Scorpion Species with Smaller Body Sizes and Narrower Chelae Have the Highest Venom Potency


1
Venom Systems & Proteomics Lab, School of Natural Sciences, Ryan Institute, National University of Ireland Galway, H91 TK33 Galway, Ireland
2
Macroecology Lab, School of Natural Sciences, Ryan Institute, National University of Ireland Galway, H91 TK33 Galway, Ireland
*
Author to whom correspondence should be addressed.
Toxins 2022, 14(3), 219; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14030219
Submission received: 4 February 2022
/
Revised: 10 March 2022
/
Accepted: 14 March 2022
/
Published: 17 March 2022
(This article belongs to the Special Issue Drivers of Venom Potency across the Animal Kingdom)
Abstract
:Scorpionism is a global health concern, with an estimation of over one million annual envenomation cases. Despite this, little is known regarding the drivers of scorpion venom potency. One widely held view is that smaller scorpions with less-developed chelae possess the most potent venoms. While this perception is often used as a guide for medical intervention, it has yet to be tested in a formal comparative framework. Here, we use a phylogenetic comparative analysis of 36 scorpion species to test whether scorpion venom potency, as measured using LD50, is related to scorpion body size and morphology. We found a positive relationship between LD50 and scorpion total length, supporting the perception that smaller scorpions possess more potent venoms. We also found that, independent of body size, scorpion species with long narrow chelae have higher venom potencies compared to species with more robust chelae. These results not only support the general perception of scorpion morphology and potency, but also the presence of an ecology trade-off with scorpions either selected for well-developed chelae or more potent venoms. Testing the patterns of venom variations in scorpions aids both our ecological understanding and our ability to address the global health burden of scorpionism.
Keywords:
venom; scorpions; LD50; potency; body size; chela morphology; telson morphology; phylogenetic comparative analyses; defense mechanisms; evolutionary trade-offKey Contribution: We conducted a comparative analysis, testing the relationship between the overall size of scorpions and their morphological features, including the relationship between the chela and telson of scorpions with their venom potency (LD50). We found that larger species of scorpions, with robust pedipalps, are less venomous than small scorpions, with slender pedipalps.
1. Introduction
Envenomation, resulting from a scorpion sting, referred to as scorpionism [1], is a major global public concern affecting Central America, South America, North Africa, the Middle East and West Asia [2]. Approximately 1.2 million scorpion stings are estimated to occur throughout the world each year, 3250 of which result in death [3,4]. Despite the global burden of scorpionism, which is likely to be higher due to unreported envenomation events [5,6], surprisingly little is understood regarding the ecology and evolution of scorpions and their venom. Such a gap in understanding and data can lead to inaccurate treatment of scorpion envenomation, through misidentification of species [7] or the misuse of antivenoms [8]. Furthermore, understanding the ecology and evolution of venoms can aid in predicting envenomation risk and its treatment, as demonstrated by the efforts related to snakebites [9]. Ecological factors have been linked to severe scorpionism, including climatic variations, venom metering and the overall morphology of the scorpion [10,11]. Understanding the ecological and evolutionary drivers of scorpion venom potency not only contributes to our fundamental understanding of these species, but also to our ability to address the global burden of scorpionism.
The concept that larger scorpions have less potent venom compared to smaller scorpions is widely expressed in the media, and has been popularized by numerous sources, such as in the Indiana Jones movie, where the main protagonist explicitly refers to this by saying “When it comes to scorpions, the bigger the better” [12]. Apart from its use in popular fiction, the concept of relying on the size of the morphological characteristics of scorpions is used as a general medical guidance in many countries where scorpionism is an issue [13,14], as most scorpions are difficult to identify at the species level by non-experts [14,15,16,17]. Support for this general concept can be found in the high median lethal dosage values (LD50), indicating venoms of lower potency, reported for larger species of scorpions, such as Hadrurus arizonensis and Heterometrus laoticus [18,19,20,21]. However, such general rules should be used with caution. For example, in Morocco, black scorpions are often considered as the most lethal species [22], despite evidence that the color of scorpions does not relate to lethality, and is usually determined by habitat [23]. However, unlike color, body size may have a clearer potential mechanistic relationship with venom potency.
Scorpions possess the following two main means to capture prey and dissuade predators: their venomous stinging apparatus and their pedipalps [24]. While present in all scorpions, their use varies across species, following a trade-off pattern [16]. Species that primarily rely on venom, such as many species within Buthidae, often have long slender chelae [5,24], while species with more powerful chelae, such as Opistopthalmus glabrifrons, typically have comparatively less-developed stinging apparatus, with a smaller metasoma, or less potent venom, as found by Lourenço [15]. From an evolutionary perspective, such a pattern may emerge, as larger species that rely more on mechanical prey capture or that avoid predators may have fewer selection pressures on the potency of their venoms. Interestingly, not only has size been used as a medical indicator in envenomation cases [25], the size of the pedipalps has been used to determine the lethality of scorpions [24,25]. However, despite popular support for these patterns, they have yet to be tested using phylogenetic comparative methods.
While phylogenetic comparative methods are a relatively recent approach to understanding venom variation [26], studies measuring venom potencies have a rich history, with numerous standardized measures of potencies, such as LD50, available across species spanning the scorpion phylogeny. Here, we use this rich history to collate LD50 potency values [20,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44] and morphology measures [14,19,35,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83] from the literature to test the commonly held belief that larger scorpions, with more powerful chelae, are comparatively less potent than smaller species. We predict that smaller species, with narrower chelae and larger telsons, will have lower LD50 values, indicating higher venom potency.
2. Results
Our dataset consisted of 36 scorpion species with 62 measures of LD50, with all the measures that met our criterion for inclusion using mammalian Mus musculus models (S1). The lethality of the species within the dataset ranges from 0.16 mg/kg−1 for the deathstalker Leiurus quinquestriatus to 1800 mg/kg−1 for the rock scorpion Hadogenes granulatus. The median body size was 70 mm, ranging from 40.5 mm for the bark scorpion Scorpion Centruroides noxius to 200 mm for H. granulatus. The chela ratio ranged from 0.7 to 6.2, with the most robust chelae found in the large-clawed scorpion Scorpio maurus and the red scorpion Rhopalurus junceus, with the most slender chelae found in the deathstalker Leiurus quinquestriatus.
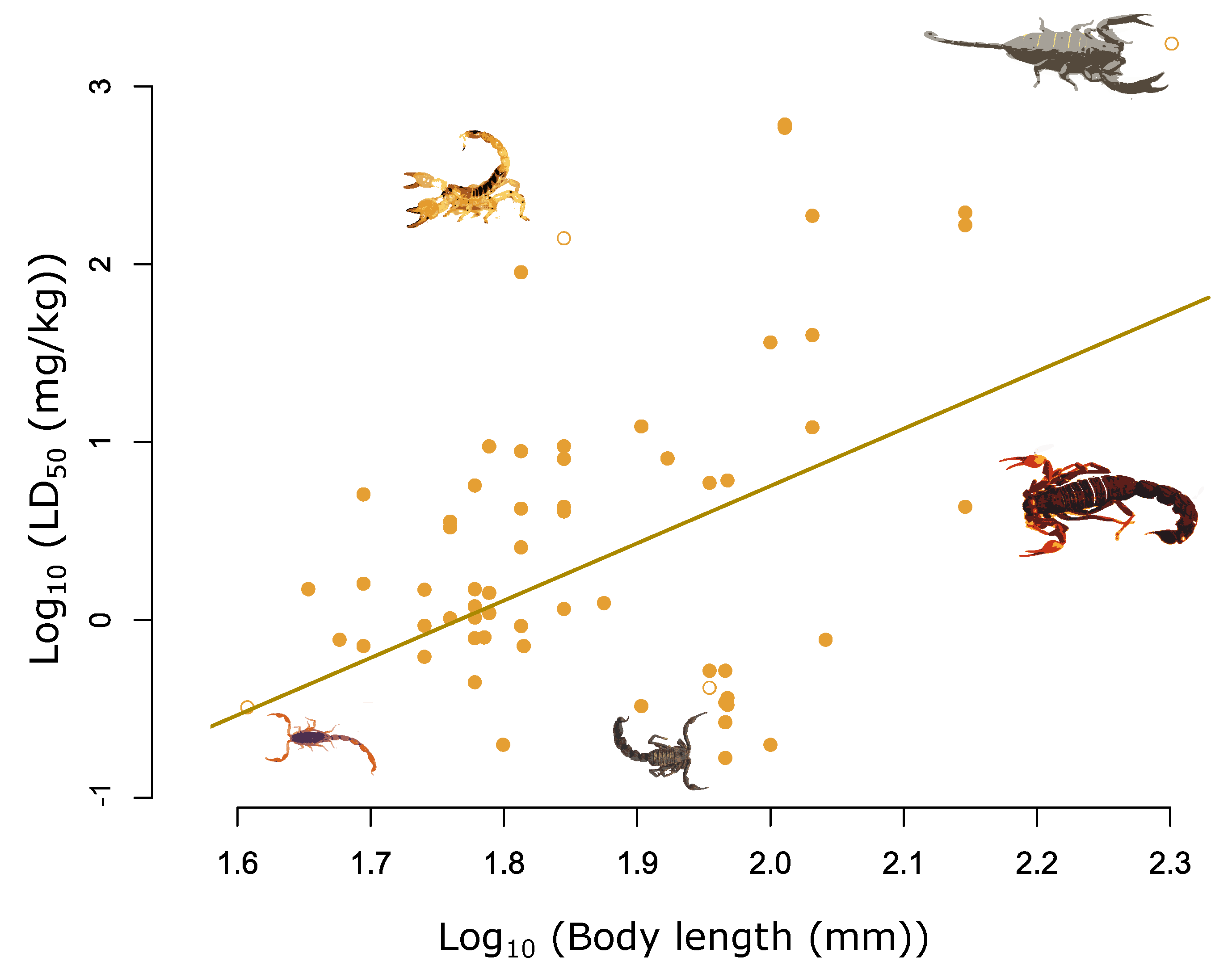
We found that body length was positively correlated with LD50, with the larger scorpions being associated with less potent venoms (Figure 1 and Figure S1 and Table 1). The significant slope between log10 body length and log10 LD50 found in our analysis corresponds to a change in LD50 of 187 mg/kg−1 across the 159 mm range of body length in our dataset (Figure 1 and Figure S1 and Table 1).
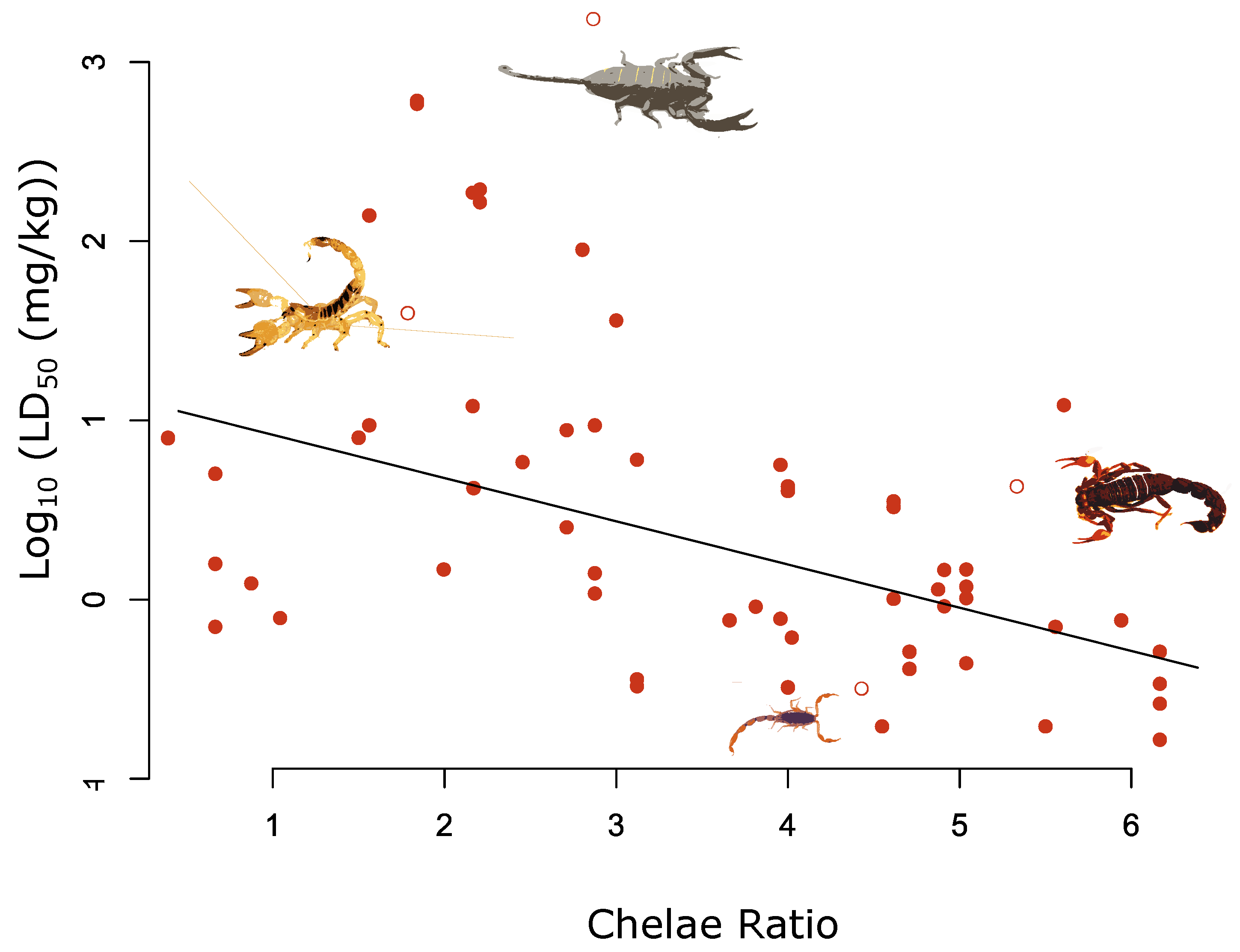
For the morphological traits, we found a significant negative relationship between log10 LD50 and chelae ratio, with species possessing more slender chelae being associated with lower log10 LD50 values (Figure 2 and Figure S1 and Table 1). Our results show that increasing the chelae ratio by one unit results in a 0.26 decrease in log10 LD50. Across the full range of chelae ratios in our dataset, this corresponds to approximately an order of magnitude decrease in LD50 (Figure 2 and Table 1).
For the telson ratio and the methods of injection for the LD50 test, we found no significant relationship with log10 LD50. The phylogenetic signal associated with LD50 was moderate to high in the analysis, with a h2 value of 0.60, and little variation associated with the species (Table 1).
3. Discussion
Here, we found strong support for the widely held perception that larger scorpions, with more robust chelae, are less potent than small species, with thin chelae. These results follow the general observations that the most potent scorpion venoms are recorded in smaller species, such as the bark scorpion Scorpion Centruroides noxius or the Brazilian yellow scorpion Tityus serrulatus. Conversely, the highest LD50 values, and, hence, the least potent scorpion venoms, are often found in some of the largest scorpion species. For example, in our data, the largest species, the rock scorpion Hadogenes granulatus, is associated with the least potent venom. Similarly, species with robust, well-developed chelae, such as the Israeli gold scorpion Scorpio maurus, were also associated with some of the least potent venoms. More potent venoms were found in species with narrow chelae, such as the South African thick-tail scorpion Parabuthus transvaalicus.
From an ecological perspective, these results support the patterns expected to arise from an evolutionary trade-off in investment between systems that can play similar defensive and predatory roles [16,84]. For prey capture, scorpions are observed to either primarily rely on mechanically capturing and subduing their prey with their pedipalps or only using them in a supportive role, with their venom primarily carrying out the function of incapacitation [85,86,87]. Such a trade-off is particularly apparent in species with wide chelae that can produce larger crushing forces, but have less-developed metasoma, such as in the rock scorpion Hadogenes granulatus.
Interestingly, we did not find a correlation between the shape of the telson, which houses the venom glands, and the venom potency. This highlights that, while there may be an evolutionary trade-off between chelae development and venom potency, this trade-off is not linked to the telson morphology. Instead, this suggests reduced selection in maintaining high levels of potency in larger species with robust chelae, but not in the ability to deliver venom. While snake species that no longer utilize venom for predation are found to have reduced potencies and, in some cases, are no longer able to deliver venom [88], this does not seem to be the case in scorpions. This retained ability to deliver venom in scorpion species that no longer use venom for predation likely points to the important defensive function of scorpion venoms. However, while our results do not support a trade-off between telson morphology and venom potency, other features of the scorpion delivery system may be associated with this trade-off, such as the metasoma. For example, Ref. [15] associated the thickness of the metasoma with venom potency. This may highlight the relationship between how frequently venom is used by a species and the fact that species that rely on venom for predation require more well-developed metasoma.
Our results highlight how ecological drivers can select for venom potency. However, the functional abilities of scorpion venoms are far more complex than the relatively one-dimensional measure of lethality using LD50. For example, it would be predicted that species with less-developed metasomas, such as the rock scorpion, may primarily retain venoms for defensive purposes, with such venoms no longer being selected for lethality, but for other attributes, such as inducing pain [89]. Conversely, while LD50 measures may be more appropriate to capture the functional ability of species that primarily rely on venom to incapacitate prey, other measures, such as the time taken to incapacitate, may be more ecologically relevant for future studies [90].
Further to the type of potency measure used, accounting for the species on which the venom is tested may also provide further understanding of potency variation across scorpion species. As the data used in our analysis consist of potency measures tested on mammalian models, more ecologically relevant test models that reflect each scorpion species’ diet may allow more detailed analyses of the drivers of scorpion venoms and composition to be conducted [26,91]. Studies have indicated some level of prey specificity in scorpion venom [92]; however, very few studies have tested scorpion venoms against their typical targets. The inclusion of such ecologically relevant test models would allow for the use of similar evolutionary distance metrics to those used in studies of snake venoms [26,93], to test and account for potency variation associated with prey specificity. Such models may also shed further light on the lack of a relationship found here between the route of venom injection and the potency. While it has been found, in studies of snake venoms, that the measures of LD50 using intravenous routes of injection are lower, the lack of such a relationship here may potentially be due to the fact that the scorpion venoms were selected to act on invertebrate prey, where the intravenous injection of venom was not selected for. However, as the potency measures used here were tested on mammalian model species, our results likely reflect the expected potency of scorpion venoms from a human medical perspective.
Our results also support the use of general scorpion morphology as a broad indicator for assessing the potential potency of a species. Such general indicators, particularly for clearly identifiable features, such as pedipalp size, may aid in envenomation cases where a species has not been identified, an issue that is common in many tropical and sub-tropical regions [94]. However, caution should be taken when using such general rules, as there is a large degree of variation in potency across the general patterns found in our analysis. For example, Scorpio maurus and Androctonus crassicauda are relatively similarly sized species, yet, despite Androctonus crassicauda being slightly larger, it is orders of magnitude more potent compared to Scorpio maurus. Hence, the use of these general indicators should be firmly based on more regional-specific information regarding the species that are likely to be associated with an envenomation event [95].
4. Conclusions
While potency is often the primary focus in understanding scorpionism risk, understanding the link between scorpion ecology and venom composition, and their associated envenomation symptoms [96], is likely to help in developing suitable envenomation remediation strategies. Testing fundamental patterns associated with the drivers of variation in scorpion venoms will not only allow for a clearer understanding of the ecology and evolution of venoms in scorpions, but will also provide a clearer path in understanding how to address the global issue of scorpionism.
5. Materials and Methods
To test our hypothesis, we collated data on venom potency and morphological features from the available literature. We performed an initial Web of Science search for scorpion potency and morphology data using the following search terms: “scorpion(s)”, “scorpion venom”, “scorpion venom potency”, “medically significant scorpions”, “lethality”, “median lethal dose”, “scorpion LD50”, “pedipalp measure”, “total body size”, “chela size”, “chela length”, “chela width”, “telson measure”, “telson length” and “telson width”. Further citations within key sources were also used in conjunction with the search terms.
For venom potency, we used median lethal dose (LD50), which was administered by intravenous (IV), subcutaneous (SC), intraperitoneal (IP) or intramuscular (IM) routes. We only included dried venom LD50 values, which reported the body mass of the test species and converted all units to mg of dried venom to kg of test subject.
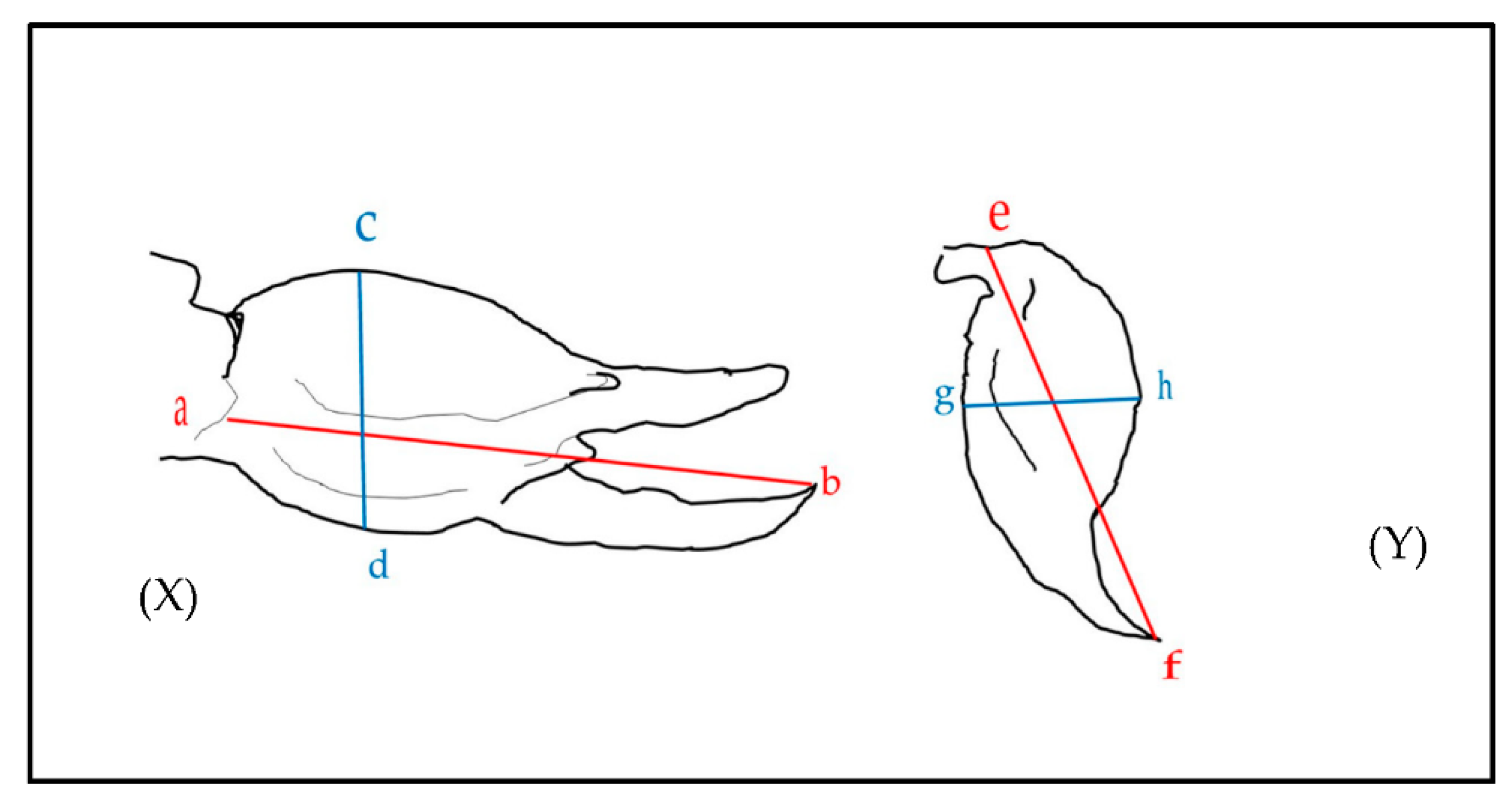
For morphological measurements, we used scorpion total body size (mm) and the length and width (mm) of the chela and telson from reported values and from diagrams and photos where scale bars were present. For measurements for the length of the chela, we used maximum distance from the tip of the tarsus (Figure 3X(a) to the base of the tibia, where it meets the distal end of the patella (Figure 3X(b). The width of the chela was measure at the widest points of the tibia (Figure 3X(c,d). For the telson, the length was measured as the maximum distance between the base of the vesicle (e) and the distal end of the aculeus (f), and the width was measured at the widest point of the vesicle (between points g and h).
As morphological measures were not available for Centruroides limpidus, we used values available from Centruroides ruana. These were described to have identical morphological features as each other.
For the analysis, we log10 transformed LD50 and body size values, as they resulted in a more normal distribution of model residuals. As body size is expected to be related to the length and width of both the chela and telson, we used length–width ratios to give mass-independent indices of their morphology. As short thick chelae are associated with delivering higher forces compared to long thick chelae [84,97], we divided the length of each chela by its width. Hence, this chela ratio ranged from high values, representing long thin chelae, to low values, representing short thick chelae. For the telson, we also divided the telson length by its dorsoventral width to give values ranging from high values, representing narrow elongated telsons, which are expected to be associated with a less-developed telson [98], to lower values, representing more bulbus telsons, which are expected to be associated with more developed use of venom [98].
To test our hypotheses, we fitted Bayesian phylogenetic mixed models (BPMM), using the MCMCglmm package [99] in R version 4.0.4 (Team 2016). These models allow for the inclusion of multiple explanatory variables as fixed effects, and random effect terms, which can be used as variance terms [99]. We controlled for pseudoreplication, due to shared ancestry between species, by using the animal term, and for the phylogenetic relationships between scorpions from our dataset using a phylogeny from the Open Tree of Life project [100]. The animal term uses a distance matrix of the phylogenetic distance between species to control for the expected similarity in trait values [99]. We calculated the relative variance attributable to the animal term as h2 [101], which can be interpreted in a similar fashion to the phylogenetic lambda value. A h2 value close to 1 indicates a Brownian model of trait evolution, while a value close to zero indicates independence between trait values [101]. In order to include multiple measures of LD50 for each species in our analysis, we also used a random term for species, similar to previous comparative models of venom variation across taxonomic groups [26]. Significance for a fixed term is determined when the 95% credibility interval (CI) does not cross zero [99]; however, we also included pMCMC values, which are a Bayesian alternative to p-values, with significance interpreted when pMCMC < 0.05 [99].
We fitted all models using standard non-informative priors, with the burn-in, thinning and number of iterations determined to ensure effective sample sizes exceeded 1000 for all parameter estimates. We tested for convergence using the Gelman–Rubin statistic over three separate chains [102].
As we found LD50 and body length to have log-normal distributions, we log10 transformed these variables. Such a transformation also allows us to fit a power–law relationship between LD50 and body length of the form LD50 ~ (body length)a. Such power relationships are typically associated with scaling relationships with body size [103], and have also been found to be associated with body size and venom properties in studies of snake venom [26]. We tested for collinearity between the independent factors using a series of regressions, finding no significant correlations between them (Supplementary Figures S2 and S3). We controlled for the effect of route of injection by including it as a fixed factor (SC, IM, IV, and IP), along with the two random terms accounting for phylogenetic variation (“animal” term) and species-level variation (“species” term). In our model, we included log10 (LD50) as the response variable, with log10 (body length), chela ratio and telson ratio as the independent variables. All data and analysis code are available in the Supplementary Files S1–S3.
Supplementary Materials
The following supporting information can be downloaded at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/toxins14030219/s1, Figure S1: Three dimensional plot of the relationship between the two significant the independent effects of log10 transformed Body size and Chela ratio with log10 transformed LD50. (for 62 observations for 36 species); Figure S2: Relationship between Chela ratio and log10 body length demonstrating no significant relationship between the variables (Slope = −0.002, p = 0.89 for 62 observations for 36 species); Figure S3: Relationship between Telson ratio and log10 body length demonstrating no significant relationship between the variables (Slope = −0.01, p = 0.45 for 62 observations for 36 species); File S1: Dataset containing measures of LD50, morphological measures and references for the data used in the analysis; File S2: Script outlining the analysis; File S3: Phylogeny file required for comparative analysis.
Author Contributions
Conceptualization, K.H., M.M.D., A.J. and A.F.; methodology, K.H., M.M.D. and A.F.; software, K.H. and A.F.; formal analysis, K.H. and A.F.; data curation, A.J. and A.F.; writing—original draft preparation, A.F.; writing—review and editing, K.H., M.M.D., A.J. and A.F.; visualization, K.H. and A.F.; supervision, K.H. and M.M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used in the analysis can be found in the Supplementary File S1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chippaux, J.-P. Emerging options for the management of scorpion stings. Drug Des. Dev. Ther. 2012, 6, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hidan, M.A.; Touloun, O.; Boumezzough, A. An epidemiological study on scorpion envenomation in the Zagora oases (Morocco). J. Coast. Life Med. 2015, 3, 704–707. [Google Scholar] [CrossRef]
- Adawi, S.H.A.A.; Adawi, S.H.A.A.; Adawi, D.H.A.A. The Scorpion sting: Epidemiology, clinical symptoms, treatment, surveillance and reporting obstacles in the Salfit district (West Bank) (2014–2015). Int. J. Trop. Dis. Health 2016, 14, 1–10. [Google Scholar] [CrossRef]
- Elyamani, R.; Soulaymani, A.; Serheir, Z. Epidemiology of scorpion envenoming in the prefecture of Figuig, Morocco. Int. J. Med. Toxicol. Forensic Med. 2019, 9, 17–24. [Google Scholar]
- Santos, M.S.; Silva, C.G.; Neto, B.S.; Grangeiro Junior, C.R.; Lopes, V.H.; Teixeira Junior, A.G.; Bezerra, D.A.; Luna, J.V.; Cordeiro, J.B.; Junior, J.G.; et al. Clinical and Epidemiological Aspects of Scorpionism in the World: A Systematic Review. Wilderness Environ. Med. 2016, 27, 504–518. [Google Scholar] [CrossRef] [Green Version]
- Chippaux, J.P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef]
- Mohammed-Geba, K.; Obuid-Allah, A.H.; El-Shimy, N.A.; Mahbob, M.A.E.-M.; Ali, R.S.; Said, S.M. DNA Barcoding for Scorpion Species from New Valley Governorate in Egypt Reveals Different Degrees of Cryptic Speciation and Species Misnaming. Conservation 2021, 1, 228–240. [Google Scholar] [CrossRef]
- Khatony, A.; Abdi, A.; Fatahpour, T.; Towhidi, F. The epidemiology of scorpion stings in tropical areas of Kermanshah province, Iran, during 2008 and 2009. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 45. [Google Scholar] [CrossRef] [Green Version]
- Murray, K.A.; Martin, G.; Iwamura, T. Focus on snake ecology to fight snakebite. Lancet 2020, 395, e14. [Google Scholar] [CrossRef]
- Chippaux, J.-P.; Williams, V.; White, J. Snake venom variability: Methods of study, results and interpretation. Toxicon 1991, 29, 1279–1303. [Google Scholar] [CrossRef]
- Vaucel, J.-A.; Larréché, S.; Paradis, C.; Labadie, M.; Courtois, A.; Grenet, G.; Langrand, J.; Tournoud, C.; Nisse, P.; Gallart, J.-C. Relationship between scorpion stings events and environmental conditions in mainland France. J. Med. Entomol. 2021, 58, 2146–2153. [Google Scholar] [CrossRef]
- Indiana Jones Wiki. Available online: https://indianajones.fandom.com/wiki/Scorpion (accessed on 18 January 2022).
- Müller, G.; Modler, H.; Wium, C.; Veale, D. Scorpion sting in southern Africa: Diagnosis and management. Contin. Med. Educ. 2012, 30, 356–361. [Google Scholar]
- Lourenço, W.R. Scorpion incidents, misidentification cases and possible implications for the final interpretation of results. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 21. [Google Scholar] [CrossRef]
- Lourenço, W.R. The coevolution between telson morphology and venom glands in scorpions (Arachnida). J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200128. [Google Scholar]
- Van Der Meijden, A.; Lobo Coelho, P.; Sousa, P.; Herrel, A. Choose your weapon: Defensive behavior is associated with morphology and performance in scorpions. PLoS ONE 2013, 8, e78955. [Google Scholar] [CrossRef] [Green Version]
- Coelho, P.; Kaliontzopoulou, A.; Rasko, M.; van der Meijden, A. A ‘striking’relationship: Scorpion defensive behaviour and its relation to morphology and performance. Funct. Ecol. 2017, 31, 1390–1404. [Google Scholar] [CrossRef] [Green Version]
- Bergman, N.J. Scorpion sting in Zimbabwe. S. Afr. Med. J. 1997, 87, 163–167. [Google Scholar]
- Kovařík, F. A review of the genus Heterometrus Ehrenberg, 1828, with descriptions of seven new species (Scorpiones, Scorpionidae). Euscorpius 2004, 2004, 1–60. [Google Scholar] [CrossRef]
- Hoang, A.N.; Vo, H.D.; Vo, N.P.; Kudryashova, K.S.; Nekrasova, O.V.; Feofanov, A.V.; Kirpichnikov, M.P.; Andreeva, T.V.; Serebryakova, M.V.; Tsetlin, V.I. Vietnamese Heterometrus laoticus scorpion venom: Evidence for analgesic and anti-inflammatory activity and isolation of new polypeptide toxin acting on Kv1. 3 potassium channel. Toxicon 2014, 77, 40–48. [Google Scholar] [CrossRef]
- van der Valk, T.; van der Meijden, A. Toxicity of scorpion venom in chick embryo and mealworm assay depending on the use of the soluble fraction versus the whole venom. Toxicon 2014, 88, 38–43. [Google Scholar] [CrossRef]
- Touloun, O.; Slimani, T.; Boumezzough, A. Epidemiological survey of scorpion envenomation in southwestern Morocco. J. Venom. Anim. Toxins 2001, 7, 199–218. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Omran, M.A.A.; Abdel-Nabi, I.M.; Ueda, H.; McVean, A. Intraspecific variation in the Egyptian scorpion Scorpio maurus palmatus venom collected from different biotopes. Toxicon 2009, 53, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.R.; Northfield, T.D.; Daly, N.L.; Wilson, D.T. Venom costs and optimization in scorpions. Front. Ecol. Evol. 2019, 7, 196. [Google Scholar] [CrossRef] [Green Version]
- El-Abd Ahmed, A.; Hassan, M.H.; Rashwan, N.I.; Sayed, M.M.; Meki, A.-R.M. Myocardial injury induced by scorpion sting envenoming and evidence of oxidative stress in Egyptian children. Toxicon 2018, 153, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Healy, K.; Carbone, C.; Jackson, A.L. Snake venom potency and yield are associated with prey-evolution, predator metabolism and habitat structure. Ecol. Lett. 2019, 22, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Newlands, G. Scorpion defensive behaviour. Afr. Wildl. 1969, 23, 147–153. [Google Scholar]
- De Sousa, L.; Parrilla-Alvarez, P.; Quiroga, M. An epidemiological review of scorpion stings in Venezuela: The Northeastern region. J. Venom. Anim. Toxins 2000, 6, 128–166. [Google Scholar] [CrossRef]
- Dent, M.A.; Possani, L.D.; Ramírez, G.A.; Fletcher, P.L., Jr. Purification and characterization of two mammalian toxins from the venom of the Mexican scorpion Centruroides noxius Hoffmann. Toxicon 1980, 18, 343–350. [Google Scholar] [CrossRef]
- El-Asmar, M.; Ismail, M.; Osman, O. Immunological studies of scorpion (Buthus minax, L. Koch) venom. Toxicon 1973, 11, 9–14. [Google Scholar] [CrossRef]
- Habermehl, G.G. Arthropoda. In Venomous Animals and Their Toxins; Springer: Berlin/Heidelberg, Germany, 1981; pp. 21–91. [Google Scholar]
- Ismall, M.; Osman, O.; Gumaa, K.; Karrar, M. Some pharmacological studies with scorpion (Pandinus exitialis) venom. Toxicon 1974, 12, 75–78. [Google Scholar] [CrossRef]
- Latifi, M.; Tabatabai, M. Immunological studies on Iranian scorpion venom and antiserum. Toxicon 1979, 17, 617–620. [Google Scholar] [CrossRef]
- Lourenço, W.R.; Cloudsley-Thompson, J. Stridulatory apparatus and the evolutionary significance of sound production in Rhopalurus species (Scorpiones: Buthidae). J. Arid Environ. 1995, 4, 423–429. [Google Scholar] [CrossRef]
- Marinkelle, C.; Stahnke, H. Toxicological and clinical studies on Centruroides margaritatus (Gervais), a common scorpion in western Colombia. J. Med. Entomol. 1965, 2, 197–199. [Google Scholar] [CrossRef]
- Nishikawa, A.; Caricati, C.P.; Lima, M.; Dos Santos, M.; Kipnis, T.; Eickstedt, V.; Knysak, I.; Da Silva, M.; Higashi, H.G.; Da Silva, W.D. Antigenic cross-reactivity among the venoms from several species of Brazilian scorpions. Toxicon 1994, 32, 989–998. [Google Scholar] [CrossRef]
- Ozkan, O.; Yagmur, E.; Ark, M. A newly described scorpion species, Leiurus abdullahbayrami (Scorpion: Buthidae), and the lethal potency and in vivo effects of its venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 414–421. [Google Scholar]
- Tu, A.T. Handbook of Natural Toxins: Insect poisons, allergens, and other invertebrate venoms. In Handbook of Natural Toxins: Insect Poisons, Allergens, and Other Invertebrate Venoms; Marcel Dekker Inc.: New York, NY, USA, 1984. [Google Scholar]
- Vazirianzadeh, B.; Fatoallahi, I.; Sharififard, M.; Moghadam, A.T. Determination of scorpion venom LD50 of Apistobuthus susanae species in Khuzestan province;(Southwest of Iran). Toxicon 2019, 159, S24. [Google Scholar] [CrossRef]
- Watt, D.D.; Simard, J.M. Neurotoxic proteins in scorpion venom. J. Toxicol. Toxin Rev. 1984, 3, 181–221. [Google Scholar] [CrossRef]
- Whittemore, F., Jr.; Keegan, H.; Borowitz, J. Studies of scorpion antivenins: 1. Paraspecificity. Bull. World Health Organ. 1961, 25, 185. [Google Scholar]
- Yağmur, E.A.; Özkan, Ö.; Karaer, K.Z. Determination of the median lethal dose and electrophoretic pattern of Hottentotta saulcyi (Scorpiones, Buthidae) scorpion venom. J. Arthropod-Borne Dis. 2015, 9, 238. [Google Scholar]
- Zlotkin, E.; Fraenkel, G.; Miranda, F.; Lissitzky, S. The effect of scorpion venom on blowfly larvae—A new method for the evaluation of scorpion venoms potency. Toxicon 1971, 9, 1–2. [Google Scholar] [CrossRef]
- Zlotkin, E.; Martinez, G.; Rochat, H.; Miranda, F. A protein from scorpion venom toxic to crustaceans. Anim. Plant Microb. Toxins 1976, 1, 73–80. [Google Scholar]
- Aboshaala, F.; Badry, A.; Sadine, S.E. Ecological considerations on Buthiscus bicalcaratus Birula, 1905 with a new locality in northern Libya (Scorpiones, Buthidae). Arch. L’Institut Pasteur D’Algérie 2020, 15, 97–101. [Google Scholar]
- Da Rosa, C.M.; Abegg, A.D.; Borges, L.M.; Bitencourt, G.S.; Di Mare, R.A. New record and occurrence map of Tityus serrulatus Lutz & Mello, 1922 (Scorpiones, Buthidae) in the state of Rio Grande do Sul, southern Brazil. Check List 2015, 11, 1556. [Google Scholar]
- De Armas, L.F.; Martín-Frías, E. El género" Centruroides" Marx, 1890 (Scorpiones: Buthidae) en el estado de Veracruz, México. Boletín SEA 2008, 43, 7–22. [Google Scholar]
- Fet, V.; Polis, G.A.; Sissom, W.D. Life in sandy deserts: The scorpion model. J. Arid Environ. 1998, 39, 609–622. [Google Scholar] [CrossRef]
- Fox, G.A.; Cooper, A.M.; Hayes, W.K. The dilemma of choosing a reference character for measuring sexual size dimorphism, sexual body component dimorphism, and character scaling: Cryptic dimorphism and allometry in the scorpion Hadrurus arizonensis. PLoS ONE 2015, 10, e0120392. [Google Scholar] [CrossRef] [Green Version]
- King, H. Notes on Sudan scorpions. Sudan Notes Rec. 1925, 8, 79–84. [Google Scholar]
- Kovařík, F. Taxonomic position of species of the genus Buthacus Birula, 1908 described by Ehrenberg and Lourenço, and description of a new species (Scorpiones: Buthidae). Euscorpius 2005, 2005, 1–13. [Google Scholar] [CrossRef]
- Kovařík, F. A revision of the genus Hottentotta Birula, 1908, with descriptions of four new species (Scorpiones, Buthidae). Euscorpius 2007, 2007, 1–107. [Google Scholar] [CrossRef]
- Kovařík, F.; Lowe, G.; Mazuch, T.; Awale, A.I.; Štundlová, J.; Šťáhlavský, F. Scorpions of the Horn of Africa (Arachnida: Scorpiones). Part XII. Pandinurus hangarale sp. n. (Scorpionidae) from Somaliland and a review of type locality and true distribution of Pandinurus smithi (Pocock, 1897). Euscorpius 2017, 253, 1–18. [Google Scholar] [CrossRef]
- Lourenço, W.; Huber, D.; Cloudsley-Thompson, J. Notes on the ecology, distribution and postembryonic development of Tityus cambridgei Pocock, 1897 (Scorpiones, Buthidae) from French Guyana and Oriental Amazonia. Entomol. Mitt. Aus Dem Zool. Mus. Hambg. 2000, 13, 197–203. [Google Scholar]
- Lourenço, W.R. Révision du Genre Rhopalurus Thorell, 1876 (Scorpiones, Buthidae). Available online: https://www.european-arachnology.org/esa/wp-content/uploads/2015/08/129-137_Lourenco.pdf (accessed on 3 February 2022).
- Lourenco, W.R. Compléments à la faune de scorpions (Arachnida) de l’Afrique du Nord, avec des considérations sur le genre Buthus Leach, 1815. Rev. Suisse Zool. 2003, 110, 875–912. [Google Scholar] [CrossRef]
- Lourenço, W.R. Nouvelles considérations taxonomiques sur les espèces du genre Androctonus Ehrenberg, 1828 et description de deux nouvelles espèces (Scorpiones, Buthidae). Rev. Suisse Zool. 2005, 112, 145–171. [Google Scholar] [CrossRef]
- Lourenço, W.R. What do we know about some of the most conspicuous scorpion species of the genus Tityus? A historical approach. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pipelzadeh, M.H.; Jalali, A.; Taraz, M.; Pourabbas, R.; Zaremirakabadi, A. An epidemiological and a clinical study on scorpionism by the Iranian scorpion Hemiscorpius lepturus. Toxicon 2007, 50, 984–992. [Google Scholar] [CrossRef]
- Prendini, L. The systematics of southern African Parabuthus Pocock (Scorpiones, Buthidae): Revisions to the taxonomy and key to the species. J. Arachnol. 2004, 32, 109–187. [Google Scholar] [CrossRef]
- Quijano-Ravell, A.F.; Ponce-Saavedra, J. A new species of scorpion of the genus Centruroides (Scorpiones: Buthidae) from the state of Michoacán, Mexico. Rev. Mex. Biodivers. 2016, 87, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Salama, W.M.; Sharshar, K.M. Surveillance study on scorpion species in Egypt and comparison of their crude venom protein profiles. J. Basic Appl. Zool. 2013, 66, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Seiter, M.; Turiel, C. First record of Androctonus australis (Linnaeus, 1758) from Jordan (Scorpiones: Buthidae). Rev. Ibérica Aracnol. 2013, 23, 95–98. [Google Scholar]
- Teruel, R.; Ponce-Saavedra, J.; Quijano-Ravell, A.F. Redescription of Centruroides noxius and description of a closely related new species from western Mexico (Scorpiones: Buthidae). Rev. Mex. Biodivers. 2015, 86, 896–911. [Google Scholar] [CrossRef] [Green Version]
- Karataş, A.; Kürtüllü, M. Duplication of pedipalp segments in the scorpion Androctonus crassicauda (Olivier, 1807). Euscorpius 2006, 2006, 1–4. [Google Scholar] [CrossRef]
- Kovařík, F. Review of Tunisian species of the genus Buthus with descriptions of two new species and a discussion of Ehrenberg’s types (Scorpiones: Buthidae). Euscorpius 2006, 2006, 1–16. [Google Scholar] [CrossRef]
- Kovařík, F.; Yağmur, E.A.; Fet, V.; Navidpour, S. On two subspecies of Mesobuthus eupeus (CL Koch, 1839) in Turkey (Scorpiones: Buthidae). Euscorpius 2011, 2011, 1–15. [Google Scholar]
- Lourenço, W.R. Further considerations on the genus Buthacus Birula, 1908. Boletín Soc. Entomol. Aragonesa 2006, 1, 59–70. [Google Scholar]
- Lourenço, W.R.; Cloudsley-Thompson, J.L. A new species of Buthus Leach, 1815 from Egypt (Scorpiones, Buthidae). Entomol. Mitt. Aus Dem Zool. Mus. Hambg. 2012, 16, 11–18. [Google Scholar]
- Lourenço, W.R.; Rossi, A. Sicily (Scorpiones, Buthidae). Biogeographical Implications. Available online: http://sea-entomologia.org/PDF/RIA22/009014RIA22ButhusSicilia.pdf (accessed on 3 February 2022).
- Booncham, U.; Sitthicharoenchai, D.; Pradatsundarasar, A.-O.; Prasarnpun, S.; Thirakhupt, K. Sexual dimorphism in the Asian giant forest scorpion, Heterometrus laoticus Couzijn, 1981. NU Int. J. Sci. 2007, 4, 42–52. [Google Scholar]
- Bucaretchi, F.; Fernandes, L.C.; Fernandes, C.B.; Branco, M.M.; Prado, C.C.; Vieira, R.J.; De Capitani, E.M.; Hyslop, S. Clinical consequences of Tityus bahiensis and Tityus serrulatus scorpion stings in the region of Campinas, southeastern Brazil. Toxicon 2014, 89, 17–25. [Google Scholar] [CrossRef]
- de Armas, L.F.; Teruel, R.; Kovařík, F. On Centruroides margaritatus (Gervais, 1841) and closely related species (Scorpiones: Buthidae). Euscorpius 2011, 2011, 1–16. [Google Scholar] [CrossRef]
- De Souza, C.A.R.; Candido, D.M.; Lucas, S.M.; Brescovit, A.D. On the Tityus stigmurus complex (Scorpiones, Buthidae). Zootaxa 2009, 1987, 1–38. [Google Scholar] [CrossRef]
- Esposito, L.A.; Yamaguti, H.Y.; Souza, C.A.; Pinto-Da-Rocha, R.; Prendini, L. Systematic revision of the neotropical club-tailed scorpions, Physoctonus, Rhopalurus, and Troglorhopalurus, revalidation of Heteroctenus, and descriptions of two new genera and three new species (Buthidae: Rhopalurusinae). Bull. Am. Mus. Nat. Hist. 2017, 2017, 1–136. [Google Scholar] [CrossRef]
- González-Santillán, E.; Galán-Sánchez, M.A.; Valdez-Velázquez, L.L. A new species of Centruroides (Scorpiones, Buthidae) from Colima, Mexico. Comptes Rendus Biol. 2019, 342, 331–344. [Google Scholar] [CrossRef]
- Mozaffari, E.; Sedaghat, M.M.; Dehkordi, A.S.; Akbarzadeh, K. Bidiversity and species composition of scorpions (Arachnida, Scorpiones) in Ilam County, Iran. J. Appl. Sci. Res. 2013, 9, 5412–5418. [Google Scholar]
- Navidpour, S.; Lowe, G. Revised diagnosis and redescription of Apistobuthus susanae (Scorpiones, Buthidae). J. Arachnol. 2009, 37, 45–59. [Google Scholar] [CrossRef]
- Prendini, L. Further additions to the scorpion fauna of Trinidad and Tobago. J. Arachnol. 2001, 29, 173–188. [Google Scholar] [CrossRef]
- Teruel, R.; de Armas, L.F. Redescripción de Rhopalurus junceus (Herbst 1800) (Scorpiones: Buthidae). Boletín Soc. Entomol. Aragonesa 2012, 50, 153–174. [Google Scholar]
- Teruel, R.; Kovařík, F. The true identity of the enigmatic scorpion Centruroides subgranosus (Kraepelin, 1898), with some taxonomic comments on Centruroides vittatus (Say, 1821) and Centruroides suffusus Pocock, 1902 (Scorpiones: Buthidae). Euscorpius 2010, 2010, 1–9. [Google Scholar] [CrossRef]
- Teruel, R.; Kovařík, F. Redescription of Androctonus bicolor Ehrenberg, 1828, and Description of Androctonus turieli sp. n. from Tunisia (Scorpiones: Buthidae). Euscorpius 2014, 186, 1–15. [Google Scholar] [CrossRef]
- Yağmur, E.A.; Koç, H.; Kunt, K.B. Description of a new species of Leiurus Ehrenberg, 1828 (Scorpiones: Buthidae) from southeastern Turkey. Euscorpius 2009, 2009, 1–20. [Google Scholar] [CrossRef]
- van der Meijden, A.; Kleinteich, T.; Coelho, P. Packing a pinch: Functional implications of chela shapes in scorpions using finite element analysis. J. Anat. 2012, 220, 423–434. [Google Scholar] [CrossRef]
- Williams, S. Scorpion bionomics. Annu. Rev. Entomol. 1987, 32, 275–295. [Google Scholar] [CrossRef]
- Edmunds, M.C.; Sibly, R.M. Optimal sting use in the feeding behavior of the scorpion Hadrurus spadix. J. Arachnol. 2010, 38, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Simone, Y.; Garcia, L.F.; Lacava, M.; van der Meijden, A.; Viera, C. Predatory versatility in females of the scorpion Bothriurus bonariensis (Scorpiones: Bothriuridae): Overcoming prey with different defensive mechanisms. J. Insect Behav. 2018, 31, 402–415. [Google Scholar] [CrossRef]
- Li, M.; Fry, B.; Kini, R.M. Eggs-only diet: Its implications for the toxin profile changes and ecology of the marbled sea snake (Aipysurus eydouxii). J. Mol. Evol. 2005, 60, 81–89. [Google Scholar] [CrossRef]
- Niermann, C.N.; Tate, T.G.; Suto, A.L.; Barajas, R.; White, H.A.; Guswiler, O.D.; Secor, S.M.; Rowe, A.H.; Rowe, M.P. Defensive Venoms: Is Pain Sufficient for Predator Deterrence? Toxins 2020, 12, 260. [Google Scholar] [CrossRef] [Green Version]
- Pekár, S.; Líznarová, E.; Bočánek, O.; Zdráhal, Z. Venom of prey-specialized spiders is more toxic to their preferred prey: A result of prey-specific toxins. J. Anim. Ecol. 2018, 87, 1639–1652. [Google Scholar] [CrossRef]
- Richards, D.P.; Barlow, A.; Wüster, W. Venom lethality and diet: Differential responses of natural prey and model organisms to the venom of the saw-scaled vipers (Echis). Toxicon 2012, 59, 110–116. [Google Scholar] [CrossRef]
- Van der Meijden, A.; Koch, B.; Van der Valk, T.; Vargas-Muñoz, L.J.; Estrada-Gómez, S. Target-specificity in scorpions; comparing lethality of scorpion venoms across arthropods and vertebrates. Toxins 2017, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Lyons, K.; Dugon, M.M.; Healy, K. Diet breadth mediates the prey specificity of venom potency in snakes. Toxins 2020, 12, 74. [Google Scholar] [CrossRef] [Green Version]
- Nejati, J.; Saghafipour, A.; Rafinejad, J.; Mozaffari, E.; Keyhani, A.; Abolhasani, A.; Kareshk, A.T. Scorpion composition and scorpionism in a high-risk area, the southwest of Iran. Electron. Physician 2018, 10, 7138. [Google Scholar] [CrossRef] [Green Version]
- El Hidan, M.A.; Touloun, O.; Bouazza, A.; Laaradia, M.A.; Boumezzough, A. Androctonus genus species in arid regions: Ecological niche models, geographical distributions, and envenomation risk. Vet. World 2018, 11, 286. [Google Scholar] [CrossRef]
- Ward, M.J.; Ellsworth, S.A.; Nystrom, G.S. A global accounting of medically significant scorpions: Epidemiology, major toxins, and comparative resources in harmless counterparts. Toxicon 2018, 151, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Van Der Meijden, A.; Herrel, A.; Summers, A. Comparison of chela size and pincer force in scorpions; getting a first grip. J. Zool. 2010, 280, 319–325. [Google Scholar] [CrossRef]
- van der Meijden, A.; Kleinteich, T. A biomechanical view on stinger diversity in scorpions. J. Anat. 2017, 230, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.D. MCMC methods for multi-response generalized linear mixed models: The MCMCglmm R package. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Hinchliff, C.E.; Smith, S.A.; Allman, J.F.; Burleigh, J.G.; Chaudhary, R.; Coghill, L.M.; Crandall, K.A.; Deng, J.; Drew, B.T.; Gazis, R. Synthesis of phylogeny and taxonomy into a comprehensive tree of life. Proc. Natl. Acad. Sci. USA 2015, 112, 12764–12769. [Google Scholar] [CrossRef] [Green Version]
- Hadfield, J.; Nakagawa, S. General quantitative genetic methods for comparative biology: Phylogenies, taxonomies and multi-trait models for continuous and categorical characters. J. Evol. Biol. 2010, 23, 494–508. [Google Scholar] [CrossRef]
- Brooks, S.P.; Gelman, A. General methods for monitoring convergence of iterative simulations. J. Comput. Graph. Stat. 1998, 7, 434–455. [Google Scholar]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
Figure 1.
Relationship between log10 total length (mm) and log10 LD50 (mg/kg) for 62 measures of LD50 across 36 species. The fitted line highlights the significant positive relationship between log10 total length and log10 LD50, adjusted for the median chelae ratio value of 3.7 (β = 3.24, pMCMC < 0.001). Selected species highlighted from left to right are highlighted by the hollow yellow circles and are as follows: Scorpion Centruroides noxius; Scorpio maurus; Androctonus crassicauda; Parabuthus transvaalicus; Hadogenes granulatus.
Figure 1.
Relationship between log10 total length (mm) and log10 LD50 (mg/kg) for 62 measures of LD50 across 36 species. The fitted line highlights the significant positive relationship between log10 total length and log10 LD50, adjusted for the median chelae ratio value of 3.7 (β = 3.24, pMCMC < 0.001). Selected species highlighted from left to right are highlighted by the hollow yellow circles and are as follows: Scorpion Centruroides noxius; Scorpio maurus; Androctonus crassicauda; Parabuthus transvaalicus; Hadogenes granulatus.

Figure 2.
Relationship between chelae and log10 LD50 (mg/kg) for 62 measures of LD50 across 36 species. The fitted line highlights the significant negative relationship between the chelae ratio value and log10 LD50 adjusted for the median log10 total length value of 70 mm (β = −0.26, pMCMC < 0.01). Selected species are highlighted by the hollow red circles and from left to right are as follows: Scorpio maurus; Hadogenes granulatus; Scorpion Centruroides noxius; Parabuthus transvaalicus.
Figure 2.
Relationship between chelae and log10 LD50 (mg/kg) for 62 measures of LD50 across 36 species. The fitted line highlights the significant negative relationship between the chelae ratio value and log10 LD50 adjusted for the median log10 total length value of 70 mm (β = −0.26, pMCMC < 0.01). Selected species are highlighted by the hollow red circles and from left to right are as follows: Scorpio maurus; Hadogenes granulatus; Scorpion Centruroides noxius; Parabuthus transvaalicus.

Figure 3.
Measurements taken from diagrams and photos with scale bars. (X) shows measurements for the length (a to b) and width (c to d) of the chela. (Y) shows the length (e to f) and width (g to h) of the telson. (a) represents the distal point where the pedipalpal patella meets the tibia; (b) is the most distal point of the pedipalpal tibia; (c) and (d) are the widest dorsoventral points on the pedipalpal tibia; (e) is the posterior end of the venom vesicle; (f) is the distal end of the aculeus; (g,h) are the widest dorsoventral points of the telson.
Figure 3.
Measurements taken from diagrams and photos with scale bars. (X) shows measurements for the length (a to b) and width (c to d) of the chela. (Y) shows the length (e to f) and width (g to h) of the telson. (a) represents the distal point where the pedipalpal patella meets the tibia; (b) is the most distal point of the pedipalpal tibia; (c) and (d) are the widest dorsoventral points on the pedipalpal tibia; (e) is the posterior end of the venom vesicle; (f) is the distal end of the aculeus; (g,h) are the widest dorsoventral points of the telson.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Main models testing the role of body size and morphology on LD50. The modes (β) and 95% credibility intervals (lower CI and upper CI) of the posterior distributions are given for all fixed and random terms in a model, with log10 of LD50 as the response variable. Fixed terms include the continuous factors log10 of scorpion total length, the ratio of chela length to width (chela ratio) and the ratio of telson length to width (telson ratio). Categorical fixed terms include the LD50 method of injection (subcutaneous (SC), intravenous (IV), intraperitoneal (IP) and intramuscular (IM)), with SC as the baseline. The random terms associated with phylogenetic relatedness (phylogeny (h2)), intraspecific variation (species) and residual variation (residual) are also presented. For more details on the parameters, see Materials and Methods. The model has 62 LD50 measures for 36 species.
Table 1.
Main models testing the role of body size and morphology on LD50. The modes (β) and 95% credibility intervals (lower CI and upper CI) of the posterior distributions are given for all fixed and random terms in a model, with log10 of LD50 as the response variable. Fixed terms include the continuous factors log10 of scorpion total length, the ratio of chela length to width (chela ratio) and the ratio of telson length to width (telson ratio). Categorical fixed terms include the LD50 method of injection (subcutaneous (SC), intravenous (IV), intraperitoneal (IP) and intramuscular (IM)), with SC as the baseline. The random terms associated with phylogenetic relatedness (phylogeny (h2)), intraspecific variation (species) and residual variation (residual) are also presented. For more details on the parameters, see Materials and Methods. The model has 62 LD50 measures for 36 species.
| β | Lower CI | Upper CI | pMCMC | |
|---|---|---|---|---|
| Fixed Terms | ||||
| Intercept | −4.82 | −8.40 | −1.68 | 0.004 |
| Log10 body length (mm) | 3.24 | 1.57 | 4.99 | <0.001 |
| LD50 methodSC | ||||
| IV | −0.04 | −0.35 | 0.29 | 0.81 |
| IP | 0.17 | −0.36 | 0.73 | 0.88 |
| IM | 0.10 | −1.05 | 1.35 | 0.52 |
| Chela ratio | −0.26 | −0.40 | −0.11 | 0.005 |
| Telson ratio | 0.04 | −0.15 | 0.20 | 0.63 |
| Random Terms | ||||
| Phylogeny (h2) | 0.60 | 0.25 | 0.87 | |
| Species | 0.01 | 0.00 | 0.46 | |
| Residuals | 0.26 | 0.10 | 0.49 |
Significant values, which are highlighted in bold, are deemed to be those with 95% of the posterior estimate above or below zero.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Forde, A.; Jacobsen, A.; Dugon, M.M.; Healy, K. Scorpion Species with Smaller Body Sizes and Narrower Chelae Have the Highest Venom Potency. Toxins 2022, 14, 219. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14030219
AMA Style
Forde A, Jacobsen A, Dugon MM, Healy K. Scorpion Species with Smaller Body Sizes and Narrower Chelae Have the Highest Venom Potency. Toxins. 2022; 14(3):219. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14030219
Chicago/Turabian StyleForde, Alannah, Adam Jacobsen, Michel M. Dugon, and Kevin Healy. 2022. "Scorpion Species with Smaller Body Sizes and Narrower Chelae Have the Highest Venom Potency" Toxins 14, no. 3: 219. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14030219
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.