Adsorption of Zearalenone by Aureobasidium pullulans Autolyzed Biomass Preparation and Its Detoxification Properties in Cultures of Saccharomyces cerevisiae Yeast



Abstract
:1. Introduction
2. Results
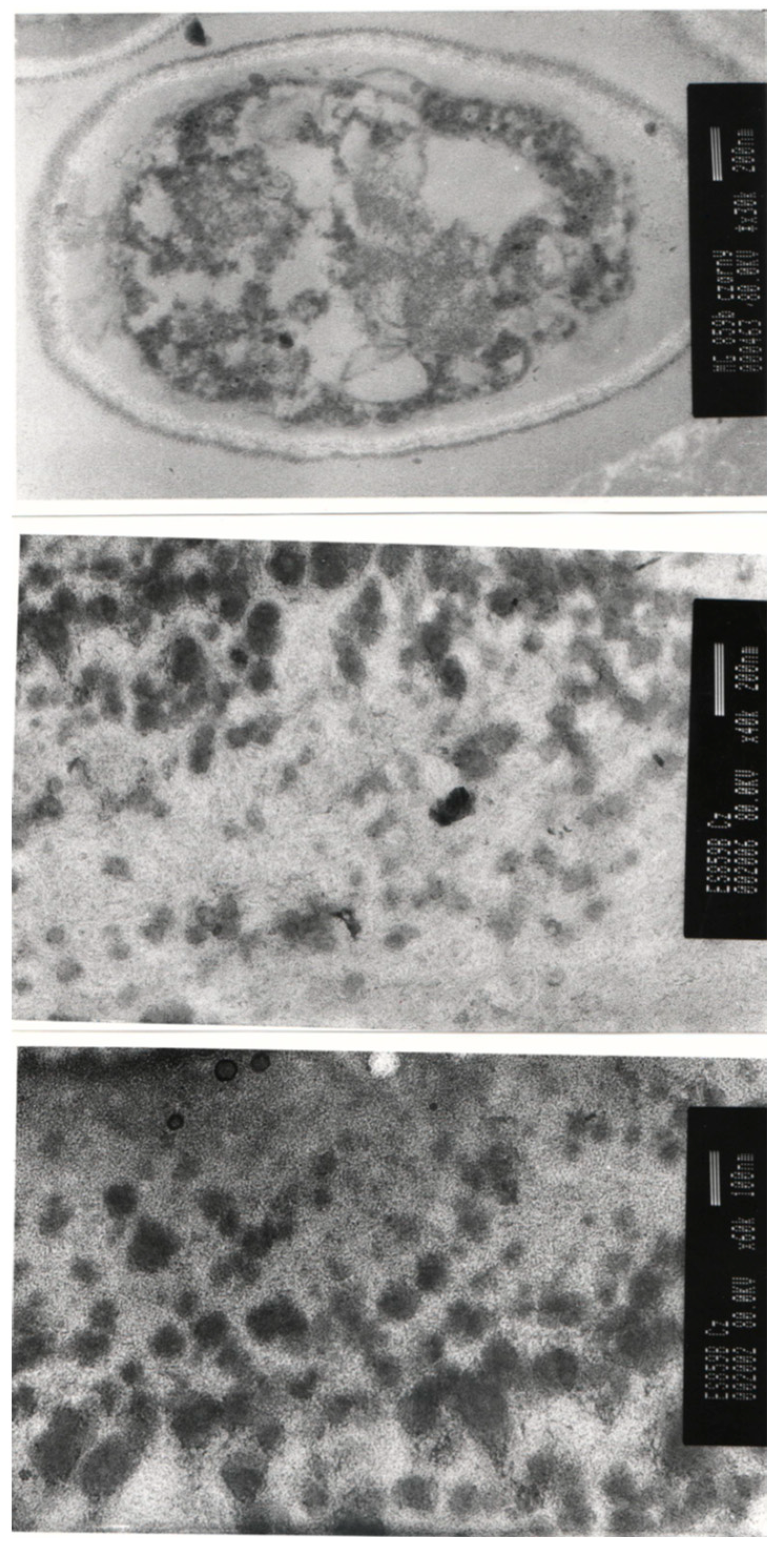
2.1. Chemical Composition of Autolyzed A. pullulans Biomass Preparation
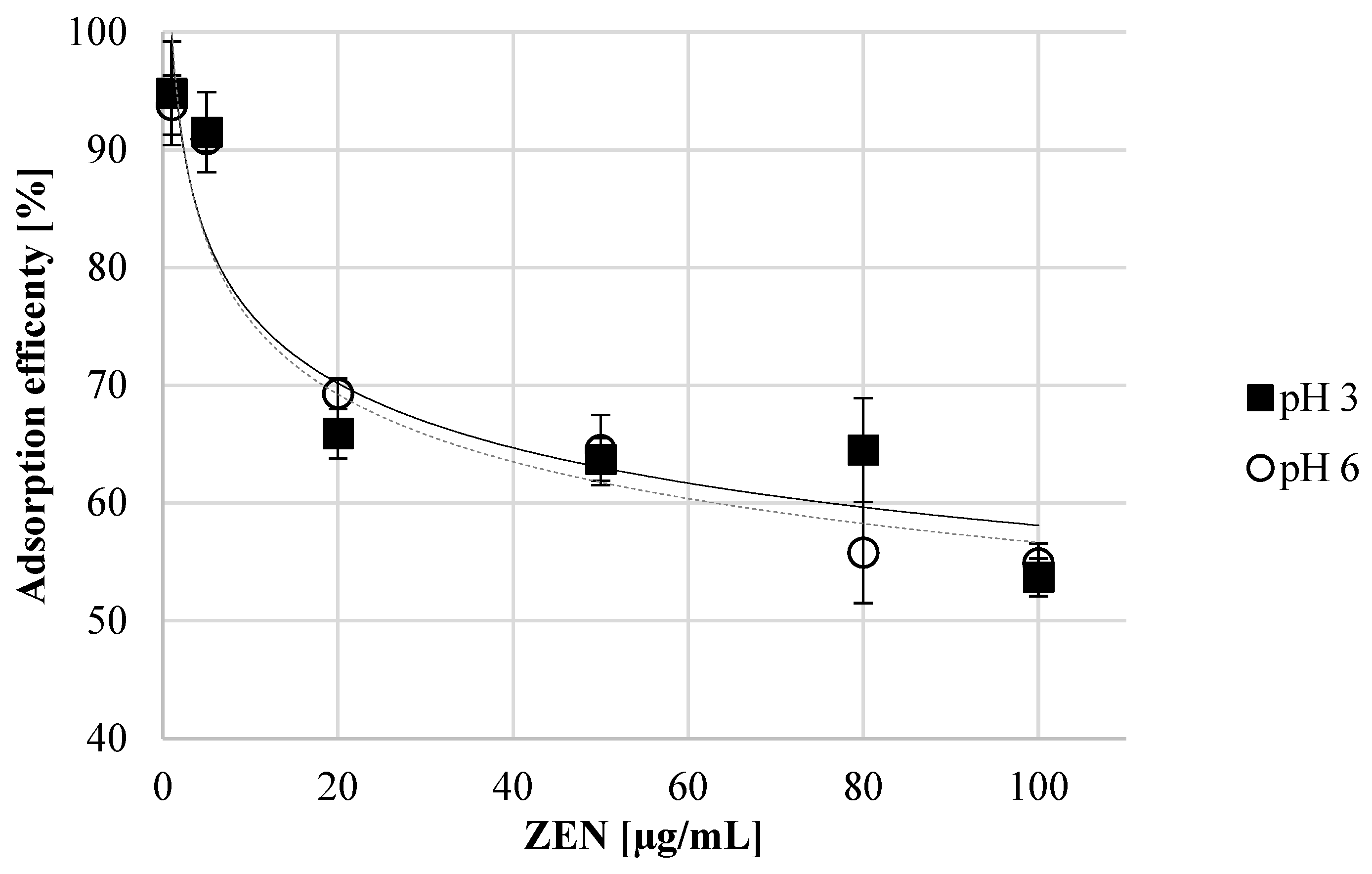
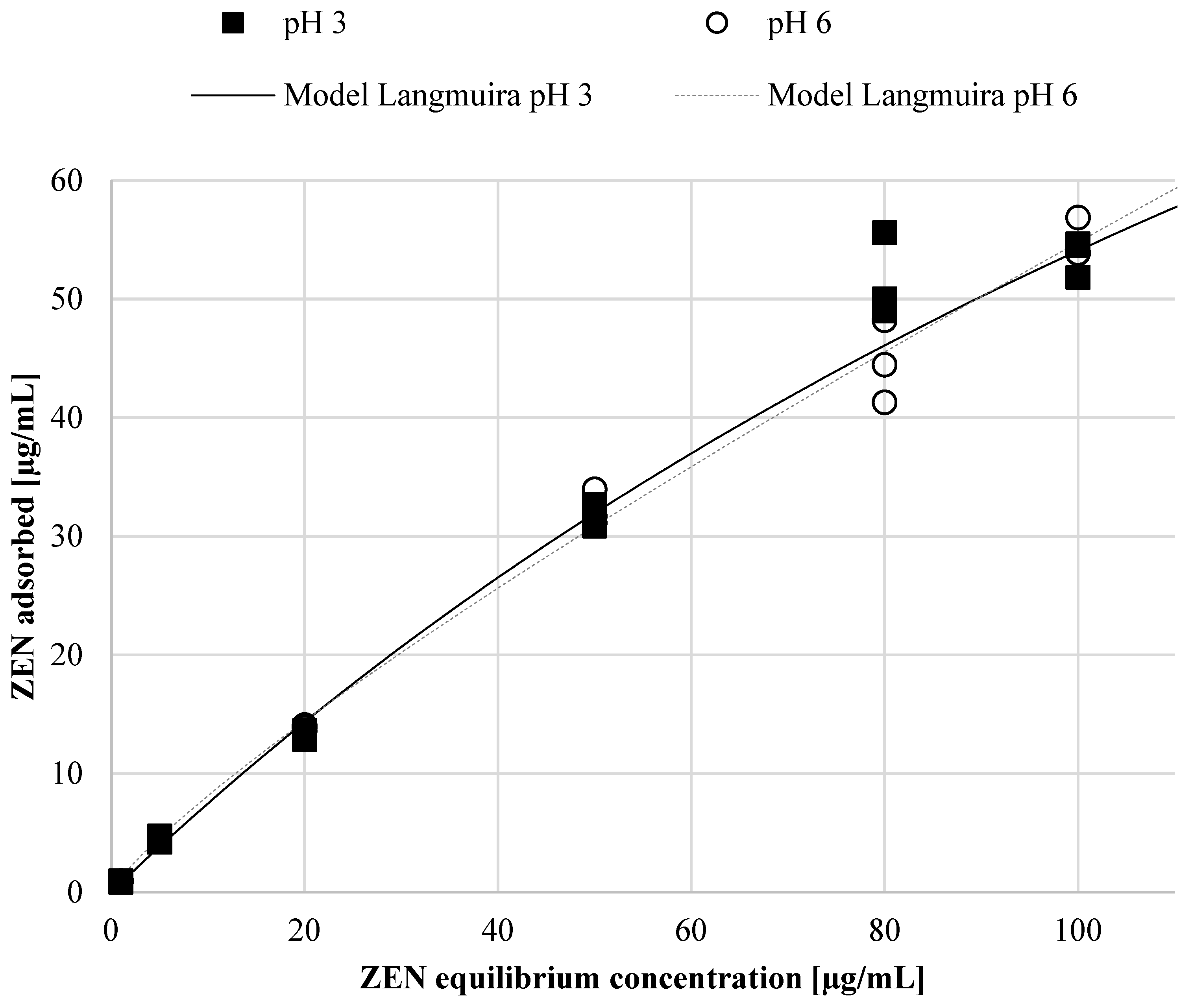
2.2. Determination of ZEN Adsorption Efficiency and Adsorption Isotherm Using Autolyzed Aureobasidium pullulans Biomass Preparation as Binding Material
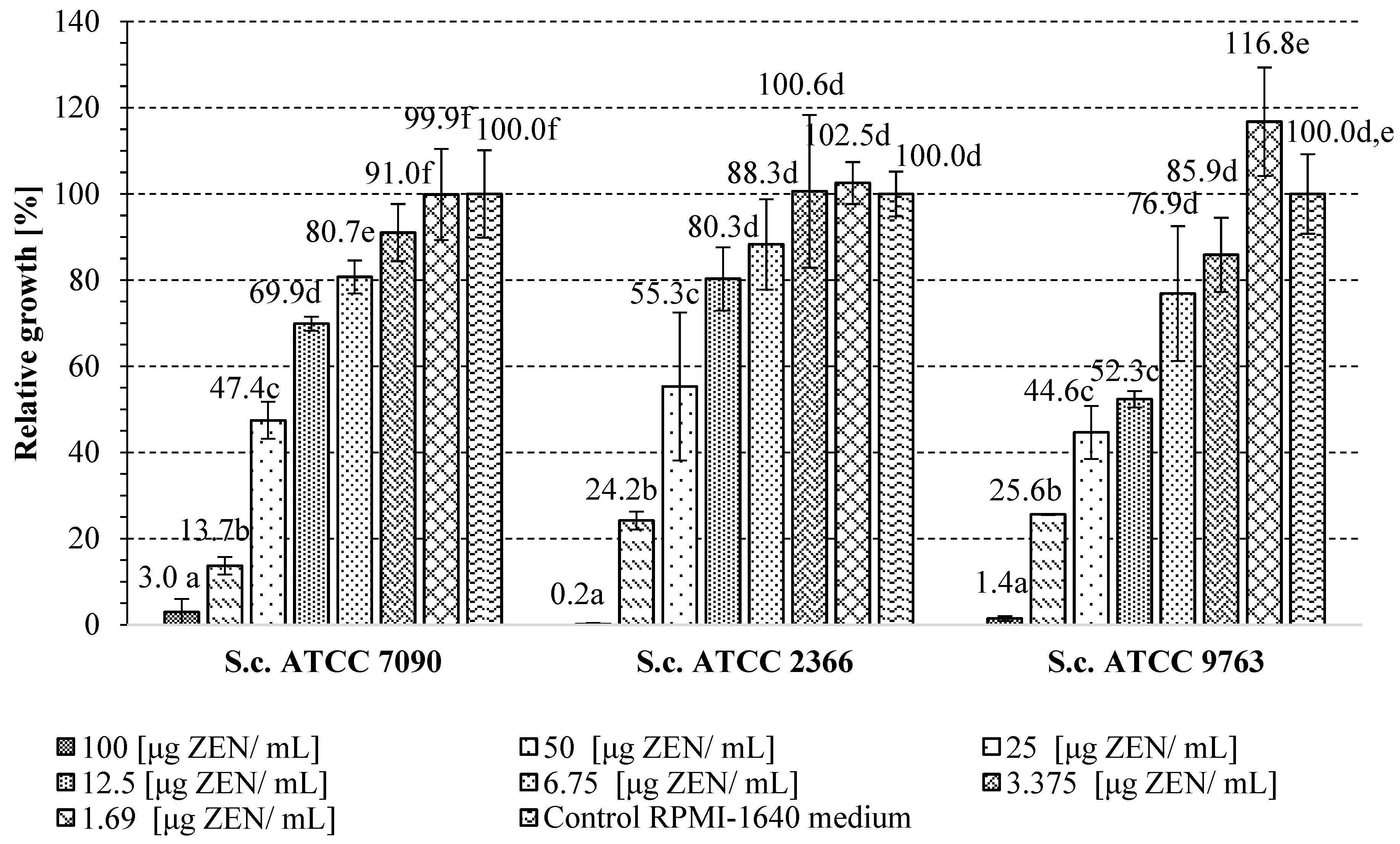
2.3. Determination of the Minimal Inhibitory Concentration of Zearalenone in Relation to S. cerevisiae Yeast Cells
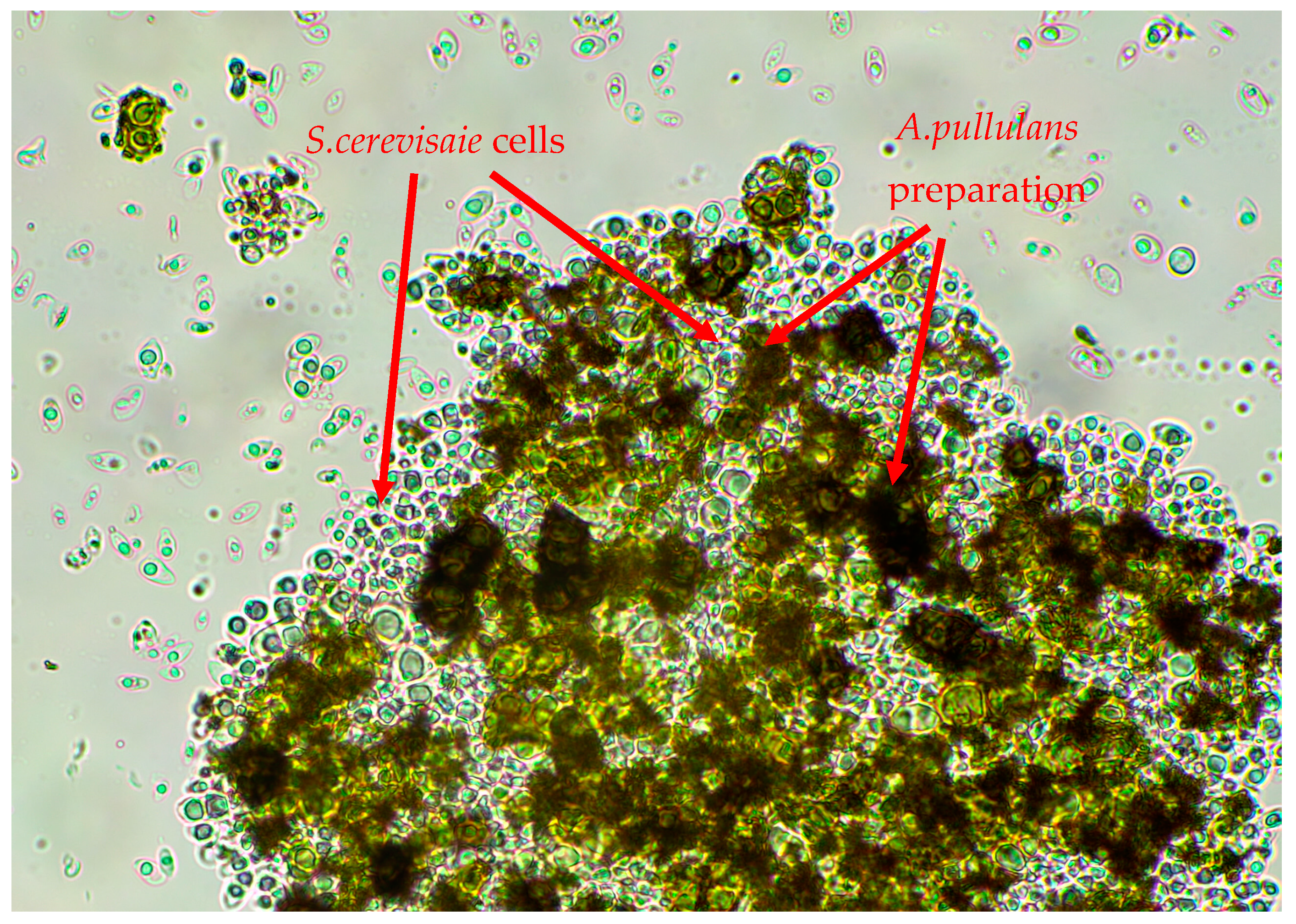
2.4. The Influence of A. pullulans Preparation on Yeast Growth in the Presence of Inhibitory ZEN Concentration
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Biological Material
5.2. A. pullulans Cultivation and Autolyzed Biomass Preparation
5.3. Chemical Characterization of the A. pullulans Autolyzed Biomass Preparation
5.4. Determination of ZEN Adsorption Isotherm Using Autolyzed A. pullulans Biomass Preparation as Binding Material
5.5. Determination of the Minimal Inhibitory Concentration of ZEN in Relation to S. cerevisiae Yeast Cells
5.6. Determination of the Effect of the A. pullulans Preparation on the Growth of Yeast in the Presence of an Inhibitory ZEN Concentration
5.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Annual Report of the Alert and Cooperation Network (RASFF, AAC and FFN). 2022. Available online: https://food.ec.europa.eu/system/files/2023-10/acn_annual-report_2022.pdf (accessed on 15 November 2023).
- Pinotti, L.; Ottoboni, M.; Giromini, C.; Dell’Orto, V.; Cheli, F. Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts. Toxins 2016, 8, 45. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Cai, P.; Liu, S.; Tu, Y.; Shan, T. Toxicity, biodegradation, and nutritional intervention mechanism of zearalenone. Sci. Total Environ. 2024, 911, 168648. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Ren Ch Gong, Y.; Gao, X.; Rajput, S.A.; Desheng, Q.; Shuai, W. The insensitive mechanism of poultry to zearalenone: A review. Anim. Nutr. 2021, 7, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2019, 60, 2710–2729. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Applegate, T. Zearalenone (ZEN) in Livestock and Poultry: Dose, toxicokinetics, toxicity and estrogenicity. Toxins 2020, 12, 377. [Google Scholar] [CrossRef] [PubMed]
- Commission Recommendation of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Feeding (2006/576/EC). Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:229:0007:0009:EN:PDF(accessed on 8 December 2023).
- European Union Commission Regulation 2023/915 on Maximum Levels for Certain Contaminants in Food and Repealing Regulation (EC) No. 1881/2006. Available online: https://eur-lex.europa.eu/eli/reg/2023/915/oj(accessed on 8 December 2023).
- EFSA Scientific Opinion. Appropriateness to set a group health-based guidance value for zearalenone and its modified forms. EFSA J. 2016, 14, 4425. [Google Scholar]
- Zhang, W.; Zhang, L.; Jiang, X.; Liu, X.; Li, Y.; Zhang, Y. Enhanced adsorption removal of aflatoxin B1, zearalenone and deoxynivalenol from dairy cow rumen fluid by modified nano-montmorillonite and evaluation of its mechanism. Anim. Feed Sci. Technol. 2020, 259, 114366. [Google Scholar] [CrossRef]
- Joannis-Cassan, C.; Tozlovanu, M.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Binding of Zearalenone, Aflatoxin B1, and Ochratoxin A by Yeast-Based Products: A Method for Quantification of Adsorption Performance. J. Food Prot. 2011, 74, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Armando, M.R.; Pizzolitto, R.P.; Dogi, C.A.; Cristofolini, A.; Merkis, C.; Poloni, V.; Dalcero, A.M.; Cavaglieri, L.R. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness. J. Appl. Microbiol. 2012, 113, 256–264. [Google Scholar] [CrossRef]
- de Felice, D.V.; Solfrizzo, M.; De Curtis, F.; Lima, G.; Visconti, A.; Castoria, R. Strains of Aureobasidium pullulans Can Lower Ochratoxin A Contamination in Wine Grapes. Phytopathology 2008, 98, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Andreasson, E.; Stenberg, J.A. Biological control of strawberry diseases by Aureobasidium pullulans and sugar beet extract under field conditions. J. Plant Pathol. 2023, 105, 933–941. [Google Scholar] [CrossRef]
- Singh, R.; Pandey, D.P.; Jamal, F.; Tiwari, S. Aureobasidium pullulans—An industrially important pullulan producing black yeast. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 605–622. [Google Scholar]
- EFSA Panel on Biological Hazards (BIOHAZ). Update of the List of QPS-Recommended Biological Agents Intentionally Added to Food or Feed as Notified to EFSA 13: Suitability of Taxonomic Units Notified to EFSA Until September 2020. Available online: https://0-efsa-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/abs/10.2903/j.efsa.2021.6377 (accessed on 24 January 2024).
- Vanderwaeren, L.; Dok, R.; Voordeckers, K.; Nuyts, S.; Verstrepen, K.J. Saccharomyces cerevisiae as a model system for eukaryotic cell biology, from cell cycle control to DNA damage response. Int. J. Mol. Sci. 2022, 23, 11665. [Google Scholar] [CrossRef] [PubMed]
- Fortina, M.G.; Parini, C.; Nsengumulemyi, D.J. Glucoamylase and fungal biomass production from cassava by Aureobasidium pullulans LV 10. Ann. Di Microbiol. Ed. Enzimol. 1993, 43, 91–101. [Google Scholar]
- Sevilla, M.J.; Moragues, M.D.; Uruburu, F. Growth and macromolecular content of the dimorphic fungus Aureobasidium pullulans and the effect of hydroxyurea and other inhibitors. Antonie Van Leeuwenhoek 1988, 54, 75–84. [Google Scholar] [CrossRef]
- Chi, Z.M.; Yan, K.R.; Gao, L.M.; Li, J.; Wang, X.H.; Wang, L. Diversity of marine yeasts with high protein content and evaluation of their nutritive compositions. J. Mar. Biol. Assoc. 2008, 88, 1347–1352. [Google Scholar] [CrossRef]
- Bzducha-Wróbel, A.; Błażejak St Kawarska, A.; Stasiak-Różańska, L.; Gientka, I.; Majewska, E. Evaluation of the efficiency of different disruption methods on yeast cell wall preparation for β-glucan isolation. Molecules 2014, 19, 20941–20961. [Google Scholar] [CrossRef]
- Kapteyn, J.C.; Van Den Ende, H.; Klis, F.M. The contribution of cell wall proteins to the organization of the yeast cell wall. Biochim. Biophys. Acta 1999, 1426, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Yiannikouris, A.; Shandilya, U.K.; Karrow, N.A. Comparative Assessment of Different Yeast Cell Wall-Based Mycotoxin Adsorbents Using a Model- and Bioassay-Based In Vitro Approach. Toxins 2023, 15, 104. [Google Scholar] [CrossRef]
- Brown, R.G.; Hanic, L.A.; Hsiao, M. Structure and chemical composition of yeast chlamydospores of Aureobasidium pullulans. Can. J. Microbiol. 1973, 19, 163–168. [Google Scholar] [CrossRef]
- Bzducha-Wróbel, A.; Błażejak, S.; Kieliszek, M.; Pobiega, K.; Falana, K.; Janowicz, M. Modification of the cell wall structure of Saccharomyces cerevisiae strains during cultivation on waste potato juice water and glycerol towards biosynthesis of functional polysaccharides. J. Biotechnol. 2018, 281, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bruinenberg, P.G.; Castex, M. Evaluation of a Yeast Hydrolysate from a Novel Strain of Saccharomyces cerevisiae for Mycotoxin Mitigation using In Vitro and In Vivo Models. Toxins 2022, 14, 7. [Google Scholar] [CrossRef]
- Aguilar-Uscanga, B.; François, J.M. A study of the yeast cell wall composition and structure in response to growth conditions and mode of cultivation. Lett. Appl. Microbiol. 2003, 37, 268–274. [Google Scholar] [CrossRef]
- Kim, K.S.; Yun, H.S. Production of soluble β-glucan from the cell wall of Saccharomyces cerevisiae. Enzym. Microb. Technol. 2006, 39, 496–500. [Google Scholar] [CrossRef]
- Bzducha-Wróbel, A.; Błażejak, S.; Molenda, M.; Reczek, L. Biosynthesis of β(1,3)/(1,6)-glucans of cell wall of the yeast Candida utilis ATCC 9950 strains in the culture media supplemented with deproteinated potato juice water and glycerol. Eur. Food Res. Technol. 2015, 240, 1023–1034. [Google Scholar] [CrossRef]
- Schiavone, M.; Sieczkowski, N.; Castex, M.; Dague, E.; François, M.J. Effects of the strain background and autolysis process on the composition and biophysical properties of the cell wall from two different industrial yeasts. FEMS Yeast Res. 2015, 15, 2. [Google Scholar] [CrossRef]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar] [CrossRef]
- Jouany, J.P. Methods for preventing, decontaminating and minimizing the toxicity of mycotoxins in feeds. Anim. Feed Sci. Technol. 2007, 137, 342–362. [Google Scholar] [CrossRef]
- Jakopović, Ž; Hanousek Čiča, K.; Mrvčić, J.; Pucić, I.; Čanak, I.; Frece, J.; Markov, K. Properties and fermentation activity of industrial yeasts Saccharomyces cerevisiae, S. uvarum, Candida utilis and Kluyveromyces marxianus exposed to AFB1, OTA and ZEA. Food Technol. Biotechnol. 2018, 56, 208–217. [Google Scholar] [CrossRef]
- Wang, N.; Weiwei, W.; Jiawen, P.; Miao, L. Detoxification Strategies for Zearalenone Using Microorganisms: A Review. Microorganisms 2019, 7, 208. [Google Scholar] [CrossRef]
- Yiannikouris, A.; François, J.; Poughon, L.; Dussap, C.G.; Bertin, G.; Jeminet, G.; Jouany, J.P. Alkali extraction of β-D-glucans from S. cerevisiae cell wall and study of their adsorptive properties toward zearalenone. J. Agric. Food Chem. 2004, 52, 3666–3673. [Google Scholar] [CrossRef]
- Yiannikouris, A.; Kettunen, H.; Apajalahti, J.; Pennala, E.; Moran, C.A. Comparison of the sequestering properties of yeast cell wall extract and hydrated sodium calcium aluminosilicate in three in vitro models accounting for the animal physiological bioavailability of zearalenone. Food Addit. Contam. Part A. 2013, 30, 1641–1650. [Google Scholar] [CrossRef]
- Ghosh, T.K.; Haldar, S.; Bedford, M.R.; Muthusami, N.; Samanta, I. Assessment of yeast cell wall as replacements for antibiotic growth promoters in broiler diets: Effects on performance, intestinal histo-morphology and humoral immune responses. J. Anim. Physiol. Anim. Nutr. 2012, 96, 275–284. [Google Scholar] [CrossRef]
- Liu, N.; Wang, J.; Liu, Z.; Wang, Y.; Wang, J. Effect of supplemental yeast cell walls on growth performance, gut mucosal glutathione pathway, proteolytic enzymes and transporters in growing broiler chickens. J. Anim. Sci. 2018, 96, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Bzducha-Wróbel, A.; Bryła, M.; Gientka, I.; Błażejak, S.; Janowicz, M. Candida utilis ATCC 9950 Cell Walls and β(1,3)/(1,6)-Glucan Preparations Produced Using Agro-Waste as a Mycotoxins Trap. Toxins 2019, 11, 192. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ribot, J.L.; Casanova, M.; Martinez, J.P.; Sentandreu, R. Characterization of cell wall proteins of yeast and hydrophobic mycelial cells of Candida albicans. Infect. Immun. 1991, 59, 2324–2332. [Google Scholar] [CrossRef] [PubMed]
- Nakkeeran, E.; Rathna, R.; Viveka, R. Mechanism and action of Aureobasidium pullulans on biosorption of metals. In Waste Bioremediation; Varjani, S.J., Gnansounou, E., Gurunathan, B., Pant, D., Zakaria, Z.A., Eds.; Springer: Singapore, 2018; pp. 215–231. [Google Scholar]
- Gniewosz, M.; Sobczak, E.; Zielinski, W. Optimization of saccharose and ammonium sulfate concentrations for pullulan biosynthesis by Aureobasidium pullulans in batch culture. Pol. J. Food Nutr. Sci. 1997, 6, 61–68. [Google Scholar]
- Gniewosz, M.; Duszkiewicz-Reinhard, W. Comparative studies on pullulan synthesis, melanin synthesis and morphology of white mutant Aureobasidium pullulans B-1 and parent strain A.p.-3. Carbohydr. Polym. 2008, 72, 431–438. [Google Scholar] [CrossRef]
- Saber, W.E.I.A.; Ghoniem, A.A.; Al-Otibi, F.O.; El-Hersh, M.S.; Eldadamony, N.M.; Menaa, F.; Elattar, K.M. A comparative study using response surface methodology and artificial neural network towards optimized production of melanin by Aureobasidium pullulans AKW. Sci. Rep. 2023, 13, 13545. [Google Scholar] [CrossRef]
- Boeira, L.S.; Bryce, J.H.; Stewart, G.G.; Flannigan, B. Inhibitory effect of Fusarium mycotoxins on growth of brewing yeasts. 1. Zearalenone and fumonisin B1. J. Inst. Brew. 1999, 105, 366–374. [Google Scholar] [CrossRef]
- Boeira, L.S.; Bryce, J.H.; Stewart, G.G.; Flannigan, B. The effect of combinations of Fusarium mycotoxins (deoxynivalenol, zearalenone and fumonisin B1) on growth of brewing yeasts. J. Appl. Microbiol. 2000, 88, 388–403. [Google Scholar] [CrossRef]
- Dziuba, E.; Foszczyńska, B.; Stempniewicz, R. Effect of mycotoxins DAS, ZEA and OTA one the growth of brewing yeast. Pol. J. Food Nutr. Sci. 2007, 57, 123–129. [Google Scholar]
- Doi, K.; Uetsuka, K. Mechanisms of mycotoxin-induced neurotoxicity through oxidative stress-associated pathways. Int. J. Mol. Sci. 2011, 12, 5213–5237. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.K.; Kang, S.C. Aflatoxin B1 induces reactive oxygen species-dependent caspase-mediated apoptosis in normal human cells, inhibits Allium cepa root cell division, and triggers inflammatory response in zebrafish larvae. Sci. Total Environ. 2020, 737, 139704. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Kusano, K.; Kondo, N.; Nishikawa, K.; Kuge, T.; Ohno, N. Biological Activity of High-Purity β-1,3-1,6-Glucan Derived from the Black Yeast Aureobasidium pullulans: A Literature Review. Nutrients 2021, 13, 242–266. [Google Scholar] [CrossRef] [PubMed]
- Mike, N.; Papp, G.; Certik, M.; Czibulya, Z.; Kunsagi-Mate, S.; Ember, I.; Vagvolgyi, C.; Pesti, M.; Gazdag, Z. Regulation of cytotoxic, non-estrogenic, oxidative stress induced processes of zearalenone in the fission yeast Schizosaccharomyces pombe. Toxicon 2013, 73, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Righetti, L.; Bhandari, D.R.; Rolli, E.; Tortorella, S.; Bruni, R.; Dall’Asta, C.; Spengler, B. Mycotoxin uptake in wheat—Eavesdropping Fusarium presence for priming plant defenses or a trojan horse to weaken them? Front. Plant Sci. 2021, 12, 711389. [Google Scholar] [CrossRef] [PubMed]
- Szkudzińska, K.; Smutniak, I.; Rubaj, J.; Korol, W.; Bielecka, G. Method validation for determination of amino acids in feed by UPLC. Accredit. Qual. Assur. 2017, 22, 247–252. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Mass % |
|---|---|
| Total sugars | 62.5 ± 1.3 |
| Total glucans | 38.9 ± 1.9 |
| β(1,3)/(1,6)-glucan | 37.0 ± 2.3 |
| α-glucan | 1.9 ± 0.2 |
| Protein | 9.6 ± 0.8 |
| Amino acids [mass % of total amino acids] | |
| * Asp | 6.6 ± 1.1 |
| Thr | 4.6 ± 0.9 |
| Ser | 4.3 ± 0.4 |
| Glu | 5.6 ± 1.1 |
| Pro | 6.4 ± 2.9 |
| Gly | 5.0 ± 0.1 |
| Ala | 6.0 ± 0.4 |
| Val | 6.2 ±0.8 |
| Leu | 7.5 ± 0.0 |
| Tyr | 6.9 ± 0.8 |
| His | 22.4 ± 2.4 |
| Lys | 4.0 ± 0.5 |
| Arg | 14.5 ± 1.6 |
| Model | pH Conditions during Adsorption | Isotherm Parameters Depending on pH and Model | |||
|---|---|---|---|---|---|
| Langmuir | Qmax | KL | RL | R2 | |
| pH 3 | 189.9 | 0.0042 | 0.705 | 0.990 | |
| pH 6 | 188.2 | 0.0040 | 0.712 | 0.997 | |
| Hill | Qmax | nH | RL | R2 | |
| pH 3 | 109.7 | 1.228 | - | 0.981 | |
| pH 6 | 199.1 | 0.971 | - | 0.988 | |
| Freundlich | Qmax | KF | nF | R2 | |
| pH 3 | - | 1.232 | 1.202 | 0.988 | |
| pH 6 | - | 1.236 | 1.215 | 0.998 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bzducha-Wróbel, A.; Janowicz, M.; Bryła, M.; Grzesiuk, I. Adsorption of Zearalenone by Aureobasidium pullulans Autolyzed Biomass Preparation and Its Detoxification Properties in Cultures of Saccharomyces cerevisiae Yeast. Toxins 2024, 16, 105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16020105
Bzducha-Wróbel A, Janowicz M, Bryła M, Grzesiuk I. Adsorption of Zearalenone by Aureobasidium pullulans Autolyzed Biomass Preparation and Its Detoxification Properties in Cultures of Saccharomyces cerevisiae Yeast. Toxins. 2024; 16(2):105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16020105
Chicago/Turabian StyleBzducha-Wróbel, Anna, Monika Janowicz, Marcin Bryła, and Iga Grzesiuk. 2024. "Adsorption of Zearalenone by Aureobasidium pullulans Autolyzed Biomass Preparation and Its Detoxification Properties in Cultures of Saccharomyces cerevisiae Yeast" Toxins 16, no. 2: 105. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins16020105