Monitoring Insecticide Susceptibility in Aedes Aegypti Populations from the Two Biggest Cities, Ouagadougou and Bobo-Dioulasso, in Burkina Faso: Implication of Metabolic Resistance

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
2. Methods
2.1. Study Area
2.2. Mosquito Collection and Rearing
2.3. Insecticide Susceptibility Tests
2.4. Enzyme Activities
2.5. Statistical Analyses
3. Results
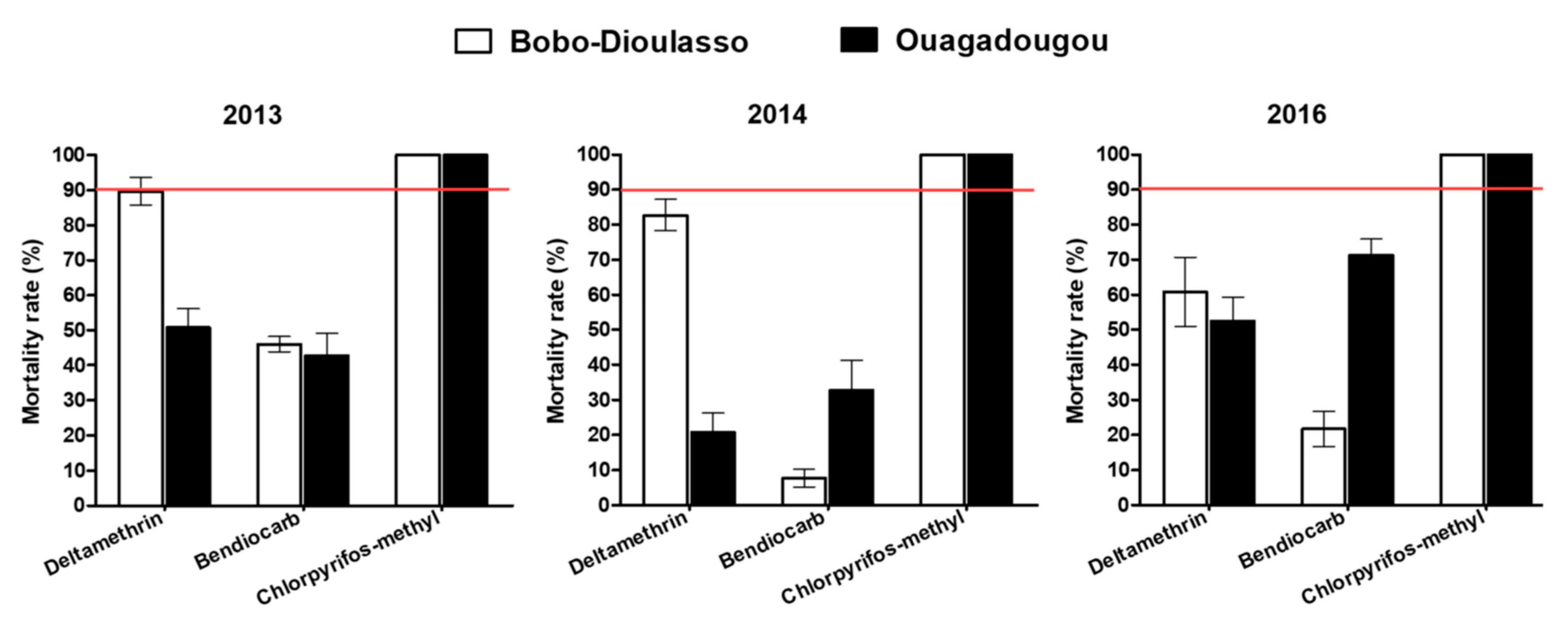
3.1. Insecticide Susceptibility Test
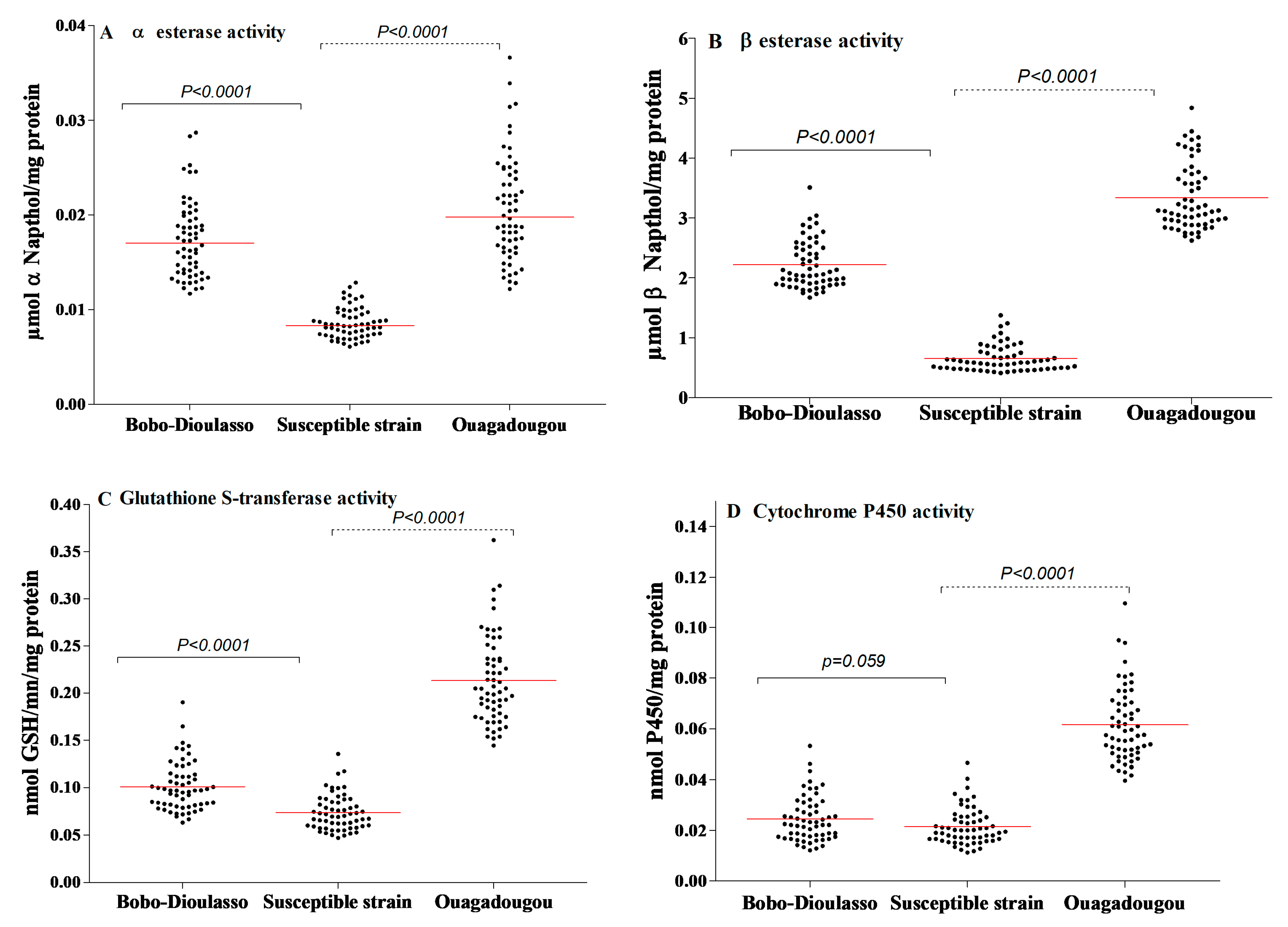
3.2. Non-Specific Esterases (NSE) and Para Nitro Phenyl Acetate (PNPA)
3.3. Gluthathione-S-Transferases (GST)
3.4. Oxidases (Cytochrome P450)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of Data and Materials
Abbreviations
| GST | Gluthatione S-transferase |
| kdr | Knockdown resistance |
| NS | Non-specific |
| NSE | Non-specific Esterase |
| PNPA | Para Nitro Phenyl Acetate |
| WHO | World Health Organization |
References
- WHO. Dengue au Burkina Faso; Bulletin d’information sur les flambées epidémiques 2016. Available online: https://www.who.int/csr/don/18-november-2016-dengue-burkina-faso/fr/ (accessed on 5 March 2020).
- Weetman, D.; Kamgang, B.; Badolo, A.; Moyes, C.L.; Shearer, F.M.; Coulibaly, M.; Pinto, J.; Lambrechts, L.; McCall, P.J. Aedes mosquitoes and Aedes-borne arboviruses in Africa: Current and future threats. Int. J. Environ. Res. Public Health 2018, 15, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, V.; Lhuillier, M.; Meunier, D.; Sarthou, J.L.; Monteny, N.; Digoutte, J.P.; Cornet, M.; Germain, M.; Cordellier, R. Yellow fever virus, dengue 2 and other arboviruses isolated from mosquitos, in Burkina Faso, from 1983 to 1986. Entomological and epidemiological considerations. Bull Soc Pathl Exot 1993, 86, 90–100. [Google Scholar]
- Ridde, V.; Agier, I.; Bonnet, E.; Carabali, M.; Dabiré, K.R.; Fournet, F.; Ly, A.; Meda, I.B.; Parra, B. Presence of three dengue serotypes in Ouagadougou (Burkina Faso): Research and public health implications. Infect. Dis. Poverty 2016, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlenker, E.H.; Goldman, M.; Holman, G. Effect of aspartic acid on control of ventilation in androgenized and ovariectomized female rats. J. Appl. Physiol. 1992, 72, 2255–2258. [Google Scholar] [CrossRef]
- Amarasinghe, A.; Kuritsky, J.N.; Letson, G.W.; Margolis, H.S. Dengue Virus Infection in Africa. Emerg. Infect. Dis. 2011, 17, 1349–1354. [Google Scholar] [CrossRef]
- Roux, V.R.J.; Saluzzo, M.L.J.F.; Sarthou, J.L.; Stanciheluni, A.; Molez, G.J.F.; Some, L.; Darriet, F.; Soudret, R.; Guiguemde, T.R.; Henneqiun, M. L’epidemie de fievre jaune au Burkina Faso en 1983. Bull World Heal. Organ 1986, 64, 873–882. [Google Scholar]
- Franco, L.; Di Caro, A.; Carletti, F.; Vapalahti, O.; Renaudat, C.; Zeller, H.; Tenorio, A. Recent expansion of dengue virus serotype 3 in West Africa. Euro Surveill. 2010, 15, 1–4. [Google Scholar]
- Eckerle, I.; Kapaun, A.; Junghanss, T.; Schnitzler, P.; Drosten, C.; Jänisch, T. Dengue virus serotype 3 infection in traveler returning from West Africa to Germany. Emerg. Infect. Dis. 2015, 21, 175. [Google Scholar] [CrossRef]
- Ouattara, L.P.E.; Sangaré, I.; Namountougou, M.; Hien, A.; Ouari, A.; Soma, D.D.; Kassié, D.; Diabaté, A.; Gnankiné, O.; Bonnet, E.; et al. Surveys of Arboviruses Vectors in Four Cities Stretching Along a Railway Transect of Burkina Faso: Risk Transmission and Insecticide Susceptibility Status of Potential Vectors. Front. Vet. Sci. 2019, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- NSDI. Projections Demographiques des Communes du Burkina Faso de 2007 à 2020; INSD. 2017. Available online: https://www.insd.bf (accessed on 5 March 2020).
- Mattingly, P.F.; Rageau, J.; La, C.A.; Des, F.; Du, M.; Xii, S.A. Contributions de la faune des moustiques du Sud-Est Asiatique. XII. Contrib. Am. Entomol. Inst. 1973, 7. [Google Scholar]
- Mouchet, J.; Bouchite, B. Distribution et gites pre-imaginaux de quelques espèces de moustiques africains presents en Haute Volta, 1964; in press.
- WHO. Guidelines for Laboratory and Field-Testing of Long-Lasting Insecticidal Nets; World Health Organization: Geneva, Switzerland, 2013; Who/Htm/Ntd/Whopes/2013.1. [Google Scholar]
- WHO. Techniques to Detect Insecticide Resistance Mechanisms (Field and Laboratory Manual); World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Namountougou, M.; Simard, F.; Baldet, T.; Diabaté, A.; Ouédraogo, J.B.; Martin, T.; Dabiré, R.K. Multiple Insecticide Resistance in Anopheles gambiae s.l. Populations from Burkina Faso, West Africa. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namountougou, M.; Soma, D.D.; Kientega, M.; Balboné, M.; Kaboré, D.P.A.; Drabo, S.F.; Coulibaly, A.Y.; Fournet, F.; Baldet, T.; Diabaté, A.; et al. Insecticide resistance mechanisms in Anopheles gambiae complex populations from Burkina Faso, West Africa. Acta Trop. 2019, 197, 105054. [Google Scholar] [CrossRef] [PubMed]
- WHO. Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes, 2nd ed.; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Tarnagda, Z.; Congo, M.; Sagna, T.; Ouédraogo, C.; Nikiéma, V.; Cissé, A.; Sanou, M.A.; Ouédraogo, A.; Bosco, J. Outbreak of dengue fever in Ouagadougou, Burkina Faso, 2013. Int. J. Microbiol. Immunol. Res. 2014, 2, 101–108. [Google Scholar]
- Ponlawat, A.; Scott, J.G.; Harrington, L.C. Insecticide susceptibility of Aedes aegypti and Aedes albopictus across Thailand. J. Med. Entomol. 2005, 42, 821–825. [Google Scholar] [CrossRef]
- Amelia-yap, Z.H.; Chen, C.D.; Sofian-azirun, M.; Low, V.L. Pyrethroid resistance in the dengue vector Aedes aegypti in Southeast Asia: Present situation and prospects for management. Parasit. Vectors 2018, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Marcombe, S.; Mathieu, R.B.; Pocquet, N.; Riaz, M.A.; Poupardin, R.; Sélior, S.; Darriet, F.; Reynaud, S.; Yébakima, A.; Corbel, V.; et al. Insecticide resistance in the dengue vector Aedes aegypti from martinique: Distribution, mechanisms and relations with environmental factors. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Ahouangninou, C.; Fayomi, B.E.; Martin, T. Evaluation des risques sanitaires et environnementaux des pratiques phytosanitaires des producteurs maraîchers dans la commune rurale de Tori-Bossito (Sud-Bénin). Cah. Agric. 2011, 20, 216–222. [Google Scholar]
- Bharati, M.; Saha, D. Multiple insecticide resistance mechanisms in primary dengue vector, Aedes aegypti (Linn.) from dengue endemic districts of sub-Himalayan West Bengal, India. PLoS ONE 2018, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Diabate, A.; Baldet, T.; Chandre, F.; Akogbeto, M.; Guiguemde, T.R.; Darriet, F.; Brengues, C.; Guillet, P.; Hemingway, J.; Small, G.J.; et al. The role of agricultural use of insecticides in resistance to pyrethroids in Anopheles gambiae s.l. in Burkina Faso. Am. J. Trop. Med. Hyg. 2002, 67, 617–622. [Google Scholar] [CrossRef] [Green Version]
- Dabiré, K.R.; Diabaté, A.; Namountougou, M.; Toé, K.H.; Ouari, A.; Kengne, P.; Bass, C.; Baldet, T. Distribution of pyrethroid and DDT resistance and the L1014F kdr mutation in Anopheles gambiae s.l. from Burkina Faso (West Africa). Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Kawada, H.; Higa, Y.; Futami, K.; Muranami, Y.; Kawashima, E.; Osei, J.H.N.; Sakyi, K.Y.; Dadzie, S.; de Souza, D.K.; Appawu, M.; et al. Discovery of Point Mutations in the Voltage-Gated Sodium Channel from African Aedes aegypti Populations: Potential Phylogenetic Reasons for Gene Introgression. PLoS Negl. Trop. Dis. 2016, 10, e0004780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Saavedra-Rodriguez, K.; Urdaneta-Marquez, L.; Rajatileka, S.; Moulton, M.; Flores, A.E.; Salas, F.-; Bisset, J.; Rodriguez, M.; Mccall, P.J.; Donnelly, M.J.; et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol Biol 2007, 16, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brengues, C.; Hawkes, N.J.; Chandre, F.; Mccarroll, L.; Duchon, S.; Guillet, P.; Manguin, S.; Morgan, J.C.; Hemingway, J. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol. 2003, 17, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Du, Y.; Nomura, Y.; Dong, K. A sodium channel mutation identified in Aedes aegypti selectively reduces cockroach sodium channel sensitivity to type I, but not type II pyrethroids. Insect Biochem. Mol. Biol. 2011, 4, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Nomura, Y.; Satar, G.; Hu, Z.; Nauen, R.; He, S.Y.; Zhorov, B.S.; Dong, K. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc. Natl. Acad. Sci. USA 2013, 110, 11785–11790. [Google Scholar] [CrossRef] [Green Version]
- Badolo, A.; Sombié, A.; Pignatelli, P.M.; Sanon, A.; Yaméogo, F.; Wangrawa, D.W.; Sanon, A.; Kanuka, H.; McCall, P.J.; Weetman, D. Insecticide resistance levels and mechanisms in Aedes aegypti populations in and around Ouagadougou, Burkina Faso. PLoS Negl. Trop. Dis. 2019, 13, e0007439. [Google Scholar] [CrossRef]
- Vontas, J.G.; Small, G.J.; Hemingway, J. Glutathione S-transferases as antioxidant defence agents confer pyrethroid resistance in Nilaparvata lugens. Biochem. J. 2001, 357, 65–72. [Google Scholar] [CrossRef]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Namountougou, M.; Soma, D.D.; Balboné, M.; Kaboré, D.A.; Kientega, M.; Hien, A.; Coulibaly, A.; Ouattara, P.E.; Meda, B.G.; Drabo, S.; et al. Monitoring Insecticide Susceptibility in Aedes Aegypti Populations from the Two Biggest Cities, Ouagadougou and Bobo-Dioulasso, in Burkina Faso: Implication of Metabolic Resistance. Trop. Med. Infect. Dis. 2020, 5, 84. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed5020084
Namountougou M, Soma DD, Balboné M, Kaboré DA, Kientega M, Hien A, Coulibaly A, Ouattara PE, Meda BG, Drabo S, et al. Monitoring Insecticide Susceptibility in Aedes Aegypti Populations from the Two Biggest Cities, Ouagadougou and Bobo-Dioulasso, in Burkina Faso: Implication of Metabolic Resistance. Tropical Medicine and Infectious Disease. 2020; 5(2):84. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed5020084
Chicago/Turabian StyleNamountougou, Moussa, Dieudonné Diloma Soma, Mahamoudou Balboné, Didier Alexandre Kaboré, Mahamadi Kientega, Aristide Hien, Ahmed Coulibaly, Parfait Eric Ouattara, Benson Georges Meda, Samuel Drabo, and et al. 2020. "Monitoring Insecticide Susceptibility in Aedes Aegypti Populations from the Two Biggest Cities, Ouagadougou and Bobo-Dioulasso, in Burkina Faso: Implication of Metabolic Resistance" Tropical Medicine and Infectious Disease 5, no. 2: 84. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed5020084