.5 a | [69,115] | |
| Culex quinquefasciatus | 0.25–0.5 a | [69,115] | |||
| Cyt2Aa3 | Chironomidae | Chironomus tepperi | 36 a | [116] | |
| Culicidae | Culex quinquefasciatus | 0.53 a | [116] | ||
| Cyt2Ba1 | Culicidae | Aedes aegypti | 0.28–33 a,b | [62,108,113,117] | |
| Anopheles stephensi | 5.5 a | [108] | |||
| Culex pipiens | 5 a | [108] | |||
| Culex quinquefasciatus | 1.8–31.5 a,b | [108,113] | |||
| Cyt2Bb1 | Culicidae | Aedes aegypti | 6.8 b | [106] | |
| Cyt2Bc1 | Culicidae | Aedes aegypti | 7 a | [108] | |
| Anopheles stephensi | 11 a | [108] | |||
| Culex pipiens | 7.3 a | [108] | |||
| Culex quinquefasciatus | 1.8 a | [108] | |||
| Mtx | Mtx1Aa1 | Culicidae | Culex quinquefasciatus | 0.01 a | [118] |
| Aedes aegypti | 0.05 a | [118] | |||
| Chironomidae | Chironomus riparius | 4.06 a | [119] | ||
| Mpp2 | Mpp2Aa1 (Mtx2) | Culicidae | Culex quinquefasciatus | 4.13–107 b | [66] |
| Mpp2Aa1 (Mtx2 strain SSII-1) | Culicidae | Culex quinquefasciatus | 0.93 a | [120] | |
| Aedes aegypti | 14.5 a | [120] | |||
| Mpp2Aa2 Mtx2 (31-2) | Culicidae | Culex quinquefasciatus | 3.90 a | [120] | |
| Aedes aegypti | 3.91 a | [120] | |||
| Mpp3 | Mpp3Aa1 (Mtx3) | Culicidae | Culex quinquefasciatus | ND a | [121] |
| Aedes aegypti | ND a | [121] | |||
| Monalysin | Monalysin | Drosophilidae | Drosophila melanogaster | ND a | [122] |
| CpbA | CpbA | Muscidae | Musca domestica | ND a | [123] |
| CpbB | CpbB | Muscidae | Musca domestica | ND a | [123] |
| CHRD | CHRD | Muscidae | Musca domestica | ND a | [123] |
| ExsC | ExsC | Muscidae | Musca domestica | ND a | [123] |
| Pmp1 | Pmp1 | Culicidae | Anopheles coluzzii | ND a | [124] |
| Two part toxins | Cry48Aa/Tpp49 (Cry49Aa) | Culicidae | Culex quinquefasciatus | 0.02/0.006 a | [125] |
| Tpp1/Tpp2 (BinA/BinB) | Culicidae | Aedes aegypti | 42 a | [126,127] | |
| Aedes atropalpus | ND a | [126] | |||
| Anopheles gambiae | 0.36 a | [127] | |||
| Anopheles stephensi | 0.39 a | [127] | |||
| Anopheles albimanus | 1 a | [127] | |||
| Anopheles quadrimaculatus | 4.6 a | [127] | |||
| Culex pipiens | 0.1 a | [127] | |||
| Culicidae | Culex quinquefasciatus | 0.013–0.03 a,b | [126,128] | ||
| Pra/Prb (PirA/PirB) | Culicidae | Aedes aegypti | ND a | [84,129] | |
| Synergy | Cry1Ca/Cyt1Aa | Culicidae | Aedes aegypti | 0.61 a | [130] |
| Cry2Aa/Cry2Ab | Culicidae | Aedes aegypti | 51.3 b | [43] | |
| Cry2Aa/Cry50Ba | Culicidae | Culex quinquefasciatus | 0.05 a | [99] | |
| Cry4Aa/Cry4Ba | Culicidae | Aedes aegypti | 0.05 a | [61] | |
| Anopheles stephensi | 0.02 a | [61] | |||
| Culex pipiens | 0.04 a | [61] | |||
| Culex quinquefasciatus | 1.49–315 b | [66] | |||
| Simuliidae | Simulium spp. | ND b | [67] | ||
| Cry4Aa/Cyt1Aa | Culicidae | Aedes aegypti | 0.07 a | [58] | |
| Chironomidae | Chironomus tepperi | 44 b | [68] | ||
| Tipulidae | Tipula paludosa | ND a | [110] | ||
| Cry4Aa/Cyt2Ba | Culicidae | Aedes aegypti | 0.013 b | [62,131] | |
| Cry4Aa/Cry11Ba | Culicidae | Culex pipiens | 0.04 a | [65] | |
| Cry4Aa/Cry46Ab | Culicidae | Culex pipiens | 0.18 a | [96] | |
| Cry4A/Mtx1Aa1 | Culicidae | Culex quinquefasciatus | 1.06–2.37 b | [66] | |
| Cry4A/Mpp2Aa1 (Mtx2) | Culicidae | Culex quinquefasciatus | 0.27–1.21 b | [66] | |
| Cry4Ba/Cry11Aa | Culicidae | Anopheles albimanus | 0.567 b | [70] | |
| Simuliidae | Simulium spp. | ND b | [67] | ||
| Cry4Ba/Cyt1Aa | Culicidae | Aedes aegypti | 0.62 a | [58] | |
| Culicidae | Anopheles albimanus | 0.33–0.77 b | [70] | ||
| Cry4Ba/Cyt2Aa2 | Culicidae | Aedes aegypti | 0.007 a | [69] | |
| Culex quinquefasciatus | 0.02 a | [69] | |||
| Cry4B/Mtx1Aa1 | Culicidae | Culex quinquefasciatus | 18.2–29.0 b | [66] | |
| Cry4B/Mpp2Aa1 (Mtx2) | Culicidae | Culex quinquefasciatus | 85.7 b | [66] | |
| Cry10Aa/Cyt1Aa | Culicidae | Aedes aegypti | 0.03–0.08 a,b | [74] | |
| Cry10Aa/Cyt2Ba | Culicidae | Aedes aegypti | 0.004 b | [62] | |
| Cry11Aa/Cyt1Aa | Culicidae | Aedes aegypti | 0.01–0.12 a | [58,79] | |
| Culicidae | Anopheles albimanus | 0.28–0.37 b | [70] | ||
| Cry11/Mtx1Aa1 | Culicidae | Culex quinquefasciatus | 0.66–3.03 b | [66] | |
| Cry11/Mpp2Aa1 (Mtx2) | Culex quinquefasciatus | 0.90 b | [66] | ||
| Cry11Bb/Cry29Aa | Culicidae | Aedes aegypti | 3.94 a | [85] | |
| Anopheles stephensi | 2.13 a | [85] | |||
| Culex pipiens | 0.73 a | [85] | |||
| Cry11Bb/Cry30Aa | Culicidae | Aedes aegypti | 16.96 a | [85] | |
| Anopheles stephensi | 1.43 a | [85] | |||
| Culex pipiens | 1.13 a | [85] | |||
| Cry4A/Cry4B/Cry11A | Culicidae | Aedes aegypti | 0.12 a | [58] | |
| Culex quinquefasciatus | 0.008–0.59 b | [66] | |||
| Cry4A/Cry4B/Cyt1A | Culicidae | Aedes aegypti | 0.08 a | [58] | |
| Cry4A/Cry4B/Mtx1Aa1 | Culicidae | Culex quinquefasciatus | 0.18–0.77 b | [66] | |
| Cry4A/Cry4B/Mpp2Aa1 (Mtx2) | Culicidae | Culex quinquefasciatus | 0.11–0.32 b | [66] | |
| Cry4A/Cry4B/Cry11A/Cyt1A | Culicidae | Aedes aegypti | 0.08 a | [58] | |
| Culex quinquefasciatus | 0.02–0.07 b | [66] | |||
| Cry4A/Cry4B/Cry11A/Mtx1Aa1 | Culicidae | Culex quinquefasciatus | 0.02–0.24 b | [66] | |
| Cry4A/Cry4B/Cry11A/Mpp2Aa1 (Mtx2) | Culicidae | Culex quinquefasciatus | 0.03–0.06 b | [66] | |
| Cry4A/Cry4B/Cry11A/Cyt1A/Mtx1Aa1 | Culicidae | Culex quinquefasciatus | 0.02–0.06 b | [66] | |
| Cry4A/Cry4B/Cry11A/Cyt1A/Mpp2Aa1(Mtx2) | Culicidae | Culex quinquefasciatus | 0.30–1.09 b | [66] | |
| Cry4Ba/Cry11Aa/Cyt1Aa | Culicidae | Anopheles albimanus | 0.7–8.33 b | [70] | |
| Cry11Bb/Cry29Aa/Cry30Aa | Culicidae | Aedes aegypti | 5.43 a | [85] | |
| Anopheles stephensi | 1.31 a | [85] | |||
| Culex pipiens | 0.85 a | [85] | |||
4.1.1. Cry Toxins from Bt ser. israelensis
Cry4 Proteins
Cry10Aa Protein
Cry11 Proteins
Mpp60A/Mpp60B Proteins (Formerly Cry60A/Cry60B)
4.1.2. Other Toxins Specific to Diptera
4.1.3. Anti-Dipteran Toxins with Cross-Order Activity
Cry1 Protein
Cry2A Protein
Other Cry Proteins
4.1.4. Anti-Dipteran Cyt Toxins
Cyt1 Proteins
Cyt2 Proteins
4.2. Anti-Dipteran Toxins from Other Microorganisms
4.2.1. Toxins from Lysinibacillus sphaericus
4.2.2. Toxins from Paraclostridium bifermentans
4.2.3. Other Dipteran-Active Proteins
5. Toxins with Synergistic Activity against Diptera
6. Bacterial Insecticides against Mosquitoes
7. Concluding Remarks and Future Perspectives
Funding
Conflicts of Interest
References
- Martin, P.A.W.; Travers, R.S. Worldwide abundance and distribution of Bacillus thuringiensis isolates. Appl. Environ. Microbiol. 1989, 55, 2437–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iriarte, J.; Bel, Y.; Ferrandis, M.D.; Andrew, R.; Murillo, J.; Ferré, J.; Caballero, P. Environmental distribution and diversity of Bacillus thuringiensis in Spain. Syst. Appl. Microbiol. 1998, 21, 97–106. [Google Scholar] [CrossRef]
- Jouzani, G.S.; Valijanian, E.; Sharafi, R. Bacillus thuringiensis: A successful insecticide with new environmental features and tidings. Appl. Microbiol. Biotechnol. 2017, 101, 2691–2711. [Google Scholar] [CrossRef]
- Caballero, P.; Ferré, J. Bioinsecticidas: Fundamentos y Aplicaciones de Bacillus Thuringiensis en el Control Integrado de Plagas; PHYTOMA-España en colaboración con la, Universidad Pública de Navarra, Ed.; Phytoma: Valencia, Spain, 2001. [Google Scholar]
- Federici, B.A. Bacillus thuringiensis. In Handbook of Biological Control; Bellows, T., Gordh, G., Fisher, T., Eds.; Academic Press: San Diego, CA, USA, 1999. [Google Scholar]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2020, 107438. [Google Scholar] [CrossRef] [PubMed]
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.-Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.-C.; Aroian, R. Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, A.; Sarjeet, G.; Mario, S. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Natl. Inst. Acess 2007, 49, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberon, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, J.J.; Dulmage, H.T.; Carlton, B. Correlation between specific plasmids and delta-endotoxin production in Bacillus thuringiensis. Plasmid 1981, 5, 352–365. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Jackson, T.A. Bacterial entomopathogens; Academic Press: Cambridge, MA, USA, 2012; ISBN 9780123849847. [Google Scholar]
- Agaisse, H.; Lereclus, D. How does Bacillus thuringiensis produce so much insecticidal crystal protein? J. Bacteriol. 1995, 177, 6027–6032. [Google Scholar] [CrossRef] [Green Version]
- De Maagd, R.A.; Bravo, A.; Berry, C.; Crickmore, N.; Schnepf, H.E. Structure, diversity, and evolution of protein toxins from spore-forming entomopathogenic bacteria. Annu. Rev. Genet. 2003, 37, 409–433. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, B.C.; Yu, Z.; Sun, M. Structural Insights into Bacillus thuringiensis Cry, Cyt and Parasporin Toxins. Toxins 2014, 6, 2732–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Maagd, R.A.; Bosch, D.; Stiekema, W. Bacillus thuringiensis toxin-mediated insect resistance in plants. Trends Plant Sci. 1999, 4, 9–13. [Google Scholar] [CrossRef]
- Bravo, A.; Gómez, I.; Porta, H.; García-Gómez, B.I.; Rodriguez-Almazan, C.; Pardo, L.; Soberón, M. Evolution of Bacillus thuringiensis Cry toxins insecticidal activity. Microb. Biotechnol. 2013, 6, 17–26. [Google Scholar] [CrossRef]
- De Maagd, R.A.; Bravo, A.; Crickmore, N. How Bacillus thuringiensis has evolved specific toxins to colonize the insect world. Trends Genet. 2001, 17, 193–199. [Google Scholar] [CrossRef]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Berry, C.; Crickmore, N. Structural classification of insecticidal proteins—Towards an in silico characterisation of novel toxins. J. Invertebr. Pathol. 2017, 142, 16–22. [Google Scholar] [CrossRef]
- Butko, P. Cytolytic toxin Cyt1A and its mechanism of membrane damage: Data and hypotheses. Appl. Environ. Microbiol. 2003, 69, 2415–2422. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.; Albeck, S.; Ben-Dov, E.; Cahan, R.; Firer, M.; Zaritsky, A.; Dym, O. Cyt1Aa toxin: Crystal structure reveals implications for its membrane-perforating function. J. Mol. Biol. 2011, 413, 804–814. [Google Scholar] [CrossRef]
- Cohen, S.; Dym, O.; Albeck, S.; Ben-Dov, E.; Cahan, R.; Firer, M.; Zaritsky, A. High-Resolution crystal structure of activated Cyt2Ba monomer from Bacillus thuringiensis subsp. Isr. J. Mol. Biol. 2008, 380, 820–827. [Google Scholar] [CrossRef]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identifiction and Bionomics; CAB Intern.: Wallingford, UK, 1992. [Google Scholar]
- Wheeler, M.R. Additions to the catalog of the world’s Drosophilidae. In The Genetics and Biology of Drosophila; Ashburner, M., Thompson, J.N., Carson, H.L., Eds.; Academic Press: London, UK, 1986; pp. 1–105. [Google Scholar]
- Vidal-Quist, J.C.; Castañera, P.; González-Cabrera, J. Cyt1Aa protein from Bacillus thuringiensis (Berliner) serovar israelensis is active against the Mediterranean fruit fly, Ceratitis capitata (Wiedemann). Pest Manag. Sci. 2010, 66, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Leibee, G.L. Influence of temperature on development and fecundity of Liriomyza trifolii (Burgess) (Diptera: Agromyzidae) on Celery. Environ. Entomol. 1984, 13, 497–501. [Google Scholar] [CrossRef]
- Minkenberg, O.P.J.M. Dispersal of Liriomyza trifolii. EPPO Bull. 1988, 18, 173–182. [Google Scholar] [CrossRef]
- McPheron, B.A.; Steck, G.J. Fruit Fly Pests: A World Assessment of Their Biology and Management; Taylor & Francis Inc., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1996; ISBN 1574440144. [Google Scholar]
- EPPO A2 List EPPO. European and Mediterranean Plant Protection Organization, A2 List of pests recommended for regulation as quarantine pests. Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/A2_list (accessed on 1 December 2020).
- WHO. Available online: http://www.who.int/malaria (accessed on 4 December 2020).
- CDC. Available online: https://www.cdc.gov/ (accessed on 4 December 2020).
- Zhang, Q.; Hua, G.; Adang, M.J. Effects and mechanisms of Bacillus thuringiensis crystal toxins for mosquito larvae. Insect Sci. 2017, 24, 714–729. [Google Scholar] [CrossRef]
- Haider, M.; Ward, E.S.; Ellar, D.J. Cloning and heterologous expression of an insecticidal delta-endotoxin gene from Bacillus thuringiensis var. aizawai ICI toxic to both lepidoptera and diptera. Gene 1987, 52, 285–290. [Google Scholar] [CrossRef]
- Omolo, E.O.; James, M.D.; Osir, E.O.; Thomson, J.A. Cloning and expression of a Bacillus thuringiensis (L1-2) gene encoding a crystal protein active against Glossina morsitans morsitans and Chilo partellus. Curr. Microbiol. 1997, 34, 118–121. [Google Scholar] [CrossRef]
- Zhong, C.; Ellar, D.J.; Bishop, A.; Johnson, C.; Lin, S.; Hart, E.R. Characterization of a Bacillus thuringiensis δ-endotoxin which is toxic to insects in three orders. J. Invertebr. Pathol. 2000, 76, 131–139. [Google Scholar] [CrossRef]
- Johnson, C.; Bishop, A.H.; Turner, C.L. Isolation and activity of strains of Bacillus thuringiensis toxic to larvae of the housefly (Diptera: Muscidae) and tropical blowflies (Diptera: Calliphoridae). J. Invertebr. Pathol. 1998, 71, 138–144. [Google Scholar] [CrossRef]
- Heath, A.C.G.; Wigley, P.J.; Shoemaker, C.B.; Chilcott, C.N.; Broadwell, A.H. Efficacy of native and recombinant Cry1B protein against experimentally induced and naturally acquired ovine myiasis (fly strike) in sheep. J. Econ. Entomol. 2004, 97, 1797–1804. [Google Scholar] [CrossRef]
- Smith, G.P.; Merrick, J.D.; Bone, E.J.; Ellar, D.J. Mosquitocidal activity of the CryIC δ-endotoxin from Bacillus thuringiensis subsp. aizawai. Appl. Environ. Microbiol. 1996, 62, 680–684. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Rauf, M.; Ellar, D.J. Toxicity and receptor binding properties of a Bacillus thuringiensis CryIC toxin active against both Lepidoptera and Diptera. J. Invertebr. Pathol. 1999, 73, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Donovan, W.P.; Dankocsik, C.; Gilbert, M.P. Molecular characterization of a gene encoding a 72-kilodalton mosquito-toxic crystal protein from Bacillus thuringiensis subsp. israelensis. J. Bacteriol. 1988, 170, 4732–4738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.W.; Bideshi, D.K.; Johnson, J.J.; Federici, B.A. Differential enhancement of Cry2A versus Cry11A yields in Bacillus thuringiensis by use of the cry3A STAB mRNA sequence. FEMS Microbiol. Lett. 1999, 181, 319–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricoldi, M.C.; Soares Figueiredo, C.; Apparecida Desiderio, J. Toxicity of Cry2 proteins from Bacillus thuringiensis subsp. thuringiensis strain T01-328 against Aedes aegypti (Diptera: Culicidae). Toxicology 2018, 85, 1–7. [Google Scholar] [CrossRef]
- Sims, S.R. Host activity spectrum of the CryIIA Bacillus thuringiensis subsp. kurstaki protein: Effects on Lepidoptera, Diptera, and non-target arthropods. Southwest. Entomol. 1997, 22, 395–404. [Google Scholar]
- Zhang, L.; Zhao, G.; Hu, X.; Liu, J.; Li, M.; Batool, K.; Chen, M.; Wang, J.; Xu, J.; Huang, T.; et al. Cry11Aa interacts with the ATP-Binding protein from Culex quinquefasciatus to improve the toxicity. J. Agric. Food Chem. 2017, 65, 10884–10890. [Google Scholar] [CrossRef]
- Moar, W.J.; Trumble, J.T.; Hice, R.H.; Backman, P.A. Insecticidal activity of the CryIIA protein from the NRD-12 isolate of Bacillus thuringiensis subsp. kurstaki expressed in Escherichia coli and Bacillus thuringiensis and in a leaf-colonizing strain of Bacillus cereus. Appl. Environ. Microbiol. 1994, 60, 896–902. [Google Scholar] [CrossRef] [Green Version]
- Misra, H.S.; Hire, R.S.; Mahajan, S.K. Cloning and characterization of an insecticidal crystal protein gene from Bacillus thuringiensis subspecies kenyae. J. Genet. 2002, 81, 5–11. [Google Scholar] [CrossRef]
- Hire, R.S.; Makde, R.D.; Dongre, T.K.; Souza, S.F.D. Expression, purification and characterization of the Cry2Aa14 toxin from Bacillus thuringiensis subsp. kenyae. Toxicon 2009, 54, 519–524. [Google Scholar] [CrossRef]
- Mcneil, B.C.; Dean, D.H. Bacillus thuringiensis Cry2Ab is active on Anopheles mosquitoes: Single D block exchanges reveal critical residues involved in activity. FEMS Microbiol. Lett. 2011, 325, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, W.; Nicholls, C.; Ellar, D.J. Cloning and expression of an entomocidal protein gene from Bacillus thuringiensis galleriae toxic to both lepidoptera and diptera. FEMS Microbiol. Lett. 1989, 59, 197–201. [Google Scholar] [CrossRef]
- Liang, H.; Liu, Y.; Zhu, J.; Guan, P.; Li, S.; Wang, S.; Zheng, A.; Liu, H.; Li, P. Characterization of Cry2-type genes of Bacillus thuringiensis strains from soil-isolated of sichuan basin, China. Braz. J. Microbiol. 2011, 42, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Widner, W.R.; Whiteley, H.R. Two highly related insecticidal crystal proteins of Bacillus thuringiensis subsp. kurstaki possess different host range specificities. J. Bacteriol. 1989, 171, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Sevim, A.; Eryüzlü, E.; Demirba, Z.; Demir, I. A novel cry2Ab gene from the indigenous isolate Bacillus thuringiensis subsp. kurstaki. J. Microbiol. Biotechnol. 2012, 22, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, M.A.F.; Alzate, O.; Mohammad, M.; McNall, R.J.; Adang, M.J.; Dean, D.H. Introduction of Culex toxicity into Bacillus thuringiensis Cry4Ba by protein engineering. Appl. Environ. Microbiol. 2003, 69, 5343–5353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angsuthanasombat, C.; Crickmore, N.; Ellar, D.J. Comparison of Bacillus thuringiensis subsp. israelensis CryIVA and CryIVB cloned toxins reveals synergism in vivo. FEMS Microbiol. Lett. 1992, 94, 63–68. [Google Scholar] [CrossRef]
- Angsuthanasombat, C.; Crickmore, N.; Ellar, D.J. Cytotoxicity of a cloned Bacillus thuringiensis subsp. israelensis CryIVB toxin to an Aedes aegypti cell line. FEMS Microbiol. Lett. 1991, 67, 273–276. [Google Scholar] [CrossRef]
- Beltrão, H.; Silva-Filha, M.H.N.L. Interaction of Bacillus thuringiensis svar. israelensis Cry toxins with binding sites from Aedes aegypti (Diptera: Culicidae) larvae midgut. FEMS Microbiol. Lett. 2007, 266, 163–169. [Google Scholar]
- Crickmore, N.; Bone, E.J.; Williams, J.A.; Ellar, D.J. Contribution of the individual components of the δ-endotoxin crystal to the mosquitocidal activity of Bacillus thuringiensis subsp. israelensis. FEMS Microbiol. Lett. 1995, 131, 249–254. [Google Scholar] [CrossRef]
- Delecluse, A.; Poncet, S.; Klier, A.; Rapoport, G. Expression of cryIVA and cryIVB genes, independently or in combination, in a crystal-negative strain of Bacillus thuningiensis subsp. israelensis. Appl. Environ. Microbiol. 1993, 59, 3922–3927. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Ikeya, T.; Sahara, K.; Bando, H.; Asano, S.I. Cloning and expression of two crystal protein genes, cry30Ba1 and cry44Aa1, obtained from a highly mosquitocidal strain, Bacillus thuringiensis subsp. entomocidus INA288. Appl. Environ. Microbiol. 2006, 72, 5673–5676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poncet, S.; Delecluse, A.; Klier, A.; Rapoport, G. Evaluation of synergistic interactions among CryIVA, CryIVB, and CryIVD toxic components of B. thuringiensis subsp. israelensis crystals. J. Invertebr. Pathol. 1995, 66, 131–135. [Google Scholar] [CrossRef]
- Valtierra-de-Luis, D.; Villanueva, M.; Lai, L.; Williams, T.; Caballero, P. Potential of Cry10Aa and Cyt2Ba, two minority δ-endotoxins produced by Bacillus thuringiensis ser. israelensis, for the control of Aedes aegypti larvae. Toxins 2020, 12, 355. [Google Scholar] [CrossRef] [PubMed]
- Ward, E.S.; Ridley, A.R.; Ellar, D.J.; Todd, J.A. Bacillus thuringiensis var. israelensis δ-endotoxin. Cloning and expression of the toxin in sporogenic and asporogenic strains of Bacillus subtilis. J. Mol. Biol. 1986, 191, 13–22. [Google Scholar] [PubMed]
- Delécluse, A.; Bourgouin, C.; Klier, A.; Rapoport, G. Specificity of action on mosquito larvae of Bacillus thuringiensis israelensis toxins encoded by two different genes. MGG Mol. Gen. Genet. 1988, 214, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Yoneda, N.; Okada, K.; Higaki, A.; Howlader, M.T.H.; Ide, T. Bacillus thuringiensis Cry11Ba works synergistically with Cry4Aa but not with Cry11Aa for toxicity against mosquito Culex pipiens (Diptera: Culicidae) larvae. Appl. Entomol. Zool. 2017. [Google Scholar] [CrossRef] [Green Version]
- Wirth, M.C.; Berry, C.; Walton, W.E.; Federici, B.A. Mtx toxins from Lysinibacillus sphaericus enhance mosquitocidal cry-toxin activity and suppress cry-resistance in Culex quinquefasciatus. J. Invertebr. Pathol. 2014, 115, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Monnerat, R.; Pereira, E.; Teles, B.; Martins, E.; Praça, L.; Queiroz, P.; Soberon, M.; Bravo, A.; Ramos, F.; Soares, C.M. Synergistic activity of Bacillus thuringiensis toxins against Simulium spp. larvae. J. Invertebr. Pathol. 2014, 121, 70–73. [Google Scholar] [CrossRef]
- Hughes, P.A.; Stevens, M.M.; Park, H.W.; Federici, B.A.; Dennis, E.S.; Akhurst, R. Response of larval Chironomus tepperi (Diptera: Chironomidae) to individual Bacillus thuringiensis var. israelensis toxins and toxin mixtures. J. Invertebr. Pathol. 2005, 88, 34–39. [Google Scholar] [CrossRef]
- Promdonkoy, B.; Promdonkoy, P.; Panyim, S. Co-expression of Bacillus thuringiensis Cry4Ba and Cyt2Aa2 in Escherichia coli revealed high synergism against Aedes aegypti and Culex quinquefasciatus larvae. FEMS Microbiol. Lett. 2005, 252, 121–126. [Google Scholar] [CrossRef]
- Fernandez-Luna, M.T.; Lanz-mendoza, H.; Gill, S.S.; Bravo, A.; Soberon, M.; Miranda-rios, J. An α-amylase is a novel receptor for Bacillus thuringiensis ssp. israelensis Cry4Ba and Cry11Aa toxins in the malaria vector mosquito Anopheles albimanus (Diptera: Culicidae). Environ. Microbiol. 2010, 12, 746–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waalwijk, C.; Dullemans, A.; Wiegers, G.; Smits, P. Toxicity of Bacillus thuringiensis variety israelensis against tipulid larvae. J. Appl. Entomol. 1992, 114, 415–420. [Google Scholar] [CrossRef]
- Angsuthanasombat, C.; Panyim, S. Biosynthesis of 130-kilodalton mosquito larvicide in the cyanobacterium Agmenellum quadruplicatum PR-6. Appl. Environ. Microbiol. 1989, 55, 2428–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Zheng, A.; Wang, S.; Liu, H.; Li, P. Characterization and expression of cry4Cb1 and cry30Ga1 from Bacillus thuringiensis strain HS18-1. J. Invertebr. Pathol. 2010, 103, 200–202. [Google Scholar] [CrossRef]
- Hernández-Soto, A.; Del Rincón-Castro, M.C.; Espinoza, A.M.; Ibarra, J.E. Parasporal body formation via overexpression of the Cry10Aa toxin of Bacillus thuringiensis subsp. israelensis, and Cry10Aa-Cyt1Aa synergism. Appl. Environ. Microbiol. 2009, 75, 4661–4667. [Google Scholar]
- Delecluse, A.; Rosso, M.L.; Ragni, A. Cloning and expression of a novel toxin gene from Bacillus thuringiensis subsp. jegathesan encoding a highly mosquitocidal protein. Appl. Environ. Microbiol. 1995, 61, 4230–4235. [Google Scholar] [CrossRef] [Green Version]
- Florez, A.M.; Suarez-Barrera, M.O.; Morales, G.M.; Rivera, K.V.; Orduz, S.; Ochoa, R.; Guerra, D.; Muskus, C. Toxic activity, molecular modeling and docking simulations of Bacillus thuringiensis Cry11 toxin variants obtained via DNA shuffling. Front. Microbiol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Orduz, S.; Realpe, M.; Arango, R.; Murillo, L.A.; Delecluse, A. Sequence of the cry11Bb11 gene from Bacillus thuringiensis subsp. medellin and toxicity analysis of its encoded protein. Biochim. Biophys. Acta 1998, 54, 267–272. [Google Scholar] [CrossRef]
- Revina, L.P.; Kostina, L.I.; Ganushkina, L.A.; Mikhailova, A.L.; Zalunin, I.A.; Chestukhina, G.G. Reconstruction of Bacillus thuringiensis ssp. israelensis Cry11A endotoxin from fragments corresponding to its N- and C-moieties restores its original biological activity. Biochemistry 2004, 69, 181–187. [Google Scholar] [CrossRef]
- Wu, D.; Johnson, J.J.; Federici, B.A. Synergism of mosquitocidal toxicity between CytA and CrylVD proteins using inclusions produced from cloned genes of Bacillus thuringiensis. Mol. Microbiol. 1994, 13, 965–972. [Google Scholar] [CrossRef]
- Bukhari, D.A.A.; Shakoori, A.R. Cloning and expression of Bacillus thuringiensis cry11 crystal protein gene in Escherichia coli. Mol. Biol. Rep. 2009, 36, 1661–1670. [Google Scholar] [CrossRef]
- Feldmann, F.; Dullemans, A.; Waalwijk, C. Binding of the CryIVD toxin of Bacillus thuringiensis subsp. israelensis to larval dipteran midgut proteins. Appl. Environ. Microbiol. 1995, 61, 2601–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.; Dai, S.M.; Frutos, R.; Federici, B.A.; Gill, S.S. Properties of a 72-kilodalton mosquitocidal protein from Bacillus thuringiensis subsp. morrisoni PG-14 expressed in B. thuringiensis subsp. kurstaki by using the shuttle vector pHT3101. Appl. Environ. Microbiol. 1992, 58, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Cheong, H.; Dhesi, R.K.; Gill, S.S. Marginal cross-resistance to mosquitocidal Bacillus thuringiensis strains in Cry11A-resistant larvae: Presence of Cry11A-like toxins in these strains. FEMS Microbiol. Lett. 1997, 153, 419–424. [Google Scholar] [CrossRef]
- Federici, B.A.; Park, H.W.; Bideshi, D.K.; Wirth, M.C.; Johnson, J.J. Recombinant bacteria for mosquito control. J. Exp. Biol. 2003, 206, 3877–3885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juárez-pérez, V.; Porcar, M.; Orduz, S.; Delécluse, A. Cry29A and Cry30A two novel delta-endotoxins isolated from Bacillus thuringiensis serovar medellin. Syst. Appl. Microbiol. 2003, 504, 502–504. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, N.; Gutierrez, D.; Patiño, M.M.; Thiéry, I.; Delécluse, A.; Orduz, S. Cloning, expression and toxicity of a mosquitocidal toxin gene of Bacillus thuringiensis subsp. medellin. Mem. Inst. Oswaldo Cruz 1997, 92, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, N.; Chawla, S.; Likitvivatanavong, S.; Lee, H.L.; Gill, S.S. The cry toxin operon of Clostridium bifermentans subsp. Malaysia is highly toxic to Aedes larval mosquitoes. Appl. Environ. Microbiol. 2014, 80, 5689–5697. [Google Scholar] [CrossRef] [Green Version]
- Rosso, M.L.; Delécluse, A. Contribution of the 65-kilodalton protein encoded by the cloned gene cry19A to the mosquitocidal activity of Bacillus thuringiensis subsp. jegathesan. Appl. Environ. Microbiol. 1997, 63, 4449–4455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.H.; Saitoh, H.; Mizuki, E.; Higuchi, K.; Ohba, M. A novel class of mosquitocidal δ-endotoxin, Cry19B, encoded by a Bacillus thuringiensis serovar higo gene. Syst. Appl. Microbiol. 1998, 21, 179–184. [Google Scholar] [CrossRef]
- Lee, H.; Gill, S.S. Molecular cloning and characterization of a novel mosquitocidal protein gene from Bacillus thuringiensis subsp. fukuokaensis. Appl. Environ. Microbiol. 1997, 63, 4664–4670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berón, C.M.; Salerno, G.L. Cloning and characterization of a novel crystal protein from a native Bacillus thuringiensis isolate highly active against Aedes aegypti. Curr. Microbiol. 2007, 54, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, H.; Hwang, S.H.; Park, Y.S.; Higuchi, K.; Mizuki, E.; Ohba, M. Cloning and characterization of a Bacillus thuringiensis serovar higo gene encoding a novel class of the δ-endotoxin protein, Cry27A, specifically active on the anopheles mosquito. Syst. Appl. Microbiol. 2000, 23, 25–30. [Google Scholar] [CrossRef]
- Tan, F.; Zheng, A.; Zhu, J.; Wang, L.; Li, S.; Deng, Q.; Wang, S.; Li, P.; Tang, X. Rapid cloning, identification, and application of one novel crystal protein gene cry30Fa1 from Bacillus thuringiensis. FEMS Microbiol. Lett. 2010, 302, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Bando, H.; Asano, S. ichiro Activation process of the mosquitocidal δ-endotoxin Cry39A produced by Bacillus thuringiensis subsp. aizawai BUN1-14 and binding property to Anopheles stephensi BBMV. J. Invertebr. Pathol. 2006, 93, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Sahara, K.; Bando, H.; Asano, S. Cloning and expression of novel crystal protein genes cry39A and 39orf2 from Bacillus thuringiensis subsp. aizawai Bun1-14 encoding mosquitocidal proteins. J. Insect Biotechnol. Sericology 2002, 71, 123–128. [Google Scholar]
- Hayakawa, T.; Sakakibara, A.; Ueda, S.; Azuma, Y.; Ide, T.; Takebe, S. Cry46Ab from Bacillus thuringiensis TK-E6 is a new mosquitocidal toxin with aerolysin-type architecture. Insect Biochem. Mol. Biol. 2017, 87, 100–106. [Google Scholar] [CrossRef]
- Gough, J.M.; Kemp, D.H.; Akhurst, R.J.; Pearson, R.D.; Kongsuwan, K. Identification and characterization of proteins from Bacillus thuringiensis with high toxic activity against the sheep blowfly, Lucilia cuprina. J. Invertebr. Pathol. 2005, 90, 39–46. [Google Scholar] [CrossRef]
- Kongsuwan, K.; Gough, J.; Kemp, D.; McDevitt, A.; Akhurst, R. Characterization of a new Bacillus thuringiensis endotoxin, Cry47Aa, from strains that are toxic to the australian sheep blowfly, Lucilia cuprina. FEMS Microbiol. Lett. 2005, 252, 127–136. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, S.; Peng, S.; Gong, J.; Qian, J.; He, J.; Dai, W.; Wang, R. Characterization of a novel mosquitocidal toxin of Cry50Ba and its potential synergism with other mosquitocidal toxins. Toxicon 2017, 138, 165–168. [Google Scholar] [CrossRef]
- Tan, F.; Zhu, J.; Tang, J.; Tang, X.; Wang, S.; Zheng, A.; Li, P. Cloning and characterization of two novel crystal protein genes, cry54Aa1 and cry30Fa1, from Bacillus thuringiensis strain BtMC28. Curr. Microbiol. 2009, 58, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zheng, A.P.; Tan, F.R.; Wang, S.Q.; Deng, Q.M.; Li, S.C.; Wang, L.X.; Li, P. Characterisation and expression of a novel holotype crystal protein gene, cry56Aa1, from Bacillus thuringiensis strain Ywc2-8. Biotechnol. Lett. 2010, 32, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhao, Q.; Xia, L.; Ding, X.; Hu, Q.; Federici, B.A.; Park, H.-W. Identification and characterization of three previously undescribed crystal proteins from Bacillus thuringiensis subsp. jegathesan. Appl. Environ. Microbiol. 2013, 79, 3364–3370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wu, Z.; Zhang, J.; Wan, Y.; Jin, W.; Li, Y.; Fang, X. Cry80Aa1, a novel Bacillus thuringiensis toxin with mosquitocidal activity to Culex pipiens pallens. J. Invertebr. Pathol. 2020, 173, 2019–2021. [Google Scholar] [CrossRef] [PubMed]
- Chilcott, C.N.; Wigley, P.J.; Broadwell, A.H.; Park, D.J.; Ellar, D.J. Activities of Bacillus thuringiensis insecticidal crystal proteins Cyt1Aa and Cyt2Aa against three species of sheep blowfly. Appl. Environ. Microbiol. 1998, 64, 4060–4061. [Google Scholar] [CrossRef] [Green Version]
- Thiery, I.; Delécluse, A.; Tamayo, M.C.; Orduz, S. Identification of a gene for Cyt1A-like hemolysin from Bacillus thuringiensis subsp. medellin and expression in a crystal-negative B. thuringiensis strain. Appl. Environ. Microbiol. 1997, 63, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Cheong, H.; Gill, S.S. Cloning and characterization of a cytolytic and mosquitocidal δ- endotoxin from Bacillus thuringiensis subsp. jegathesan. Appl. Environ. Microbiol. 1997, 63, 3254–3260. [Google Scholar] [CrossRef] [Green Version]
- Wirth, M.C.; Georghiou, G.P.; Malik, J.I.; Abro, G.H. Laboratory selection for resistance to Bacillus sphaericus in Culex quinquefasciatus (Diptera: Culicidae) from California, USA. J. Med. Entomol. 2000, 37, 534–540. [Google Scholar] [CrossRef]
- Juárez-Pérez, V.; Guerchicoff, A.; Rubinstein, C.; Delecluse, A. Characterization of Cyt2Bc toxin from Bacillus thuringiensis subsp. medellin. Appl. Environ. Microbiol. 2002, 68, 1228–1231. [Google Scholar]
- Torres-Quintero, M.C.; Gómez, I.; Pacheco, S.; Sánchez, J.; Flores, H.; Osuna, J.; Mendoza, G.; Soberón, M.; Bravo, A. Engineering Bacillus thuringiensis Cyt1Aa toxin specificity from dipteran to lepidopteran toxicity. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Oestergaard, J.; Ehlers, R.U.; Martínez-Ramírez, A.C.; Real, M.D. Binding of Cyt1Aa and Cry11Aa toxins of Bacillus thuringiensis serovar israelensis to brush border membrane vesicles of Tipula paludosa (Diptera: Nematocera) and subsequent pore formation. Appl. Environ. Microbiol. 2007, 73, 3623–3629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koni, P.A.; Ellar, D.J. Biochemical characterization of Bacillus thuringiensis cytolytic d-endotoxins. Microbiology 1994, 140, 1869–1880. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Yu, Y.M.; Dai, S.M.; Law, S.K.; Gill, S.S. High-level cryIVD and cytA gene expression in Bacillus thuringiensis does not require the 20-kilodalton protein, and the coexpressed gene products are synergistic in their toxicity to mosquitoes. Appl. Environ. Microbiol. 1993, 59, 815–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, M.C.; Delecluse, A.; Walton, W.E. Cyt1Ab1 and Cyt2Ba1 from Bacillus thuringiensis subsp. medellin and B. thuringiensis subsp. israelensis synergize Bacillus sphaericus against Aedes aegypti and resistant Culex quinquefasciatus (Diptera: Culicidae). Appl. Environ. Microbiol. 2001, 67, 3280–3284. [Google Scholar] [CrossRef] [Green Version]
- Payne, J.; Narva, K.E.; Uyeda, K.A.; Stalder, C.J.; Michaels, T.E. Bacillus thuringiensisisolate PS201T6 toxin. U.S. Patent 5,436,002, 25 July 1995. [Google Scholar]
- Promdonkoy, B.; Chewawiwat, N.; Tanapongpipat, S.; Luxananil, P.; Panyim, S. Cloning and characterization of a cytolytic and mosquito larvicidal δ-endotoxin from Bacillus thuringiensis subsp. darmstadiensis. Curr. Microbiol. 2003, 46, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, T.; Sun, Z.; Guan, P.; Zhu, J.; Wang, S.; Li, S.; Deng, Q.; Wang, L.; Zheng, A.; et al. Co-expression and synergism analysis of Vip3Aa29 and Cyt2Aa3 insecticidal proteins from Bacillus thuringiensis. Curr. Microbiol. 2012, 64, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Nisnevitch, M.; Cohen, S.; Ben-Dov, E.; Zaritsky, A.; Sofer, Y.; Cahan, R. Cyt2Ba of Bacillus thuringiensis israelensis: Activation by putative endogenous protease. Biochem. Biophys. Res. Commun. 2006, 344, 99–105. [Google Scholar] [CrossRef]
- Thanabalu, T.; Hindley, J.; Berry, C. Proteolytic processing of the mosquitocidal toxin from Bacillus sphaericus SSII-1. J. Bacteriol. 1992, 174, 5051–5056. [Google Scholar] [CrossRef] [Green Version]
- Partridge, M.R.; Berry, C. Insecticidal activity of the Bacillus sphaericus Mtx1 toxin against Chironomus riparus. J. Invertebr. Pathol. 2002, 79, 135–136. [Google Scholar] [CrossRef]
- Chan, S.W.; Thanabalu, T.; Wee, B.Y.; Porter, A.G. Unusual amino acid determinants of host range in the Mtx2 family of mosquitocidal toxins. Biochemistry 1996, 271, 14183–14187. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.W.; Porter, A.G.; Wee, B.Y.; Thanabalu, T. New gene from nine Bacillus sphaericus strains encoding highly conserved 35.8-kilodalton mosquitocidal toxins. Appl. Environ. Microbiol. 1996, 62, 2174–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opota, O.; Vallet-Gély, I.; Vincentelli, R.; Kellenberger, C.; Iacovache, I.; Gonzalez, M.R.; Roussel, A.; van der Goot, F.G.; Lemaitre, B. Monalysin, a novel β-pore-forming toxin from the Drosophila pathogen Pseudomonas entomophila, contributes to host intestinal damage and lethality. PLoS Pathog. 2011, 7, e1002259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marche, M.G.; Mura, M.E.; Falchi, G.; Ruiu, L. Spore surface proteins of Brevibacillus laterosporus are involved in insect pathogenesis. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Contreras, E.; Masuyer, G.; Qureshi, N.; Chawla, S.; Dhillon, H.S.; Lee, H.L.; Chen, J.; Stenmark, P.; Gill, S.S. A neurotoxin that specifically targets Anopheles mosquitoes. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.W.; Nielsen-Leroux, C.; Yang, Y.; Yuan, Z.; Dumas, V.F.; Monnerat, R.G.; Berry, C. A new Cry toxin with a unique two-component dependency from Bacillus sphaericus. FASEB J. 2007, 21, 4112–4120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, C.; Hindley, J.; Ehrhardt, A.F.; Grounds, T.; De Souza, I.; Davidson, E.W. Genetic determinants of host ranges of Bacillus sphaericus mosquito larvicidal toxins. J. Bacteriol. 1993, 175, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, E.W. Variation in binding of Bacillus sphaericus toxin and wheat germ agglutinin to larval midgut cells of six species of mosquitoes. J. Invertebr. Pathol. 1989, 53, 251–259. [Google Scholar] [CrossRef]
- Oei, C.; Hindley, J.; Berry, C. Binding of purified Bacillus sphaericus binary toxin and its deletion derivatives to Culex quinquefasciatus gut: Elucidation of functional binding domains. J. Gen. Microbiol. 1992, 138, 1515–1526. [Google Scholar] [CrossRef] [Green Version]
- Ahantarig, A.; Chantawat, N.; Waterfield, N.R.; Ffrench-Constant, R.; Kittayapong, P. PirAB toxin from Photorhabdus asymbiotica as a larvicide against dengue vectors. Appl. Environ. Microbiol. 2009, 75, 4627–4629. [Google Scholar] [CrossRef] [Green Version]
- González-Villarreal, S.E.; García-Montelongo, M.; Ibarra, J.E. Insecticidal activity of a Cry1Ca toxin of Bacillus thuringiensis Berliner (Firmicutes: Bacillaceae) and its synergism with the Cyt1Aa toxin against Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 1852–1856. [Google Scholar] [CrossRef]
- Manasherob, R.; Itsko, M.; Sela-Baranes, N.; Ben-Dov, E.; Berry, C.; Cohen, S.; Zaritsky, A. Cyt1 Ca from Bacillus thuringiensis subsp. israelensis: Production in Escherichia coli and comparison of its biological activities with those of other Cyt-like proteins. Microbiology 2006, 152, 2651–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, L.J.; Margalith, J. A bacterial spore demonstrating rapid larvicidal activity against Anopheles sergentii, Uranotaenia unguiculata, Culex univitattus, Aedes aegypti and Culex pipiens. Mosq. News 1977, 1, 355–362. [Google Scholar]
- Fillinger, U.; Knols, B.G.J.; Becker, N. Efficacy and efficiency of new Bacillus thuringiensis var. israelensis and Bacillus sphaericus formulations against afrotropical anophelines in western Kenya. Trop. Med. Int. Health 2003, 8, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Margalith, Y.; Ben-Dov, E. Biological control by Bacillus thuringiensis subsp. israelensis. In Insect Pest Management: Techniques for Environmental Protection; Recheigl, J.E., Recheigl, N.A., Eds.; U.C.P.L.: Boca Raton, FL, USA, 2000. [Google Scholar]
- Ben-Dov, E. Bacillus thuringiensis subsp. israelensis and its dipteran-specific toxins. Toxins (Basel) 2014, 6, 1222–1243. [Google Scholar] [CrossRef]
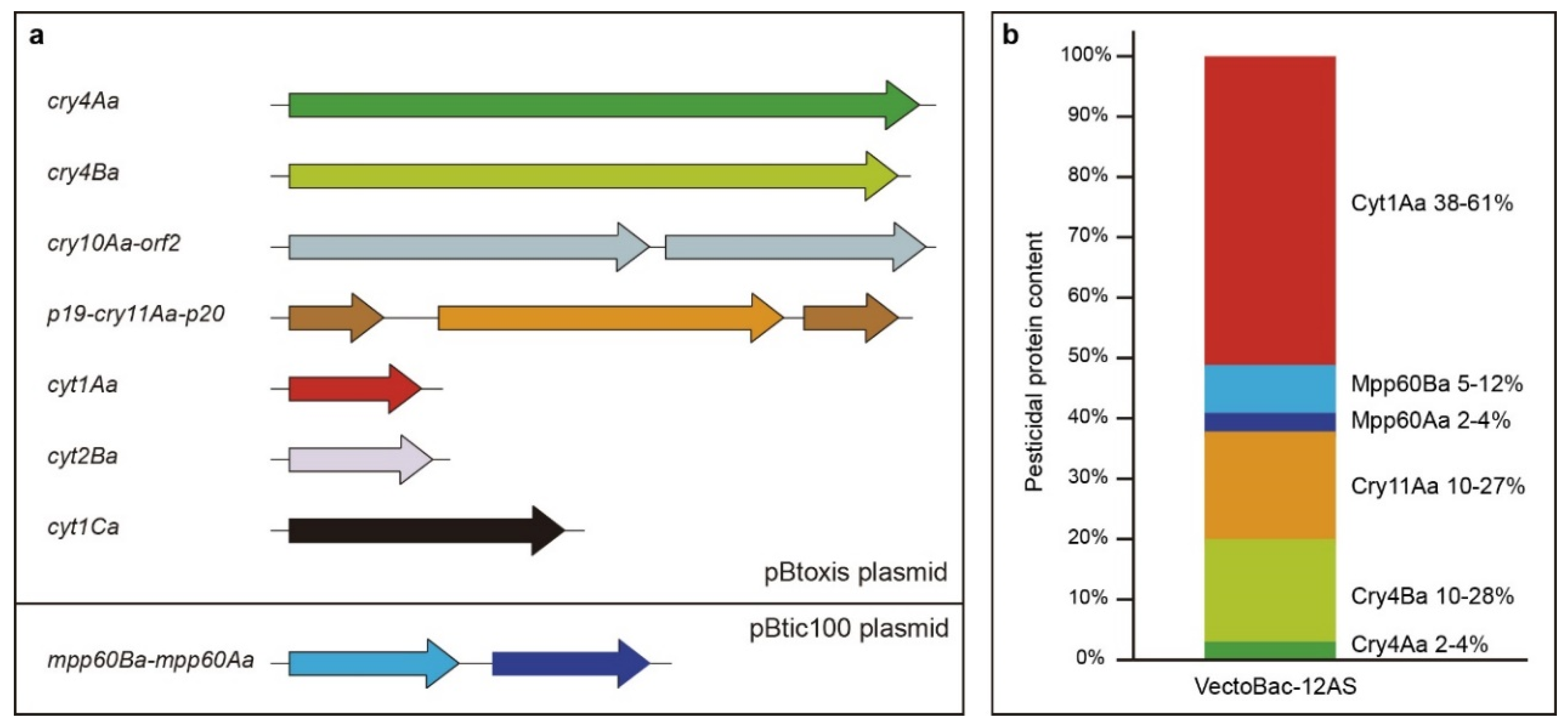
- Berry, C.; Ben-dov, E.; Jones, A.F.; Murphy, L.; Quail, M.A.; Holden, M.T.G.; Harris, D.; Zaritsky, A.; Parkhill, J. Complete sequence and organization of pBtoxis, the toxin-coding plasmid of Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 2002, 68, 5082–5095. [Google Scholar] [CrossRef] [Green Version]
- Bolotin, A.; Gillis, A.; Sanchis, V.; Nielsen-LeRoux, C.; Mahillon, J.; Lereclus, D.; Sorokin, A. Comparative genomics of extrachromosomal elements in Bacillus thuringiensis subsp. israelensis. Res. Microbiol. 2017, 168, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Gillis, A.; Fayad, N.; Makart, L.; Bolotin, A.; Sorokin, A.; Kallassy, M.; Mahillon, J. Role of plasmid plasticity and mobile genetic elements in the entomopathogen Bacillus thuringiensis serovar israelensis. FEMS Microbiol. Rev. 2018, 42, 829–856. [Google Scholar] [CrossRef]
- Caballero, J.; Jiménez-Moreno, N.; Orera, I.; Williams, T.; Fernández, A.B.; Villanueva, M.; Ferré, J.; Caballero, P.; Ancín-Azpilicueta, C. Unraveling the composition of insecticidal crystal proteins in Bacillus thuringiensis: A proteomics approach. Appl. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Bietlot, H.P.L.; Vishnubhatla, I.; Carey, P.R.; Pozsgay, M.; Kaplan, H. Characterization of the cysteine residues and disulphide linkages in the protein crystal of Bacillus thuringiensis. Biochem. J. 1990, 267, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Couche, G.A.; Pfannenstiel, M.A.; Nickerson, K.W. Structural disulfide bonds in the Bacillus thuringiensis subsp. israelensis protein crystal. J. Bacteriol. 1987, 169, 3281–3288. [Google Scholar] [CrossRef] [Green Version]
- Bourgouin, C.; Delecluse, A.; Ribier, J.; Klier, A.; Rapoport, G. A Bacillus thuringiensis subsp. israelensis gene encoding a 125-kilodalton larvicidal polypeptide is associated with inverted repeat sequences. J. Bacteriol. 1988, 170, 3575–3583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otieno-Ayayo, Z.N.; Zaritsky, A.; Wirth, M.C.; Manasherob, R.; Khasdan, V.; Cahan, R.; Ben-Dov, E. Variations in the mosquito larvicidal activities of toxins from Bacillus thuringiensis ssp. israelensis. Environ. Microbiol. 2008, 10, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Thorne, L.; Garduno, F.; Thompson, T.; Decker, D.; Zounes, M.; Wild, M.; Waldfield, A.; Pollock, T.J. Structural similarity between the Lepidoptera- and Diptera-specific insecticidal endotoxin genes of Bacillus thuringiensis subsp. “kurstaki” and “israelensis”. J. Bacteriol. 1986, 166, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.G.; Eckblad, W.; Lee, A.B. Diversity of protein inclusion bodies and identification of mosquitocidal protein in Bacillus thuringiensis subsp. israelensis. Biochem. Biophys. Res. Commun. 1985, 126, 953–960. [Google Scholar] [CrossRef]
- Garduno, F.; Thorne, L.; Walfield, A.M.; Pollock, T.J. Structural relatedness between mosquitocidal endotoxins of Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 1988, 54, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, P.; Alzate, O.; Orduz, S. A theoretical model of the tridimensional structure of Bacillus thuringiensis subsp. medellin Cry 11Bb toxin deduced by homology modelling. Mem. Inst. Oswaldo Cruz 2001, 96, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Dervyn, E.; Poncet, S.; Klier, A.; Rapoport, G. Transcriptional regulation of the cryIVD gene operon from Bacillus thuringiensis subsp. israelensis. J. Bacteriol. 1995, 177, 2283–2291. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Federici, B.A. Improved production of the insecticidal CryIVD protein in Bacillus thuringiensis using cryIA(c) promoters to express the gene for an associated 20 kDa protein. Appl. Microbiol. Biotechnol. 1995, 42, 697–702. [Google Scholar] [CrossRef]
- Yamagiwa, M.; Sakagawa, K.; Sakai, H. Functional analysis of two processed fragments of Bacillus thuringiensis Cry11A toxin. Biosci. Biotechnol. Biochem. 2004, 68, 523–528. [Google Scholar] [CrossRef] [Green Version]
- Dai, S.M.; Gill, S.S. In vitro and in vivo proteolysis of the Bacillus thuringiensis subsp. israelensis CryIVD protein by Culex quinquefasciatus larval midgut proteases. Insect Biochem. Mol. Biol. 1993, 23, 273–283. [Google Scholar] [CrossRef]
- Fernandez, L.E.; Aimanova, K.G.; Gill, S.S.; Bravo, A.; Soberón, M. A GPI-anchored alkaline phosphatase is a functional midgut receptor of Cry11Aa toxin in Aedes aegypti larvae. Biochem. J. 2006, 394, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Aimanova, K.G.; Pan, S.; Gill, S.S. Identification and characterization of Aedes aegypti aminopeptidase N as a putative receptor of Bacillus thuringiensis Cry11A toxin. Insect Biochem. Mol. Biol. 2009, 39, 688–696. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Aimanova, K.G.; Fernandez, L.E.; Bravo, A.; Soberon, M.; Gill, S.S. Aedes aegypti cadherin serves as a putative receptor of the Cry11Aa toxin from Bacillus thuringiensis subsp. israelensis. Biochem. J. 2009, 424, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Pérez, C.; Fernández, L.E.; Gill, S.S.; Segovia, L.; Bravo, A.; Rodríguez, M.H.; Soberón, M. Cry11Aa toxin from Bacillus thuringiensis binds its receptor in Aedes aegypti mosquito larvae through loop α-8 of domain II. FEBS Lett. 2005, 579, 3508–3514. [Google Scholar]
- Likitvivatanavong, S.; Gill, S.S.; Bravo, A.; Soberón, M. Cadherin, alkaline phosphatase, and aminopeptidase N as receptors of Cry11Ba toxin from Bacillus thuringiensis subsp. jegathesan in Aedes aegypti. Appl. Environ. Microbiol. 2011, 77, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, I.; Sorokin, A.; Kapatral, V.; Reznik, G.; Bhattacharya, A.; Mikhailova, N.; Burd, H.; Joukov, V.; Kaznadzey, D.; Walunas, T.; et al. Comparative genome analysis of Bacillus cereus group genomes with Bacillus subtilis. FEMS Microbiol. Lett. 2005, 250, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Chilcott, C.N.; Wigley, P.J. Opportunities for finding new Bacillus thuringiensis strains. Agric. Ecosyst. Environ. 1994, 49, 51–57. [Google Scholar] [CrossRef]
- Saleem, F.; Shakoori, A.R. Characterization of cry2A-type gene(s) from pakistani isolates of Bacillus thuringiensis toxic to lepidopteran and dipteran insects. Pak. J. Zool. 2010, 42, 181–193. [Google Scholar]
- Nicholls, C.N.; Ahmad, W.; Ellar, D.J. Evidence for two different types of insecticidal P2 toxins with dual specificity in Bacillus thuringiensis subspecies. J. Bacteriol. 1989, 171, 5141–5147. [Google Scholar] [CrossRef] [Green Version]
- Goje, L.J.; Elmi, E.D.; Bracuti, A.; Courty, T.; Rao, T.; Alzahrani, F.A.; Crickmore, N. Identification of Aedes aegypti specificity motifs in the N-terminus of the Bacillus thuringiensis Cry2Aa pesticidal protein. J. Invertebr. Pathol. 2020, 174, 107423. [Google Scholar] [CrossRef]
- Shu, C.; Zhang, F.; Chen, G.; Joseph, L.; Barqawi, A.; Evans, J.; Song, F.; Li, G.; Zhang, J.; Crickmore, N. A natural hybrid of a Bacillus thuringiensis Cry2A toxin implicates Domain I in specificity determination. J. Invertebr. Pathol. 2017, 150, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Chilcott, C.; Ellar, D. Comparative toxicity of Bacillus thuringiensis var. israelensis crystal proteins in vivo and in vitro. J. Gen. Microbiol. 1988, 134, 2551–2558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, L.F.; Visick, J.E.; Whiteley, H.R. A 20-kilodalton protein is required for efficient production of the Bacillus thuringiensis subsp. israelensis 27-kilodalton crystal protein in Escherichia coli. J. Bacteriol. 1989, 171, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-yahyaee, S.A.S.; Ellar, D.J. Maximal toxicity of cloned CytA δ-endotoxin from Bacillus thuringiensis subsp. israelensis requires proteolytic processing from both the N- and C-termini. Microbiology 1995, 141, 3141–3148. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Singh, G.J.P.; Hornung, J.M. Cell membrane interaction of Bacillus thuringiensis subsp. israelensis cytolytic toxins. Infect. Immun. 1987, 55, 1300–1308. [Google Scholar] [CrossRef] [Green Version]
- Haider, M.Z.; Ellar, D.J. Mechanism of action of Bacillus thuringiensis insecticidal δ-endotoxin: Interaction with phospholipid vesicles. BBA Biomembr. 1989, 978, 216–222. [Google Scholar] [CrossRef]
- Knowles, B.H.; Blatt, M.R.; Tester, M.; Horsnell, J.M.; Carroll, J.; Menestrina, G.; Ellar, D.J. A cytolytic δ-endotoxin from Bacillus thuringiensis var. israelensis forms cation-selective channels in planar lipid bilayers. FEBS Lett. 1989, 244, 259–262. [Google Scholar] [CrossRef] [Green Version]
- Butko, P.; Huang, F.; Pusztai-Carey, M.; Surewicz, W.K. Membrane permeabilization induced by cytolytic δ-endotoxin CytA from Bacillus thuringiensis var. israelensis. Biochemistry 1996, 35, 11355–11360. [Google Scholar] [CrossRef] [PubMed]
- Butko, P.; Huang, F.; Pusztai-Carey, M.; Surewicz, W.K. Interaction of the δ-endotoxin CytA from Bacillus thuringiensis var. israelensis with lipid membranes. Biochemistry 1997, 36, 12862–12868. [Google Scholar] [CrossRef] [PubMed]
- Anaya, P.; Onofre, J.; Torres-Quintero, M.C.; Sánchez, J.; Gill, S.S.; Bravo, A.; Soberón, M. Oligomerization is a key step for Bacillus thuringiensis Cyt1Aa insecticidal activity but not for toxicity against red blood cells. Insect Biochem. Mol. Biol. 2020, 119, 103317. [Google Scholar] [CrossRef] [PubMed]
- Orduz, S.; Diaz, T.; Restrepo, N.; Patiño, M.M.; Tamayo, M.C. Biochemical, immunological and toxicological characteristics of the crystal proteins of Bacillus thuringiensis subsp. medellin. Mem. Inst. Oswaldo Cruz 1996, 91, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valtierra-de-Luis, D.; Villanueva, M.; Caballero, J.; Matas, I.M.; Williams, T.; Caballero, P. Quantification of dose-mortality responses in adult Diptera: Validation using Ceratitis capitata and Drosophila suzukii responses to spinosad. PLoS ONE 2019, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Guerchicoff, A.; Ugalde, R.A.; Rubinstein, C.P. Identification and characterization of a previously undescribed cyt gene in Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 1997, 63, 2716–2721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Koni, P.A.; Ellar, D.J. Structure of the mosquitocidal δ-endotoxin CytB from Bacillus thuringiensis sp. kyushuensis and implications for membrane pore formation. J. Mol. Biol. 1996, 257, 129–152. [Google Scholar] [CrossRef] [PubMed]
- Loth, K.; Costechareyre, D.; Effantin, G.; Rahbé, Y.; Condemine, G.; Landon, C.; Da Silva, P. New Cyt-like δ -endotoxins from Dickeya dadantii: Structure and aphicidal activity. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef]
- Thanabalu, T.; Hindley, J.; Jackson-Yap, J.; Berry, C. Cloning, sequencing, and expression of a gene encoding a 100-kilodalton mosquitocidal toxin from Bacillus sphaericus SSII-1. J. Bacteriol. 1991, 173, 2776–2785. [Google Scholar] [CrossRef] [Green Version]
- Thanabalu, T.; Berry, C.; Hindley, J. Cytotoxicity and ADP-ribosylating activity of the mosquitocidal toxin from Bacillus sphaericus SSII-1: Possible roles of the 27- and 70-kilodalton peptides. J. Bacteriol. 1993, 175, 2314–2320. [Google Scholar] [CrossRef] [Green Version]
- Thanabalu, T.; Porter, A.G. A Bacillus sphaericus gene encoding a novel type of mosquitocidal toxin of 31.8 kDa. Gene 1996, 170, 85–89. [Google Scholar] [CrossRef]
- Berry, C. The bacterium, Lysinibacillus sphaericus, as an insect pathogen. J. Invertebr. Pathol. 2012, 109, 1–10. [Google Scholar] [CrossRef]
- Jones, G.W.; Wirth, M.C.; Monnerat, R.G.; Berry, C. The Cry48Aa-Cry49Aa binary toxin from Bacillus sphaericus exhibits highly restricted target specificity. Environ. Microbiol. 2008, 10, 2418–2424. [Google Scholar] [CrossRef] [Green Version]
- Silva Filha, M.H.N.L.; Berry, C.; Regis, L. Lysinibacillus sphaericus: Toxins and mode of action, applications for mosquito control and resistance management. In Advances in Insect Physiology; Academic Press: Cambridge, MA, USA, 2014; Volume 47, pp. 89–176. ISBN 9780128001974. [Google Scholar]
- Pener, H.; Wilamowski, A. Susceptibility of larvae of the sandfly Phlebotomus papatasi (Diptera: Psychodidae) to Bacillus sphaericus. Bull. Entomol. Res. 1996, 86, 173–175. [Google Scholar] [CrossRef]
- Yiallouros, M.; Storch, V.; Thiery, I.; Becker, N. Efficacy of Clostridium bifermentans serovar Malaysia on target and nontarget organisms. J. Am. Mosq. Control Assoc. 1994, 10, 51–55. [Google Scholar]
- Nicolas, L.; Charles, J.-F.; de Barjac, H. Clostridium bifermentans serovar malaysia: Characterization of putative mosquito larvicidal proteins. FEMS Microbiol. Lett. 1993, 113, 23–28. [Google Scholar] [CrossRef]
- Barloy, F.; Delécluse, A.; Nicolas, L.; Lecadet, M.M. Cloning and expression of the first anaerobic toxin gene from Clostridium bifermentans subsp. malaysia, encoding a new mosquitocidal protein with homologies to Bacillus thuringiensis delta-endotoxins. J. Bacteriol. 1996, 178, 3099–3105. [Google Scholar] [CrossRef] [Green Version]
- Juárez-Pérez, V.; Delécluse, A. The cry toxins and the putative hemolysins of Clostridium bifermentans ser. malaysia are not involved in mosquitocidal activity. J. Invertebr. Pathol. 2001, 78, 57–58. [Google Scholar]
- Lee, C.T.; Chen, I.T.; Yang, Y.T.; Ko, T.P.; Huang, Y.T.; Huang, J.Y.; Huang, M.F.; Lin, S.J.; Chen, C.Y.; Lin, S.S.; et al. The opportunistic marine pathogen Vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin. Proc. Natl. Acad. Sci. USA 2015, 112, 10798–10803. [Google Scholar] [CrossRef] [Green Version]
- Soberón, M.; López-Díaz, J.A.; Bravo, A. Cyt toxins produced by Bacillus thuringiensis: A protein fold conserved in several pathogenic microorganisms. Peptides 2013, 41, 87–93. [Google Scholar] [CrossRef]
- Georghiou, G.P.; Wirth, M.C. Influence of exposure to single versus multiple toxins of Bacillus thuringiensis subsp. israelensis on development of resistance in the mosquito Culex quinquefasciatus (Diptera: Culicidae). Appl. Environ. Microbiol. 1997, 63, 1095–1101. [Google Scholar] [CrossRef] [Green Version]
- Wirth, M.C.; Georghiou, G.P. Cross-Resistance among CryIV toxins of Bacillus thuringiensis subsp israelensis in Culex quinquefasciatus (Diptera: Culicidae). J. Econ. Entomol. 1997, 90, 1471. [Google Scholar] [CrossRef]
- Wirth, M.C.; Park, H.W.; Walton, W.E.; Federici, B.A. Cyt1A of Bacillus thuringiensis delays evolution of resistance to Cry11A in the mosquito Culex quinquefasciatus. Appl. Environ. Microbiol. 2005, 71, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Cantón, P.E.; Reyes, E.Z.; Ruiz De Escudero, I.; Bravo, A.; Soberon, M. Binding of Bacillus thuringiensis subsp. israelensis Cry4Ba to Cyt1Aa has an important role in synergism. Peptides 2011, 32, 595–600. [Google Scholar]
- Perez, C.; Fernandez, L.E.; Sun, J.; Folch, J.L.; Gill, S.S.; Soberon, M.; Bravo, A. Bacillus thuringiensis subsp. israelensis Cyt1Aa synergizes Cry11Aa toxin by functioning as a membrane-bound receptor. Proc. Natl. Acad. Sci. USA 2005, 102, 18303–18308. [Google Scholar] [CrossRef] [Green Version]
- Pérez, C.; Muñoz-Garay, C.; Portugal, L.C.; Sánchez, J.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis ssp. israelensis Cyt1Aa enhances activity of Cry11Aa toxin by facilitating the formation of a pre-pore oligomeric structure. Cell. Microbiol. 2007, 9, 2931–2937. [Google Scholar]
- Elleuch, J.; Tounsi, S.; Belguith Ben Hassen, N.; Lacoix, M.N.; Chandre, F.; Jaoua, S.; Zribi Zghal, R. Characterisation of novel Bacillus thuringiensis isolates against Aedes aegypti (Diptera: Culicidae) and Ceratitis capitata (Diptera: Tephridae). J. Invertebr. Pathol. 2015, 124, 90–97. [Google Scholar] [CrossRef]
- López-Diaz, J.A.; Cantón, P.E.; Gill, S.S.; Soberón, M.; Bravo, A. Oligomerization is a key step in Cyt1Aa membrane insertion and toxicity but not necessary to synergize Cry11Aa toxicity in Aedes aegypti larvae. Environ. Microbiol. 2013, 15, 3030–3039. [Google Scholar] [CrossRef] [Green Version]
- Lailak, C.; Khaokhiew, T.; Promptmas, C.; Promdonkoy, B.; Pootanakit, K.; Angsuthanasombat, C. Bacillus thuringiensis Cry4Ba toxin employs two receptor-binding loops for synergistic interactions with Cyt2Aa2. Biochem. Biophys. Res. Commun. 2013, 435, 216–221. [Google Scholar] [CrossRef]
- Thiéry, I.; Hamon, S.; Delécluse, A.; Orduz, S. The introduction into Bacillus sphaericus of the Bacillus thuringiensis subsp. medellin cyt1Ab1 gene results in higher susceptibility of resistant mosquito larvae populations to B. sphaericus. Appl. Environ. Microbiol. 1998, 64, 3910–3916. [Google Scholar] [CrossRef] [Green Version]
- Wirth, M.C. Mosquito resistance to bacterial larvicidal toxins. Open Toxinology J. 2010, 3, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Becker, N. Ice granules containing endotoxins of microbial agents for the control of mosquito larvae—A new application technique. J. Am. Mosq. Control Assoc. 2003, 19, 63–66. [Google Scholar]
- Aly, C. Filtration rates of mosquito larvae in suspensions of latex microspheres and yeast cells. Entomol. Exp. Appl. 1988, 46, 55–61. [Google Scholar] [CrossRef]
- Lacey, L.A.; Smittle, B.J. The effects of gamma radiation on espore viability and mosquito larvicidal activity of Bacillus sphaericus and Bacillus thuringiensis var. israelensis. Bull. Soc. Vector Ecol. 1985, 10, 98–101. [Google Scholar]
- Burges, H.D. Formulation of Microbial Biopesticides: Beneficial Microorganisms, Nematodes and Seed Treatments; Springer: Dordrecht, The Netherlands, 1998. [Google Scholar]
- Lacey, L. Microbial control of black flies and mosquitoes. Annu. Rev. Entomol. 1986, 31, 265–296. [Google Scholar] [CrossRef]
- Mulligan, F.S.; Schaefer, C.H.; Wilder, W.H. Efficacy and persistence of Bacillus sphaericus and B. thuringiensis H. 14 against mosquitoes under laboratory and field conditions. J. Econ. Entomol. 1980, 73, 684–688. [Google Scholar] [CrossRef]
- Skovmand, O.; Bauduin, S. Efficacy of a granular formulation of Bacillus sphaericus against Culex quinquefasciatus and Anopheles gambiae in west african countries. J. Vector Ecol. 1998, 43–51. [Google Scholar]
- Bar, E.; Sandler, N.; Makayoto, M.; Keynan, A. Expression of chromosomally inserted Bacillus thuringiensis israelensis toxin genes in Bacillus sphaericus. J. Invertebr. Pathol. 1998, 72, 206–213. [Google Scholar] [CrossRef]
- Poncet, S.; Bernard, C.; Dervyn, E.; Cayley, J.; Klier, A.; Rapoport, G. Improvement of Bacillus sphaericus toxicity against dipteran larvae by integration, via homologous recombination, of the Cry11A toxin gene from Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 1997, 63, 4413–4420. [Google Scholar] [CrossRef] [Green Version]
- Lacey, L.A. Bacillus thuringiensis serovariety israelensis and Bacillus sphaericus for mosquito control. J. Am. Mosq. Control Assoc. 2007, 23, 133–163. [Google Scholar] [CrossRef]
- Priest, F.G. Biological control of mosquitoes and other biting flies by Bacillus sphaericus and Bacillus thuringiensis. J. Appl. Bacteriol. 1992, 72, 357–369. [Google Scholar] [CrossRef]
- Ingabire, C.M.; Hakizimana, E.; Rulisa, A.; Kateera, F.; Van Den Borne, B.; Muvunyi, C.M.; Mutesa, L.; Van Vugt, M.; Koenraadt, C.J.M.; Takken, W.; et al. Community-based biological control of malaria mosquitoes using Bacillus thuringiensis var. israelensis (Bti) in Rwanda: Community awareness, acceptance and participation. Malar. J. 2017, 16, 1–13. [Google Scholar] [CrossRef]
- Lacey, L.A.; Escaffre, H.; Philippon, B.; Sékétéli, A.; Guillet, P. Large river treatment with Bacillus thuringiensis (H-14) for the control of Simulium damnosum s.l. in the onchocerciasis control programme. Trop. Parasitol 1982, 33, 97–101. [Google Scholar]
- Becker, N. Microbial control of mosquitoes: Management of the upper rhine mosquito population as a model programme. Parasitol. Today 1997, 485–487. [Google Scholar] [CrossRef]
- Chalegre, K.D.D.M.; Romão, T.P.; Amorim, L.B.; Anastacio, D.B.; De Barros, R.A.; De Oliveira, C.M.F.; Regis, L.; De-Melo-Neto, O.P.; Silva-Filha, M.H.N.L. Detection of an allele conferring resistance to Bacillus sphaericus binary toxin in Culex quinquefasciatus populations by molecular screening. Appl. Environ. Microbiol. 2009, 75, 1044–1049. [Google Scholar] [CrossRef] [Green Version]
- Bonin, A.; Paris, M.; Frérot, H.; Bianco, E.; Tetreau, G.; Després, L. The genetic architecture of a complex trait: Resistance to multiple toxins produced by Bacillus thuringiensis israelensis in the dengue and yellow fever vector, the mosquito Aedes aegypti. Infect. Genet. Evol. 2015, 35, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Gammon, K.; Jones, G.W.; Hope, S.J.; De Oliveira, C.M.F.; Regis, L.; Silva Filha, M.H.N.L.; Dancer, B.N.; Berry, C. Conjugal transfer of a toxin-coding megaplasmid from Bacillus thuringiensis subsp. israelensis to mosquitocidal strains of Bacillus sphaericus. Appl. Environ. Microbiol. 2006, 72, 1766–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federici, B.A.; Park, H.; Bideshi, D.K. Overview of the basic biology of Bacillus thuringiensis with emphasis on genetic engineering of bacterial larvicides for mosquito control. Open Toxinology J. 2010, 3, 154–171. [Google Scholar] [CrossRef] [Green Version]
- Csiszàr, K.; Houmard, J.; Damerval, T.; de Marsac, N.T. Transcriptional analysis of the cyanobacterial gvpABC operon in differentiated cells: Occurrence of an antisense RNA complementary to three overlapping transcripts. Gene 1987, 60, 29–37. [Google Scholar] [CrossRef]
- Xu, X.; Kong, R.; Hu, Y. High larvicidal activity of intact recombinant cyanobacterium Anabaena sp. PCC 7120 expressing Gene 51 and Gene 42 of Bacillus sphaericus sp. 2297. FEMS Microbiol. Lett. 1993, 107, 247–250. [Google Scholar]
- Wai, H.Y.; Thanabalu, T.; Porter, A.G. Expression of mosquitocidal toxin genes in a gas-vacuolated strain of Ancylobacter aquaticus. Appl. Environ. Microbiol. 1994, 60, 4199–4202. [Google Scholar]
- Luxananil, P.; Tanapongpipat, S.; Promdonkoy, B.; Atomi, H.; Imanaka, T.; Panyim, S. Expression of binary toxin genes in the mosquito-colonizable bacteria, Bacillus cereus, leads to high toxicity against Culex quinquefasciatus larvae. Curr. Microbiol. 2003, 47, 372–375. [Google Scholar] [CrossRef]
- Zaritsky, A.; Zalkinder, V.; Ben-Dov, E.; Barak, Z. Bioencapsulation and delivery to mosquito larvae of Bacillus thuringiensis H14 toxicity by Tetrahymena pyriformis. J. Invertebr. Pathol. 1991, 58, 455–457. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valtierra-de-Luis, D.; Villanueva, M.; Berry, C.; Caballero, P. Potential for Bacillus thuringiensis and Other Bacterial Toxins as Biological Control Agents to Combat Dipteran Pests of Medical and Agronomic Importance. Toxins 2020, 12, 773. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120773
Valtierra-de-Luis D, Villanueva M, Berry C, Caballero P. Potential for Bacillus thuringiensis and Other Bacterial Toxins as Biological Control Agents to Combat Dipteran Pests of Medical and Agronomic Importance. Toxins. 2020; 12(12):773. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120773
Chicago/Turabian StyleValtierra-de-Luis, Daniel, Maite Villanueva, Colin Berry, and Primitivo Caballero. 2020. "Potential for Bacillus thuringiensis and Other Bacterial Toxins as Biological Control Agents to Combat Dipteran Pests of Medical and Agronomic Importance" Toxins 12, no. 12: 773. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120773