
Nationwide Distribution of Dengue Virus Type 3 (DENV-3) Genotype I and Emergence of DENV-3 Genotype III during the 2019 Outbreak in Bangladesh

 and
and
Abstract
:1. Introduction
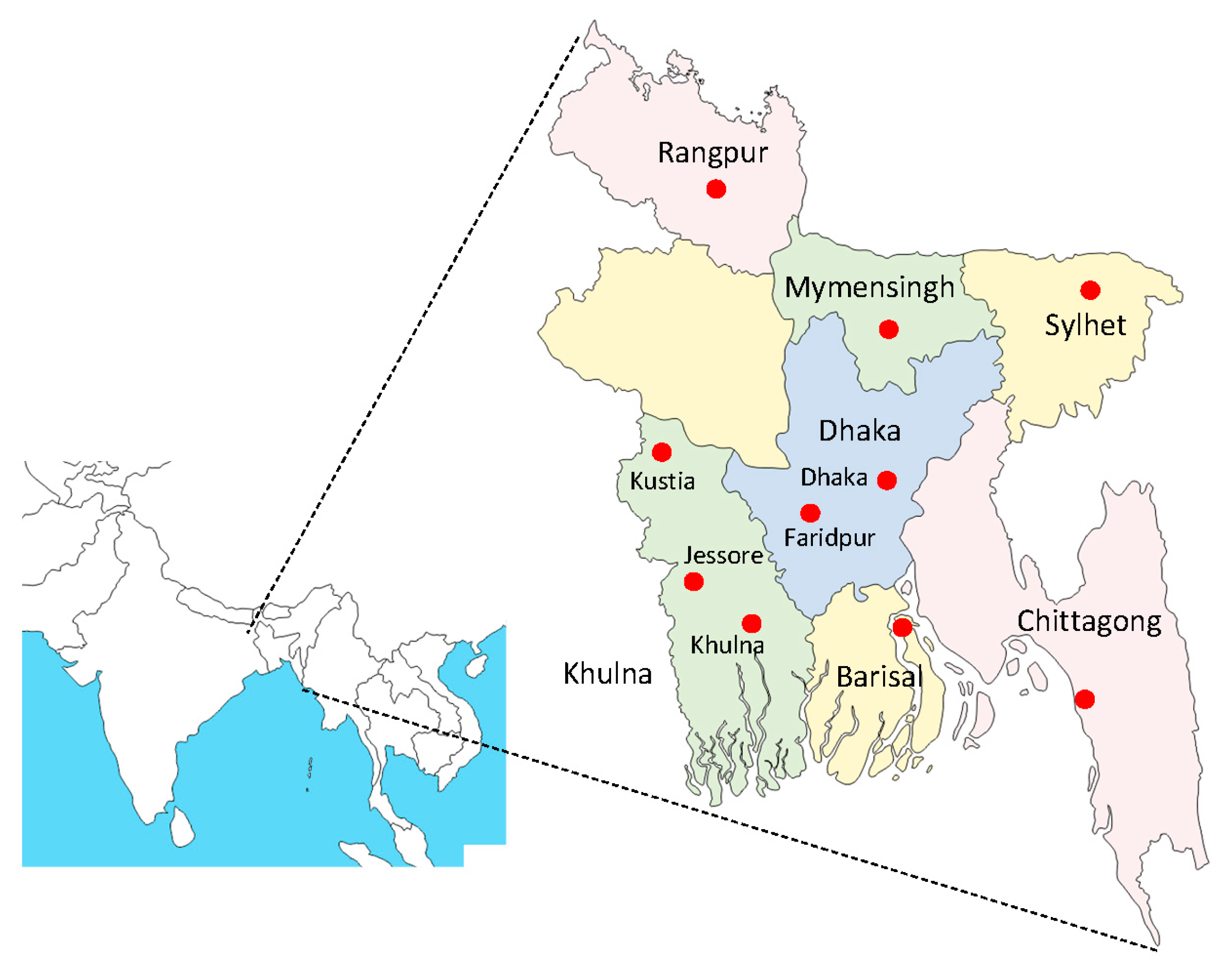
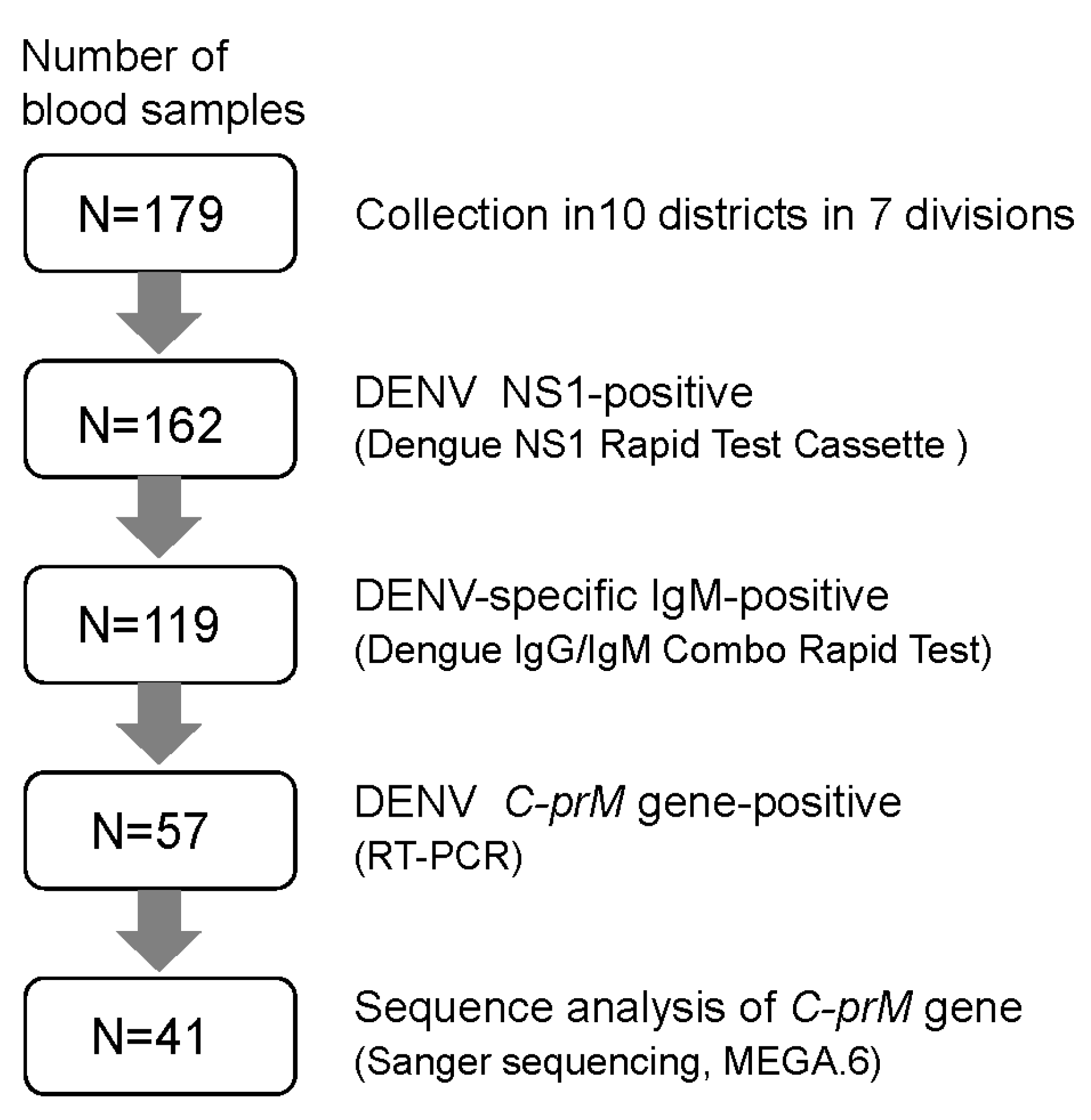
2. Materials and Methods
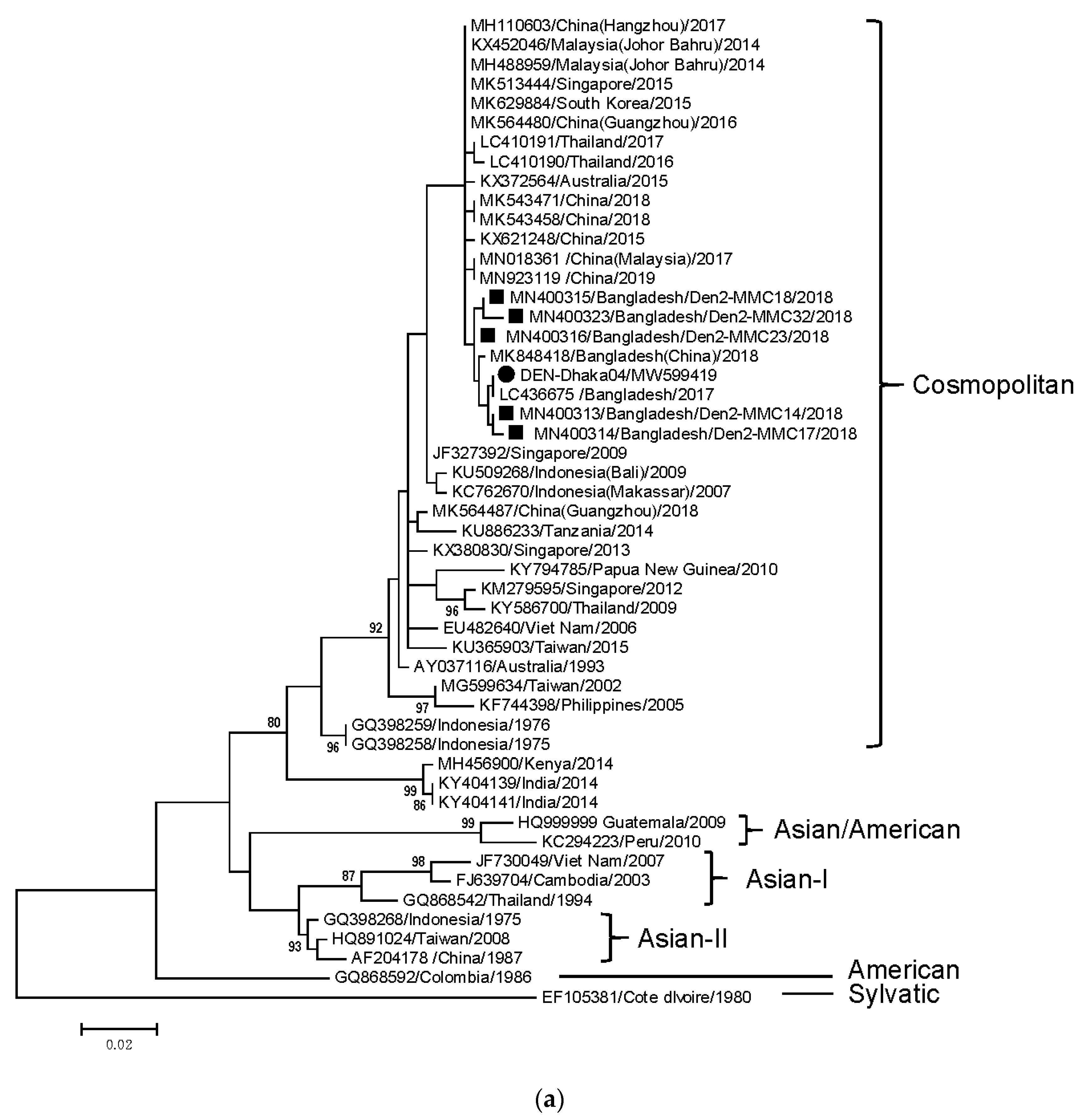
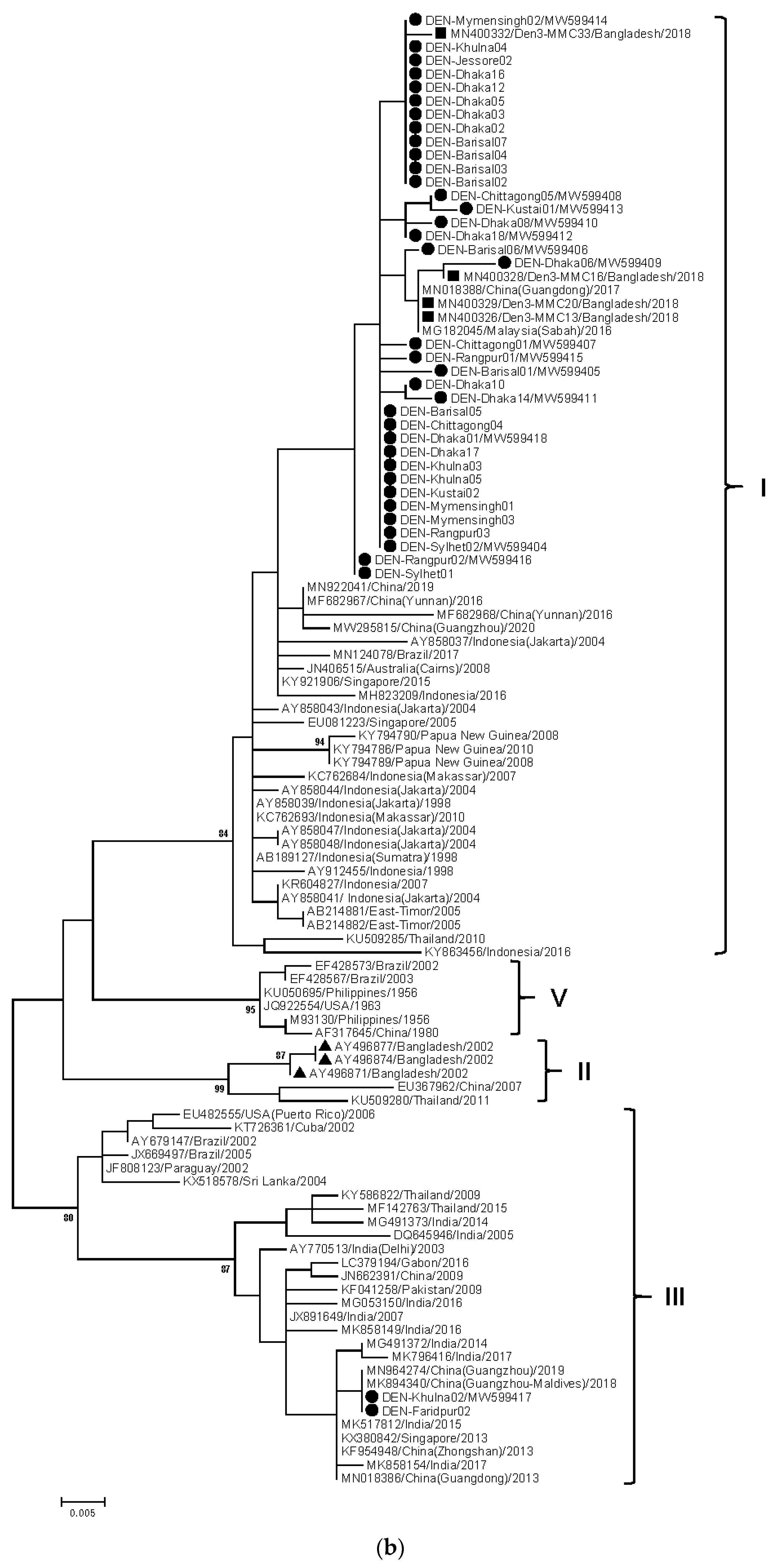
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Tsheten, T.; Gray, D.J.; Clements, A.C.A.; Wangdi, K. Epidemiology and challenges of dengue surveillance in the WHO South-East Asia Region. Trans. R Soc. Trop. Med. Hyg. 2021. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Alvarez, M.; Halstead, S.B. Secondary infection as a risk factor for dengue hemorrhagic fever/dengue shock syndrome: An historical perspective and role of antibody-dependent enhancement of infection. Arch. Virol. 2013, 158, 1445–1459. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Sabeena, S.P.; Varma, M.; Arunkumar, G. Current Understanding of the Pathogenesis of Dengue Virus Infection. Curr. Microbiol. 2021, 78, 17–32. [Google Scholar] [CrossRef]
- Rico-Hesse, R.; Harrison, L.M.; Salas, R.A.; Tovar, D.; Nisalak, A.; Ramos, C.; Boshell, J.; de Mesa, M.T.; Nogueira, R.M.; da Rosa, A.T. Origins of dengue type 2 viruses associated with increased pathogenicity in the Americas. Virology 1997, 230, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Kochel, T.J.; Watts, D.M.; Halstead, S.B.; Hayes, C.G.; Espinoza, A.; Felices, V.; Caceda, R.; Bautista, C.T.; Montoya, Y.; Douglas, S.; et al. Effect of dengue-1 antibodies on American dengue-2 viral infection and dengue haemorrhagic fever. Lancet 2002, 360, 310–312. [Google Scholar] [CrossRef]
- Podder, G.; Breiman, R.F.; Azim, T.; Thu, H.M.; Velathanthiri, N.; Mai, L.Q.; Lowry, K.; Aaskov, J.G. Origin of dengue type 3 viruses associated with the dengue outbreak in Dhaka, Bangladesh, in 2000 and 2001. Am. J. Trop. Med. Hyg. 2006, 74, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Riad, M.H.; Cohnstaedt, L.W.; Scoglio, C.M. Risk Assessment of Dengue Transmission in Bangladesh Using a Spatiotemporal Network Model and Climate Data. Am. J. Trop. Med. Hyg. 2021. [Google Scholar] [CrossRef]
- Muraduzzaman, A.K.M.; Alam, A.N.; Sultana, S.; Siddiqua, M.; Khan, M.H.; Akram, A.; Haque, F.; Flora, M.S.; Shirin, T. Circulating dengue virus serotypes in Bangladesh from 2013 to 2016. Virusdisease 2018, 29, 303–307. [Google Scholar] [CrossRef]
- Ahsan, A.; Haider, N.; Kock, R.; Benfield, C. Possible Drivers of the 2019 Dengue Outbreak in Bangladesh: The Need for a Robust Community-Level Surveillance System. J. Med. Entomol. 2021, 58, 37–39. [Google Scholar]
- Suzuki, K.; Phadungsombat, J.; Nakayama, E.E.; Saito, A.; Egawa, A.; Sato, T.; Rahim, R.; Hasan, A.; Lin, M.Y.; Takasaki, T.; et al. Genotype replacement of dengue virus type 3 and clade replacement of dengue virus type 2 genotype Cosmopolitan in Dhaka, Bangladesh in 2017. Infect. Genet. Evol. 2019, 75, 103977. [Google Scholar] [CrossRef] [PubMed]
- Shirin, T.; Muraduzzaman, A.K.M.; Alam, A.N.; Sultana, S.; Siddiqua, M.; Khan, M.H.; Akram, A.; Sharif, A.R.; Hossain, S.; Flora, M.S. Largest dengue outbreak of the decade with high fatality may be due to reemergence of DEN-3 serotype in Dhaka, Bangladesh, necessitating immediate public health attention. New Microbes New Infect. 2019, 29, 100511. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.U.; Paul, S.K.; Aung, M.S.; Mazid, R.; Alam, M.; Ahmed, S.; Haque, N.; Hossain, M.A.; Paul, S.; Sharmin, R.; et al. Co-circulation of dengue virus type 3-genotype I and type 2-Cosmopolitan genotype in 2018 outbreak in Dhaka, Bangladesh. New Microbes New Infect. 2019, 33, 100629. [Google Scholar] [CrossRef]
- Hsan, K.; Hossain, M.M.; Sarwar, M.S.; Wilder-Smith, A.; Gozal, D. Unprecedented rise in dengue outbreaks in Bangladesh. Lancet Infect. Dis. 2019, 19, 1287. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.S.; Siddiqee, M.H.; Siddiqi, U.R.; Raheem, E.; Akter, R.; Hu, W. Dengue in a crowded megacity: Lessons learnt from 2019 outbreak in Dhaka, Bangladesh. PLoS Negl. Trop. Dis. 2020, 14, e0008349. [Google Scholar] [CrossRef]
- Start Network. Dhaka, Bangladesh: Dengue Outbreak-Anticipatory Briefing Note-27 April 2020. Available online: https://reliefweb.int/report/bangladesh/dhaka-bangladesh-dengue-outbreak-anticipatory-briefing-note-27-april-2020 (accessed on 31 January 2021).
- Ho, T.S.; Wang, S.M.; Lin, Y.S.; Liu, C.C. Clinical and laboratory predictive markers for acute dengue infection. J. Biomed. Sci. 2013, 20, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Rajarethinam, J.; Ang, L.W.; Ong, J.; Ycasas, J.; Hapuarachchi, H.C.; Yap, G.; Chong, C.S.; Lai, Y.L.; Cutter, J.; Ho, D.; et al. Dengue in Singapore from 2004 to 2016: Cyclical Epidemic Patterns Dominated by Serotypes 1 and 2. Am. J. Trop. Med. Hyg. 2018, 99, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Carreño, M.F.; Jiménez-Silva, C.L.; Rey-Caro, L.A.; Conde-Ocazionez, S.A.; Flechas-Alarcón, M.C.; Velandia, S.A.; Ocazionez, R.E. Dengue in Santander State, Colombia: Fluctuations in the prevalence of virus serotypes are linked to dengue incidence and genetic diversity of the circulating viruses. Trop. Med. Int. Health 2019, 24, 1400–1410. [Google Scholar] [CrossRef] [PubMed]
- Patil, J.A.; Alagarasu, K.; Kakade, M.B.; More, A.M.; Gadekar, K.A.; Jadhav, S.M.; Parashar, D.; Shah, P.S. Emergence of dengue virus type 1 and type 3 as dominant serotypes during 2017 in Pune and Nashik regions of Maharashtra, Western India. Infect. Genet. Evol. 2018, 66, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Ahmed, M.U.; Begum, N.; Chowdhury, N.A.; Khan, A.H.; Parquet Mdel, C.; Bipolo, S.; Inoue, S.; Hasebe, F.; Suzuki, Y.; et al. Molecular characterization and clinical evaluation of dengue outbreak in 2002 in Bangladesh. Jpn. J. Infect. Dis. 2006, 59, 85–91. [Google Scholar]
- Waman, V.P.; Kale, M.M.; Kulkarni-Kale, U. Genetic diversity and evolution of dengue virus serotype 3: A comparative genomics study. Infect. Genet. Evol. 2017, 49, 234–240. [Google Scholar] [CrossRef]
- Usme-Ciro, J.A.; Mendez, J.A.; Tenorio, A.; Rey, G.J.; Domingo, C.; Gallego-Gomez, J.C. Simultaneous circulation of genotypes I and III of dengue virus 3 in Colombia. Virol. J. 2008, 5, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suppiah, J.; Ching, S.M.; Amin-Nordin, S.; Mat-Nor, L.A.; Ahmad-Najimudin, N.A.; Low, G.K.; Abdul-Wahid, M.Z.; Thayan, R.; Chee, H.Y. Clinical manifestations of dengue in relation to dengue serotype and genotype in Malaysia: A retrospective observational study. PLoS Negl. Trop. Dis. 2018, 12, e0006817. [Google Scholar] [CrossRef]
- Lee, K.S.; Lai, Y.L.; Lo, S.; Barkham, T.; Aw, P.; Ooi, P.L.; Tai, J.C.; Hibberd, M.; Johansson, P.; Khoo, S.P.; et al. Dengue virus surveillance for early warning, Singapore. Emerg. Infect. Dis. 2010, 16, 847–849. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.A.; Depelsenaire, A.C.; Young, P.R. Clinical and Laboratory Diagnosis of Dengue Virus Infection. J. Infect. Dis. 2017, 215 (Suppl. 2), S89–S95. [Google Scholar] [CrossRef]
- Guo, C.; Zhou, Z.; Wen, Z.; Liu, Y.; Zeng, C.; Xiao, D.; Ou, M.; Han, Y.; Huang, S.; Liu, D.; et al. Global Epidemiology of Dengue Outbreaks in 1990–2015: A Systematic Review and Meta-Analysis. Front. Cell Infect. Microbiol. 2017, 7, 317. [Google Scholar] [CrossRef]
- Soo, K.M.; Khalid, B.; Ching, S.M.; Chee, H.Y. Meta-Analysis of Dengue Severity during Infection by Different Dengue Virus Serotypes in Primary and Secondary Infections. PLoS ONE 2016, 11, e0154760. [Google Scholar] [CrossRef] [Green Version]
- Rafi, A.; Mousumi, A.N.; Ahmed, R.; Chowdhury, R.H.; Wadood, A.; Hossain, G. Dengue epidemic in a non-endemic zone of Bangladesh: Clinical and laboratory profiles of patients. PLoS Negl. Trop. Dis. 2020, 14, e0008567. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Division (District) | No. of NS1-Positive | IgM-Positive (%) | RT-PCR Positive (%) | Serotype (Genotype) * | |
|---|---|---|---|---|---|
| Total | IgM Only/IgM + IgG | ||||
| Barisal | 22 | 16 (72.7) | 4 (18.2)/12 (54.5) | 10 (45.5) | DENV-3 (I) |
| Chittagong | 18 | 14 (77.8) | 7 (38.9)/7 (38.9) | 7 (38.9) | DENV-3 (I) |
| Dhaka | 40 | 27 (67.5) | 7 (17.5)/20 (50) | 18 (45) | DENV-2 (Cosmopolitan), DENV-3 (I) |
| Dhaka (Faridpur) | 11 | 9 (81.8) | 4 (36.4)/5 (45.5) | 2 (18.2) | DENV-3 (III) |
| Khulna | 16 | 9 (56.3) | 5 (31.3)/4 (25) | 6 (37.5) | DENV-3 (I, III) |
| Khulna (Jessore) | 10 | 7 (70) | 4 (40)/3 (30) | 2 (20) | DENV-3 (I) |
| Khulna (Kustia) | 12 | 9 (75) | 4 (33.3)/5 (41.7) | 4 (33.3) | DENV-3 (I) |
| Mymensingh | 10 | 8 (80) | 3 (30)/5 (50) | 3 (30) | DENV-3 (I) |
| Rangpur | 13 | 11 (84.6) | 3 (23.1)/8 (61.5) | 3 (23.1) | DENV-3 (I) |
| Sylhet | 10 | 9 (90) | 6 (60)/3 (30) | 2 (20) | DENV-3 (I) |
| Total | 162 | 119 (73.5) | 47 (29.0)/72 (44.4) | 57 (35.2) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Titir, S.R.; Paul, S.K.; Ahmed, S.; Haque, N.; Nasreen, S.A.; Hossain, K.S.; Ahmad, F.U.; Nila, S.S.; Khanam, J.; Nowsher, N.; et al. Nationwide Distribution of Dengue Virus Type 3 (DENV-3) Genotype I and Emergence of DENV-3 Genotype III during the 2019 Outbreak in Bangladesh. Trop. Med. Infect. Dis. 2021, 6, 58. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed6020058
Titir SR, Paul SK, Ahmed S, Haque N, Nasreen SA, Hossain KS, Ahmad FU, Nila SS, Khanam J, Nowsher N, et al. Nationwide Distribution of Dengue Virus Type 3 (DENV-3) Genotype I and Emergence of DENV-3 Genotype III during the 2019 Outbreak in Bangladesh. Tropical Medicine and Infectious Disease. 2021; 6(2):58. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed6020058
Chicago/Turabian StyleTitir, Snigdha Rahman, Shyamal Kumar Paul, Salma Ahmed, Nazia Haque, Syeda Anjuman Nasreen, Khondoker Shoaib Hossain, Fahim Uddin Ahmad, Sultana Shabnam Nila, Jobyda Khanam, Neaz Nowsher, and et al. 2021. "Nationwide Distribution of Dengue Virus Type 3 (DENV-3) Genotype I and Emergence of DENV-3 Genotype III during the 2019 Outbreak in Bangladesh" Tropical Medicine and Infectious Disease 6, no. 2: 58. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed6020058