1. Introduction
Staphylococcal food poisoning is one of the most common food-borne diseases worldwide that results from the ingestion of pre-formed enterotoxins produced by
Staphylococcus aureus [
1]. The European Food Safety Authority (EFSA) reported in 2015 a total of 434 food-borne outbreaks caused by staphylococcal toxins [
2]. In the United States, 352 outbreak-associated illnesses and 27 hospitalizations were caused by
S. aureus toxins in 2016 [
3]. The pathogenicity of this bacterium is due to a combination of toxin-mediated virulence, invasiveness and antibiotic resistance [
4]. Indeed, methicillin-resistant
S. aureus (MRSA) is one of the leading causes of hospital-acquired infections.
It is also remarkable the increase in infections caused by multidrug-resistant
S. aureus and community-associated methicillin-resistant
S. aureus (CA-MRSA) as it results in a diminished effectiveness of the antibiotic treatment [
5]. For instance, many methicillin-resistant strains of
S. aureus (MRSA) show a decreased susceptibility to glycopeptides such as vancomycin [
6,
7,
8]. An interesting survey addressed by the European Centre for Disease Prevention and Control (ECDC) in the period 2011–2012 showed a 41% of invasive
S. aureus isolates carrying methicillin resistance [
9].
Since multidrug resistance is rapidly evolving in several species including
S. aureus [
10], there is a clear need for novel approaches to circumvent this problem. Bacteriophages have been proposed as a suitable antimicrobial alternative to the use of antibiotics, as they are natural enemies of bacteria [
11,
12]. Currently, there is a renaissance of phage therapy studies in western countries, and the results from clinical trials in animals are confirming the efficacy of phages as therapeutics. Despite the regulatory hurdles, several biotechnological companies are assaying bacteriophages in human clinical trials to promote their future commercialization (reviewed by [
13]). Bacteriophages are also suitable to control pathogenic bacteria along the food chain [
14]. Several studies have demonstrated their effectiveness in controlling bacterial pathogens in agro-food industry, such as
Salmonella,
S. aureus,
Campylobacter jejuni,
Escherichia coli O157: H7, and
Listeria monocytogenes [
15]. Usefulness of bacteriophages in food safety includes their application as disinfectants to remove bacterial biofilms from industrial surfaces [
16,
17,
18] and the development of tools to improve pathogen detection [
19]. Since the approval of phage-based products to use in the food industry sector by the US FDA in 2006, some new bacteriophage products are commercially available. These products consist of a mixture of one or more bacteriophages infecting
L. monocytogenes (PhageGuard Listex, Micreos Food Safety B.V., Wageningen, The Netherlands; ListShieldTM, Intralytix Inc., Baltimore, MD, USA),
E. coli O157: H7 (EcoShieldTM, Intralytix Inc., Baltimore, MD, USA) and
Salmonella (SalmoFreshTM, Intralytix Inc., Baltimore, MD, USA; PhageGuard S, Micreos Food Safety B.V., Wageningen, The Netherlands).
We had previously isolated and characterized the
S. aureus infecting phage vB_SauM-phiIPLA-RODI (in short, phiIPLA-RODI) that belongs to the
Myoviridae family and exhibits a wide host range [
16]. In challenge assays against
S. aureus cultures, the phage reduced the viable counts by 5 log units in 8 h. Moreover, exposure of biofilms to this phage also reduced adhered bacteria by 2 log units [
16]. Therefore, the lytic ability of phiIPLA-RODI against planktonic and sessile cells supports its potential as an antimicrobial to both remove staphylococcal biofilms and to treat
S. aureus infections.
The use of phages as antimicrobials in the clinic and the food industry requires large-scale and reproducible production of phage cultivation. Temperature, media composition, bacteria and bacteriophage concentration are factors that can affect phage production [
20].
In this regard, the aim of the present work was to optimize the propagation of phage phiIPLA-RODI by using a food-grade species instead of the original pathogenic host in a laboratory scale applying the response surface methodology (RSM). This methodology is a collection of mathematical and statistical techniques that allow the analysis of the relationship between a set of controllable experimental factors and the observed results of the variable of interest (response) to optimize the response [
21,
22]. Unlike the present work, the scarce studies on the production of bacteriophages that have already applied RSM have used the original bacterial host and other factors that affect phage production [
23].
We used a food-grade strain of the species Staphylococcus xylosus as an alternative host to reduce the risk of growing massive quantities of a bacterial pathogen. The effects of different levels of the expected influential variables such as temperature, agitation, initial bacterial and bacteriophage concentration were investigated by using RSM. The equation at the region of maximum phage yield was validated, and a definitive model incorporating the validation data was deduced. Based on this model, and using the desirability approach, the optimal operating conditions were established.
2. Materials and Methods
2.1. Bacterial Strains, Bacteriophage and Media
Staphylococcal strains were isolated in Baird-Parker (BP) agar supplemented with egg yolk tellurite emulsion (Scharlau, Barcelona, Spain) and routinely grown in Tryptic Soy Broth (TSB, Scharlau, Barcelona, Spain) with shaking (Excella E24 Incubator Shaker, New Brunswich Scientific, Edison, NJ, USA) or Tryptic Soy Agar (TSA), at 37 °C.
The food-grade strain Staphylococcus xylosus CTC1642, isolated from a fermented meat product (IRTA, Monells, Girona, Spain) and the strain S. aureus IPLA1, isolated from a dairy product (IPLA-CSIC, Villaviciosa, Asturias, Spain), were used to propagate the S. aureus-infecting bacteriophage phiIPLA-RODI.
Phage titer was assessed by plaque assay. One-hundred μL of an overnight culture (about 10
9 colony-forming unit (CFU)/mL) of the host strains were mixed with 100 μL of serial phage dilutions. These mixtures were added to 5 mL of molten TSA overlay (0.7% agar), poured onto TSA plates, incubated at 37 °C for 18–24 h, and the lysis plaques counted [
16].
For selecting the food-grade host for phage propagation purposes, preliminary plaque assays with phage phiIPLA-RODI were performed on several food-grade strains, kindly provided by Dr Margarita Garriga (IRTA, Monells, Girona, Spain): S. xylosus strains (CTC1638, CTC1642 and CTC1644) and Staphylococcus carnosus strains (CTC6064, CTC6071 and CTC6072). The strain S. carnosus TM300, kindly provided by Dr Gabi Bierbaum (University of Bonn, Germany) was also tested. The efficiency of plating (EOP) was calculated by dividing the phage titer obtained on each of the tested strains by the phage titer on the reference strain S. aureus IPLA1.
The host range of phiIPLA-RODI lysates obtained by propagation on CTC1642 and IPLA1 was determined by plaque assay on some other staphylococcal species previously tested [
16]. Some of them such as
S. lugdunensis ZL5-11,
S. pasteuri ZL16-6,
S. arlettae ZL114-5,
S. xylosus ZL61-2,
S. gallinarum ZL90-5 and
S. kloosii ZL74-2, were isolated from women’s breast milk [
16], while others (
S. aureus IPLA15 and IPLA16) were isolated from meat industry surfaces [
24]. The efficiency of plating (EOP) of both phage lysates on each strain was calculated as indicated above.
2.2. Biofilm Removal by Phage phiIPLA-RODI Propagated on S. xylosus CTC1642 and S. aureus IPLA1
Overnight cultures of
S. aureus IPLA16 were diluted to 10
6 CFU/mL into fresh TSB supplemented with 0.25% glucose. Aliquots of 200 μL of each culture were poured into the wells of a polystyrene microtiter plate (TC Microwell 96U w/lid nunclon DSI plates, Thermo Scientific, Madrid, Spain). Biofilms were grown for 24 h at 37 °C. Wells were then washed twice with PBS buffer (137 mM NaCl, 2.7 mM KCl, 10 mM 138 Na
2HPO
4 and 2 mM KH
2PO
4; pH 7.4). To compare the biofilm degradation ability of each phage lysate, 200 μL of phiIPLA-RODI propagated on
S. aureus IPLA1 or
S. xylosus CTC1642 were added to each well (10
8 plaque-forming unit (PFU)/well). SM buffer (20 mM Tris HCl, 10 mM of MgSO
4, 10 mM of Ca(NO
3)
2 and 0.1 M of NaCl, pH 7.5) was added for control purposes. The microwell plates were incubated for 4 h at 37 °C. The supernatants were removed, and wells washed once with SM buffer (20 mM Tris HCl, 10 mM MgSO
4, 10 mM Ca(NO
3)
2 and 0.1 M NaCl, pH 7.5) and air-dried for 15 min at room temperature. The biomass adhered to the wells was determined by crystal violet (0.1%,
w/
v) staining as described previously [
25]. All the assays were performed using three biological replicates.
2.3. One-Step Growth Curve
One-step growth curve assays were carried out with phage phiIPLA-RODI, using the sensitive strains
S. aureus IPLA1 and
S. xylosus CTC1642. A standardised protocol, previously described [
26], was adapted for this study.
Curves were performed in TSB supplemented with Ca(NO3)2 (10 mmol L−1) and MgSO4 (10 mmol L−1) using a multiplicity of infection (MOI) of 0.01. Mid-exponential-phase cultures (10 mL) of S. aureus IPLA1 and S. xylosus CTC1642 (OD600 = 0.1) were collected by centrifugation and suspended into 1 mL of fresh TSB. The phage was added and allowed to adsorb for 5 min at 37 °C with shaking. The mixture was then centrifuged, pelleted cells resuspended in 10 mL of TSB, and incubation continued at 37 °C. Samples were first taken at 5 min intervals for 30 min, and subsequently at 10 min intervals. Each sample was immediately diluted and plated for phage titration.
2.4. Bacteriophage Amplification: Conventional Phage Propagation
Bacteriophage phiIPLA-RODI was routinely propagated on S. aureus IPLA1 and S. xylosus CTC1642, according to the following procedure: TSB broth was inoculated with 1% (v/v) inoculum of an overnight culture of the strains indicated above, and incubated at 37 °C with shaking until an OD600 = 0.1 (107 CFU/mL) was reached. Phage was added to the bacterial culture at MOI of 1.0 and incubation proceeded for a further 3.5 h at 37 °C with shaking. Phage preparations were obtained by centrifugation and further filtration to remove bacterial cells and debris.
The phage titer was determined by the plaque assay using 100 µL of an
S. aureus IPLA1 overnight culture as a host, and 100 µL of the phage dilution. This mixture was added to 5 mL of molten TSA overlay (0.7% agar) and poured onto TSA plates and incubated at 37 °C for 18–24 h [
16].
2.5. Bacteriophage Amplification: Phage Propagation for Optimization Purposes
Frozen stocks (−80 °C) of the strain S. xylosus CTC1642 (about 108 CFU/mL) were quickly thawed and used to inoculate at different concentrations (CFU/mL) in 50 mL Falcon tubes filled with 10 mL of TSB broth. The actual viable cell counts were determined immediately after inoculation by plating decimal dilutions of samples onto TSA.
For phage propagation, cultures were infected with different concentrations (PFU/mL) of phage phiIPLA-RODI. The combined effect of initial phage titer, initial host concentration, temperature, and agitation on phage yield (final phage titer) and phage amplification ratio (estimated as the difference between final phage titer and the initial phage titer, expressed in log10 values) was evaluated after 3.5 h of incubation. The phage titer was determined as described in the previous section.
2.6. Experimental Design
The assays consisted of three successive designs that allowed estimating the effects of four independent continuous variables (initial phage titer, initial bacterial concentration, temperature, and agitation) on the phage yield and optimizing the conditions that could eventually result in the highest phage yield. The first design was used for an initial exploration of the experimental region in which phage production was expected to occur. It consisted of a quadratic Central Composite Design with the following ranges of the variables: initial phage titer (5.00 to 8.00 log
10 PFU/mL); initial bacterial concentration (6.0 to 8.0 log
10 CFU/mL); temperature (21 to 37 °C) and agitation (20 to 250 rpm). The objective of the second design was focused on the effect of temperature, using a D-optimal design, at fixed agitation (135 rpm). It had the following ranges for the variables: initial phage titer (6.0 to 8.0 log
10 PFU/mL); initial bacterial concentration (5.0 to 7.0 log
10 CFU/mL) and temperature (34 to 40 °C). Finally, the third design aimed at developing a Response Surface equation, using Central Composite Design, to predict phage production in the region of highest yield. It included only the initial phage titer (5.79 to 7.21 log
10 PFU/mL) and initial bacterial concentration (5.59 to 8.41 log
10 CFU/mL), while temperature and agitation were fixed at 38 °C and 135 rpm, respectively. The characteristics of these designs (and their respective yields) are summarized in
Table 1. The levels of variables for all the designs were given by the program Design-Expert software version 7.0 (State-Ease, Inc., Minneapolis, MN, USA), provided their ranges and type of design. The order of the run performance was always randomly chosen. However, the values for initial bacterial populations, although intended to be those proposed by the designs, were difficult to fix accurately. Therefore, the actual bacterial concentrations reached just after inoculation, as determined by viable cell counts, were used for the statistical analysis.
2.7. Analysis of Results, Model Validation, and Final Response Surface (RS) Equation
The effects of the above-indicated variables (factors) on phage propagation (initial phage titer (A), initial bacterial concentration (B), temperature (C) and agitation (D)) were analyzed by the response surface methodology (RSM), using the following general quadratic model:
where
k was 4, 3 and 2 for first, second, and third design, respectively [
27]. The significant influential variables were those suggested by the sequential sum of squares (Type I) and supported by the corresponding ANOVA (partial sum of squares type III). The final models were obtained by a stepwise process, using
p ≤ 0.05 and
p ≥ 0.10 as criteria for entering and removal of variables, respectively. The effects of the variables and model fits were also checked graphically. For the validation of the third model, nine additional assays were performed with the initial phage titer fixed at 6.5 log
10 PFU/mL and the initial bacterial populations around the levels of maximal phage yield. A final equation for the model was developed by enlarging the data from the third design with the validation results. This last model was used for obtaining the conditions which maximize the phage yield and amplification.
2.8. Statistical Analysis
Statistical analyses for phage propagation were performed using the statistical package IBM SPSS Statistics for Windows Version 23 (IBM Corp. Armonk, NY, USA). Data related to phage propagation carried out with the control host (S. aureus IPLA1), and test host (S. xylosus CTC1642) were subjected to one-way ANOVA, and the Student-Newman-Keuls (SNK) test was used for comparison of means at a level of significance p < 0.05. Three biological replicates were used in all the assays.
For optimizing the phage propagation on S. xylosus CTC1642, the experiments were always designed and analyzed using Design-Expert software version 7.0 (State-Ease, Inc., Minneapolis, MN, USA). Final optimization was achieved using the desirability approach, which finds operating conditions that provide the “most desirable” response values. The desirability function di (Yi) assigns numbers between 0 (undesirable value) and 1 (ideal response) to the possible values of Yi (phage yield or phage amplification ratio). Usually, the individual desirability values are combined using the geometric mean, which gives the overall desirability (D) which is maximized with respect to the controlled variables.
4. Discussion
To exploit the advantages of bacteriophages as antimicrobials, it is essential to adapt their production to meet quality and safety requirements for sustainable phage therapy products [
28]. In this context, the use of a surrogate host such as a food-grade bacterium minimizes the risk of toxin or host contamination on phage preparations. The ability of phage phiIPLA-RODI to infect a wide spectrum of staphylococcal strains [
16] allowed selecting a
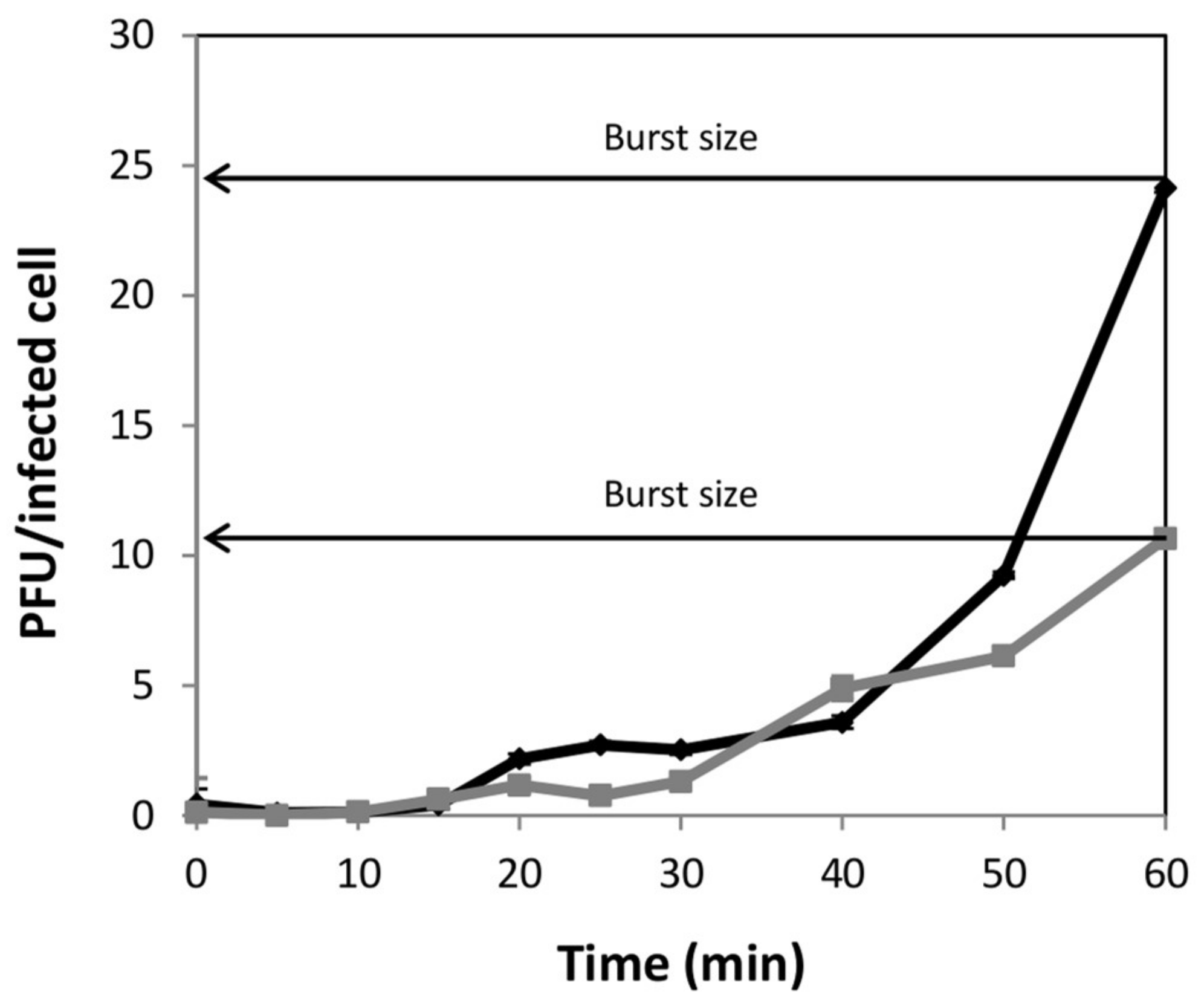
S. xylosus strain, from meat origin, to propagate the phage. A reduction in the phage yield (final phage titer) could be expected due to the lower burst size calculated in
S. xylosus compared with
S. aureus (10 versus 25 phage particles per infected cell). In this regard, a reduction in the burst size was also observed when the
Salmonella phage phi PVP-SE3 was propagated in the non-pathogenic strain
E. coli BL21 [
29], while a similar number of phage particles per infected cell were obtained after infection of
S. xylosus by the
S. aureus infecting phage Team1 [
30]. As previously observed in other phages [
30], the host range and the EOP value shown by phiIPLA-RODI when propagated on both
S. xylosus CTC1642 and
S. aureus IPLA1 were fairly similar. Besides, the ability of phiIPLA-RODI to remove biofilms was similar regardless of the bacterial host used for phage propagation. Overall, the lack of pathogenicity of the host strain
S. xylosus CTC1642 and consequently, the safety of the final phage product justify, in our opinion, the use of the food-grade strain even though a lower phage yield is obtained. In an attempt to counteract this disadvantage, we optimized the phiIPLA-RODI propagation process by using RSM since it is handy for studying processes in which the response is influenced by several variables [
22]. In fact, this method was previously used to optimize the production of an
E. coli infecting filamentous phage [
23]. It should be noticed, however, that filamentous phages do not reproduce by lysing bacteria, instead, they are secreted into the environment without killing the host, while the life cycle of myovirus phages ends with the lysis of the host. Likewise, Grieco et al. [
23] have used temperature, dissolved oxygen and pH as independent variables while, in our study, phage and bacterial concentration, temperature and agitation were tentatively considered initial influential on the phage yield when
S. xylosus is used as propagation host. Remarkably, the initial bacterial concentration was particularly significant for phiIPLA-RODI propagation as it had a quadratic effect on the phage yield.
The propagation process is the result of a succession of several infection cycles, and each of them consists of an adsorption period, nucleic acid uptake, latent period and progeny release. The duration of these steps and the number of phage particles produced per infected cell will determine the phage yield [
31,
32]. It is well known that phage-host interactions are affected by environmental conditions that alter the physiological state of bacteria, through changing their susceptibility to phage infection and the phage productivity [
33]. Moreover, optimal bacteriophage infection traditionally requires bacterial host growing exponentially, although some bacteriophages like T4 can efficiently infect
E. coli in stationary phase and kill the host after a hibernation period [
34].
In this context, a first design was used as initial guidance for approaching the production of phages in an experimental region (space limited by the range of variables) in which, according to previous non-systematic assays, high phage yield (response) were expected. The results showed that agitation within the range assayed did not affect phage yield, however, agitation might favor the encounter between bacteria and phage needed for the phage infection to take place and prevent the bacterial cell from sedimentation. Therefore, the use of an intermediate level (135 rpm) was fixed for further assays. For the variable temperature, a wide range of 21–37 °C was initially selected to include values even lower than the optimal for
Staphylococcus growth because other authors have previously observed improved phage production at temperatures below the optimum for bacterial host growth [
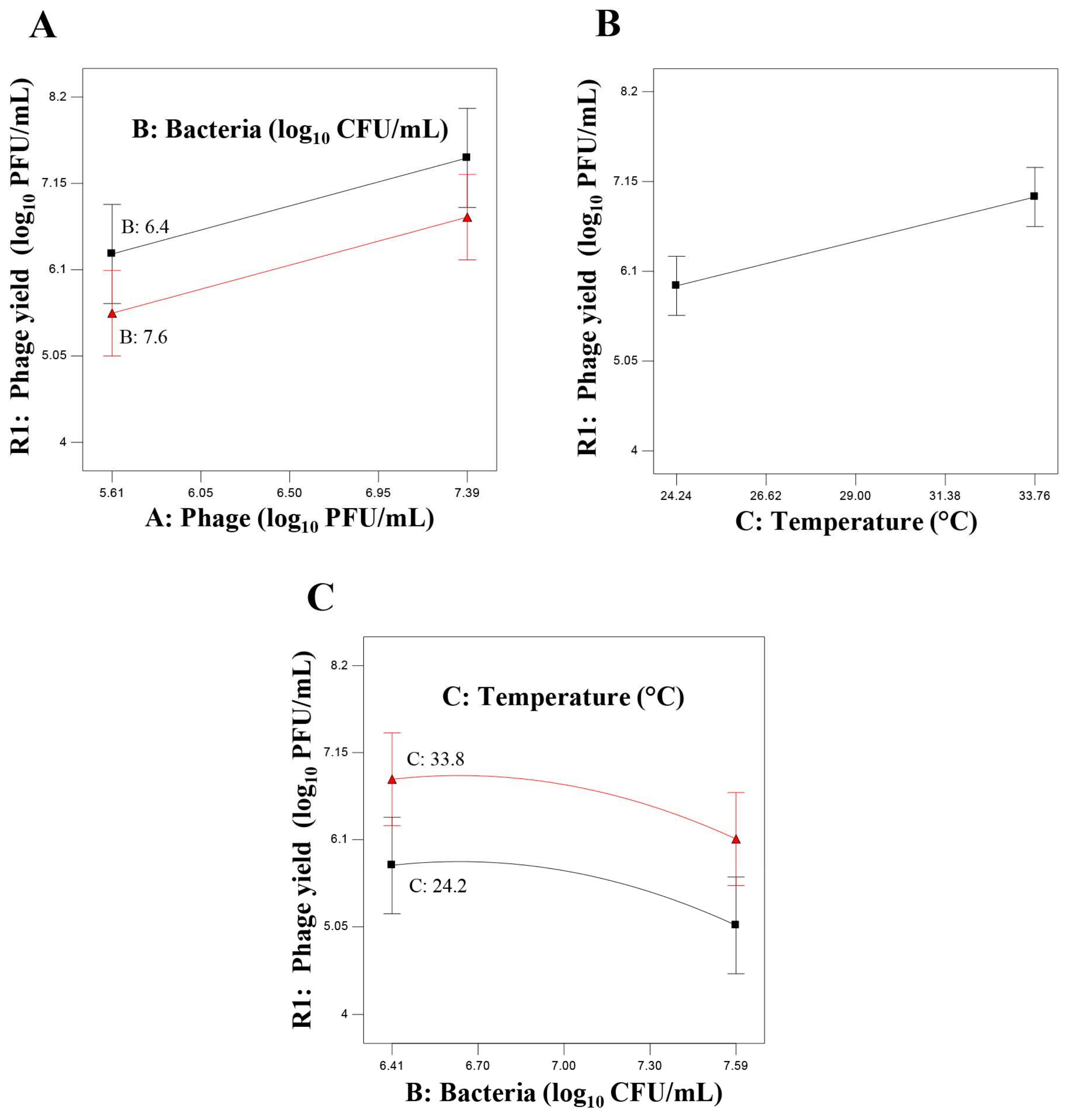
23]. The data from the assays proposed by the first design pointed out that the range of temperature was not appropriate since the effect on the phage yield was linear and increased progressively as the temperature rose. Therefore, these results supported the notion of using a higher temperature around those that favor host metabolism for additional designs. The linear effect of initial phage titer and the quadratic effect of the initial bacterial concentration on the phage yield also suggested to increase the initial phage concentration and to decrease the initial bacteria concentration. Hence, the analysis of the data from the experiments proposed by the first design reduced the influential variables to three: initial phage titer, initial bacterial concentration and temperature.
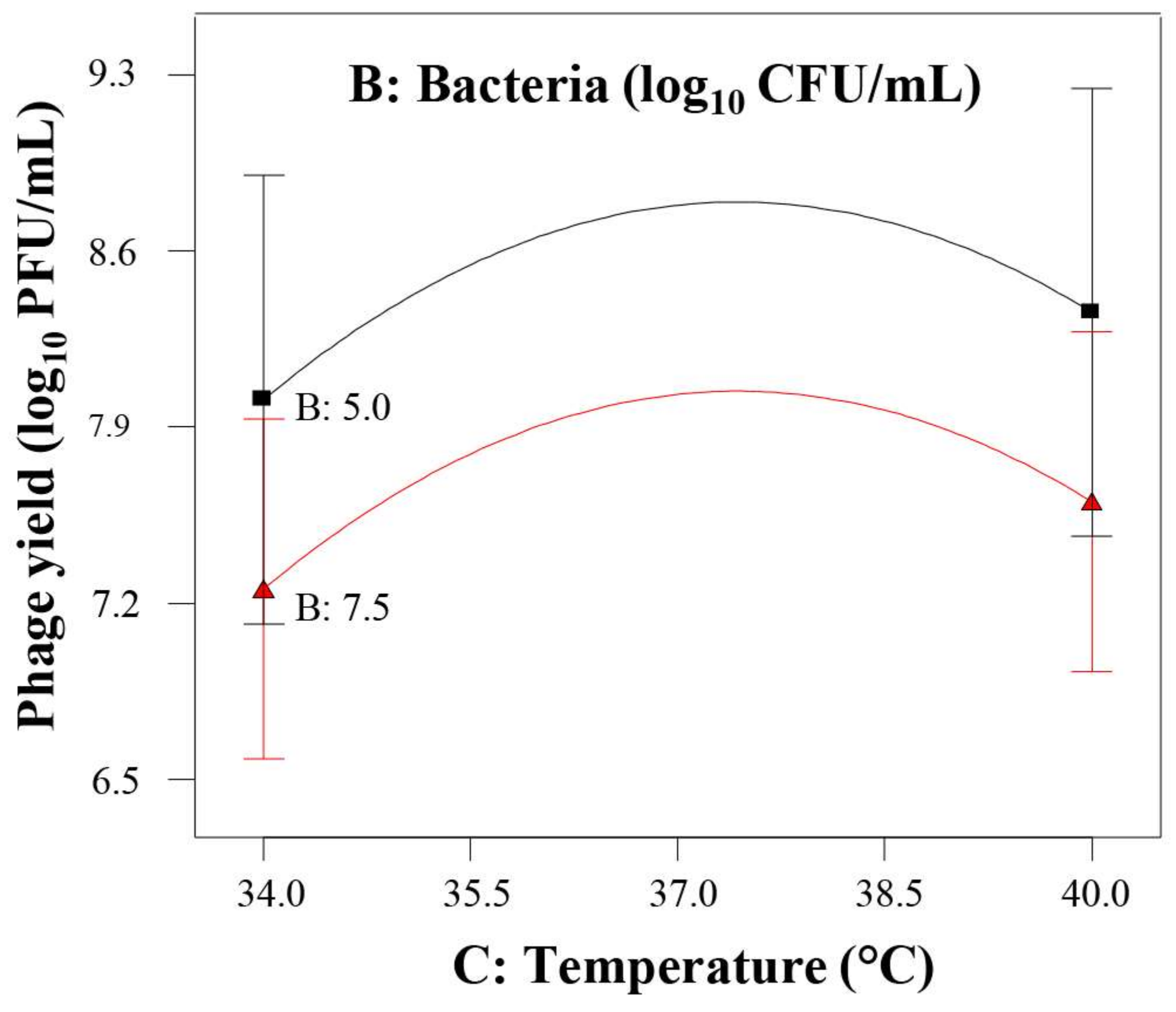
Subsequently, the main goal of the second design was exploring the effect of temperature in detail, covering the region of optimal host growth and, eventually, the optimum level of phage yield. Its results confirmed the pertinence of increasing temperature and showed a quadratic effect defining the optimum temperature at 38 °C that was adopted for further assays. The range of the initial bacterial population was still appropriate, and the ANOVA led to the identification of a quadratic effect (p ≤ 0.10) for this variable, in agreement with the results of the first design. The effect of initial phage concentration was not statistically significant under the experimental conditions proposed by the second design. The reason for this observation could be the different ranges of temperature tested. Indeed, it is well known that the lytic phage infection process is very much dependent on the bacterial host metabolic machinery. Therefore, a temperature that improves bacterial metabolic activity will result in a better phage yield. In this regard, it should be noted that the range used in the second design (34–40 °C) is more appropriate for the bacterial host growth than that used in the first design (21–37 °C), resulting in an enhanced phage yield and consequently, the initial phage concentration has not longer statistically significant effect on the response.
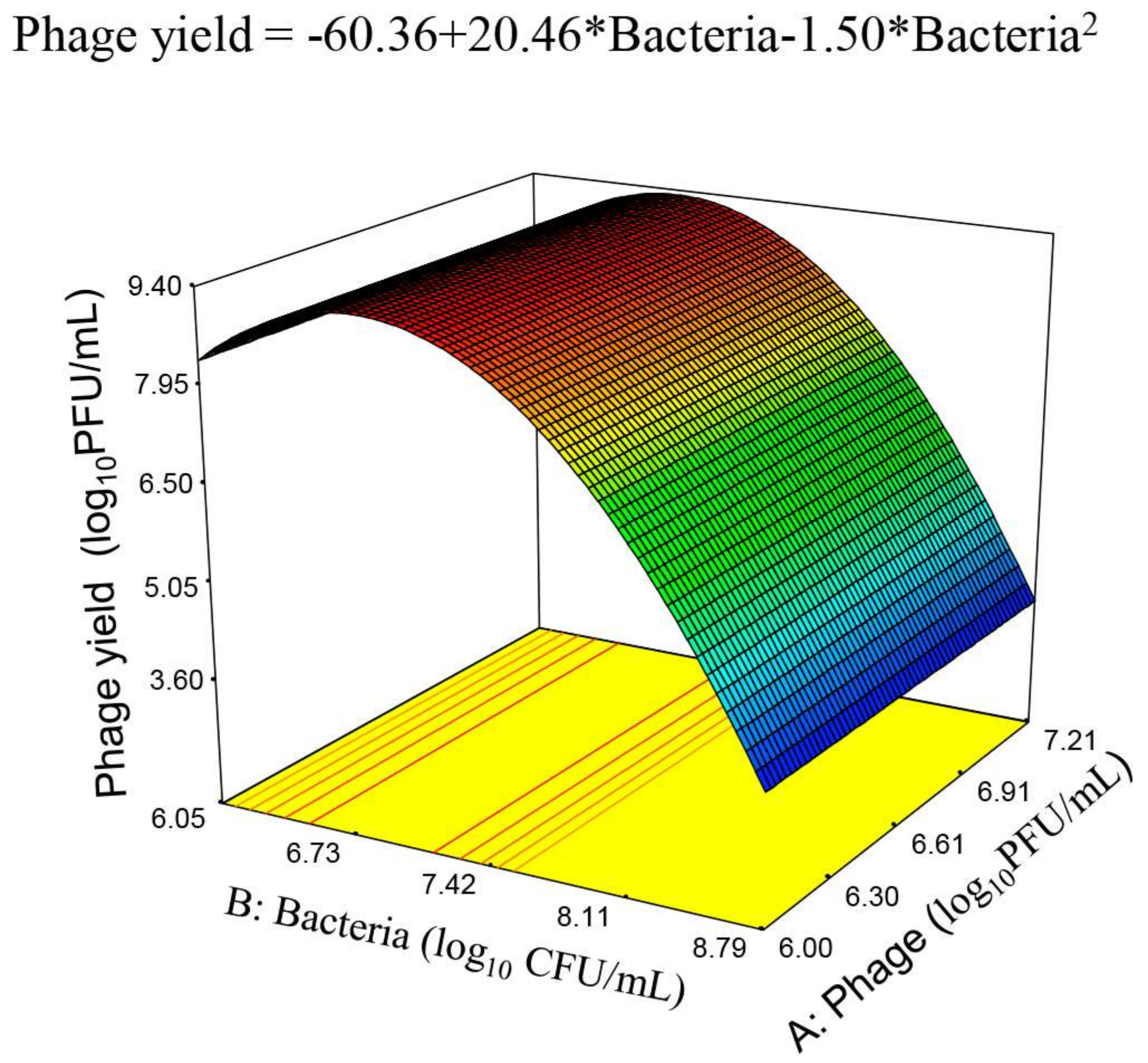
Once the optimum temperature was defined, the influential variables were reduced to only two: initial phage titer and bacterial concentration. Further analysis of the relationship between these variables was approached by the third design in which the interest was focused on finding the equation of the RS defining the region of maximum phage production, already outlined from the results of the previous design. The ranges of both variables were established taking into account that, for an efficient infection process, phages should be able to encounter the bacterial cell host. For this purpose, the bacterial population must exceed its proliferation threshold [
35,
36], to facilitate the initial phage titer to increase. It should be noticed that MOI (as phage/bacteria ratio) has not been used as independent variable in the current study because similar MOI values are obtained from different concentrations of initial phage and bacteria. In fact, the particular phage and bacteria concentrations used for phage propagation did really result in different values of the final phage titer. Having fixed agitation and temperature, the data from the experimental assays of the third design confirmed the quadratic effect of bacteria regardless of the initial phage titer, which was not significant within the studied range. All the parameters related to the model fit were appropriate. In addition, several independent validation experiments showed also good agreement with its predictions and, remarkably, the incorporation of validation data into those from the third model allowed deducing a final RS model with a higher degree of freedom for the medium standard error (MSE) and narrower confidence limits for the estimations. Also, from the third model, enlarged with the validation data, a final model could be deduced which led not only to the conditions of optimal phage production but also revealed a sharp decrease in the phage yield at bacterial concentrations above 7.5 log
10 CFU/mL. This behavior could be due to the adsorption of a proportion of the new viral progeny to the host cells that were not initially infected, which could partially hamper its detection by phage titration.
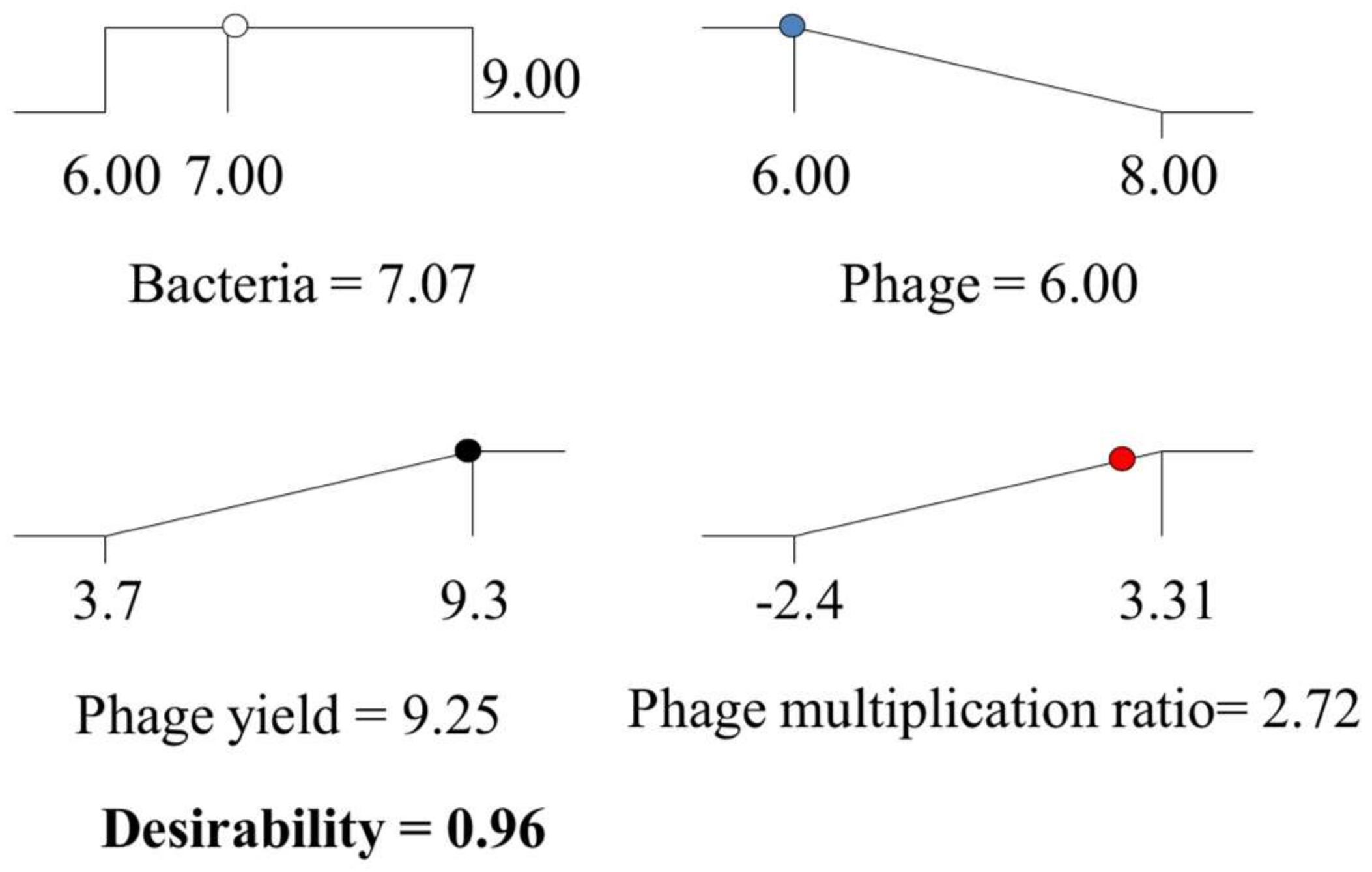
The optimization process allowed therefore maximizing the phage yield and the phage amplification ratio, while minimizing the initial phage concentration and using an initial bacterial population between the ranges studied in the third design. The optimization process deduced several possible combinations. Among them, it was chosen that providing the highest desirability which consisted of the initial bacterial population, 7.07 log
10 CFU/mL; and initial phage titer, 6.00 log
10 PFU/mL (
Figure 5), while maintaining agitation (135 rpm) and temperature (38 °C) at their fixed levels. Using these conditions, it is expected a phage yield of up to 9.25 ± 0.35 log
10 CFU/mL along with a phage amplification ratio of 2.72 log units over the initial phage titer. The global desirability, defined as the geometric mean of desirability values of the phage yield and phage amplification ratio, would be in this case high (0.96) and fairly close to the most “desirable” response 1.00. These results support the use of the food-grade strain
S. xylosus CTC1642 as an appropriate alternative of the pathogenic strain
S. aureus IPLA1 for phiIPLA-RODI propagation at the setting conditions established by the final RS model. The setting parameters could be the starting point for performing the upscaled production of the phage that would be required for its potential use in clinical [
28] and food safety [
15] applications.

 ,
, 




 ) and phage multiplication ratio (
) and phage multiplication ratio (  ), as a function of initial bacterial populations (
), as a function of initial bacterial populations (  ) and phage titers (
) and phage titers (  ), using the desirability approach as implemented in Design Expert.
), using the desirability approach as implemented in Design Expert.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}